Reproductive Soldier Development Is Controlled by Direct Physical Interactions with Reproductive and Soldier Termites


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Termites
2.2. Observation of Soldier and Reproductive Soldier Differentiation
2.3. Observation of Mating Behavior
2.4. Morphometry of Emerged Individuals
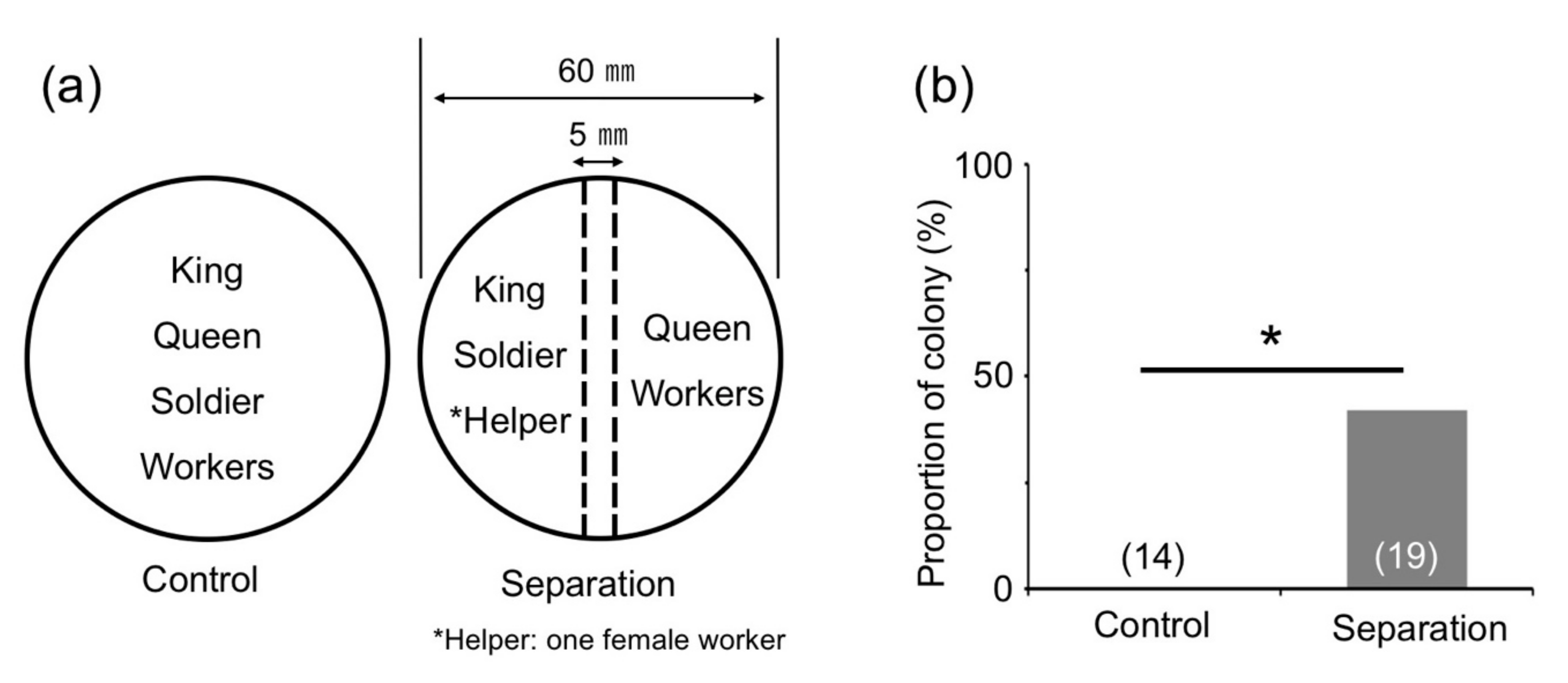
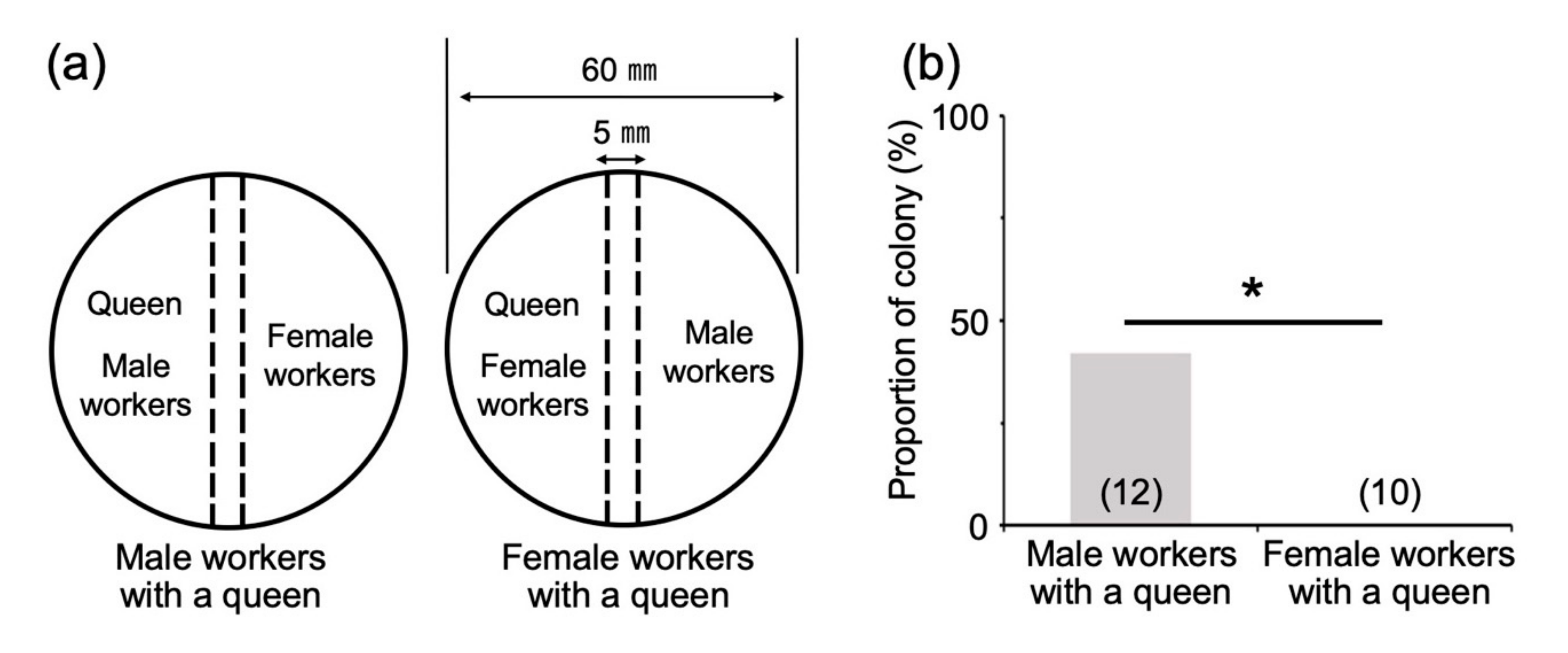
2.5. Effects of Direct or Indirect Contact with Reproductives and Soldiers
3. Results
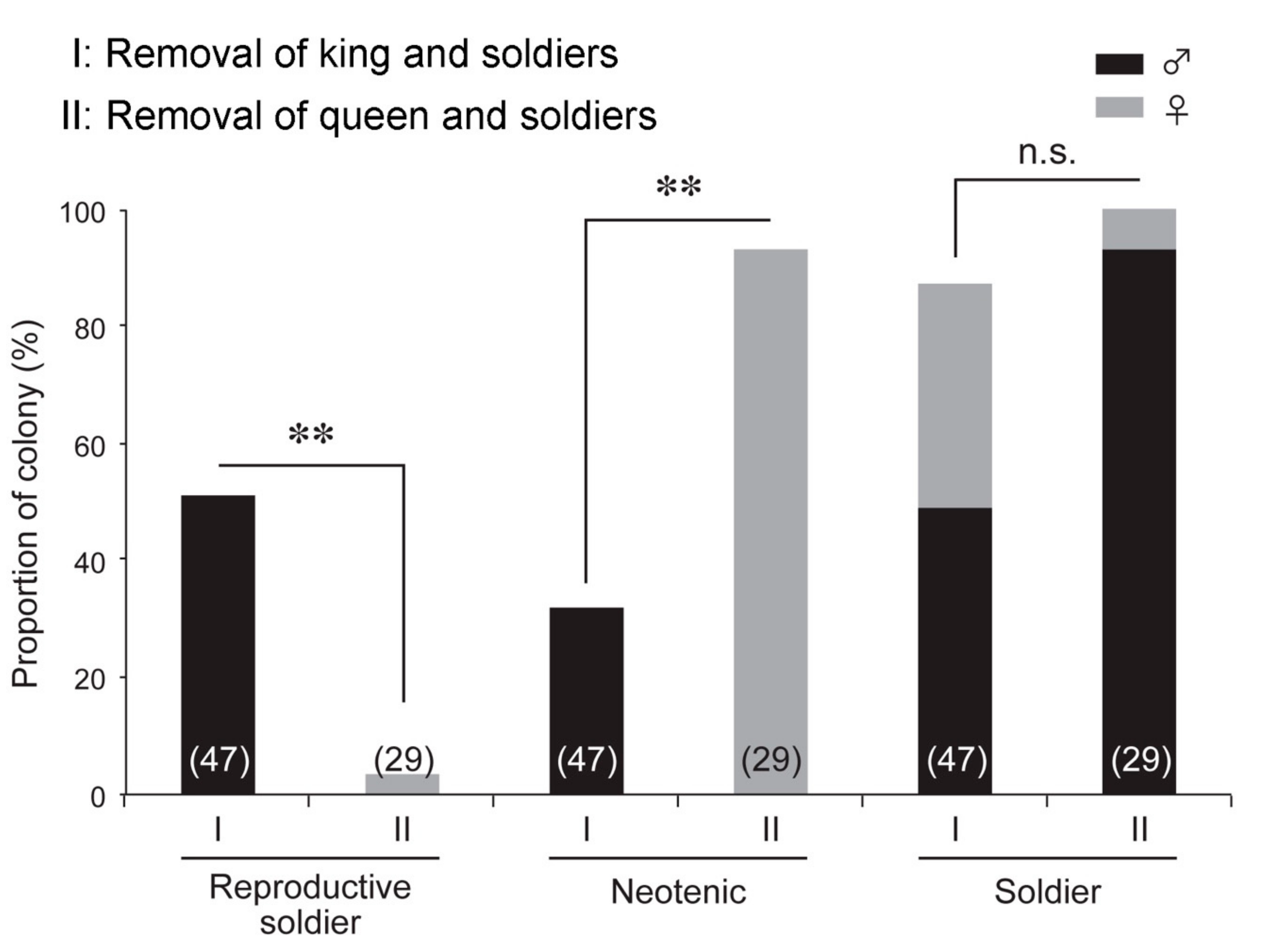
3.1. Effects of King and Soldier Removal Treatment on Reproductive Soldier Development
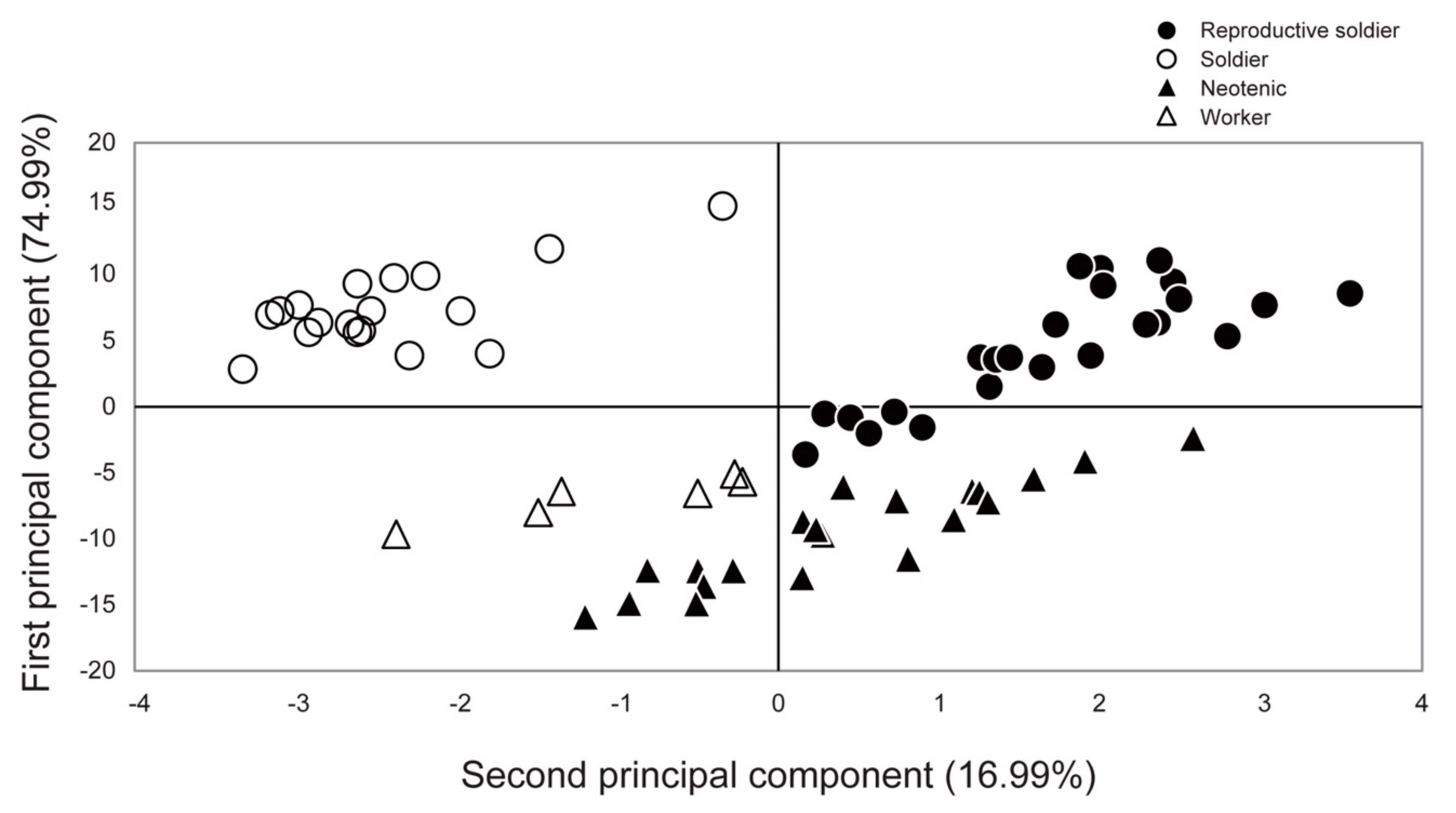
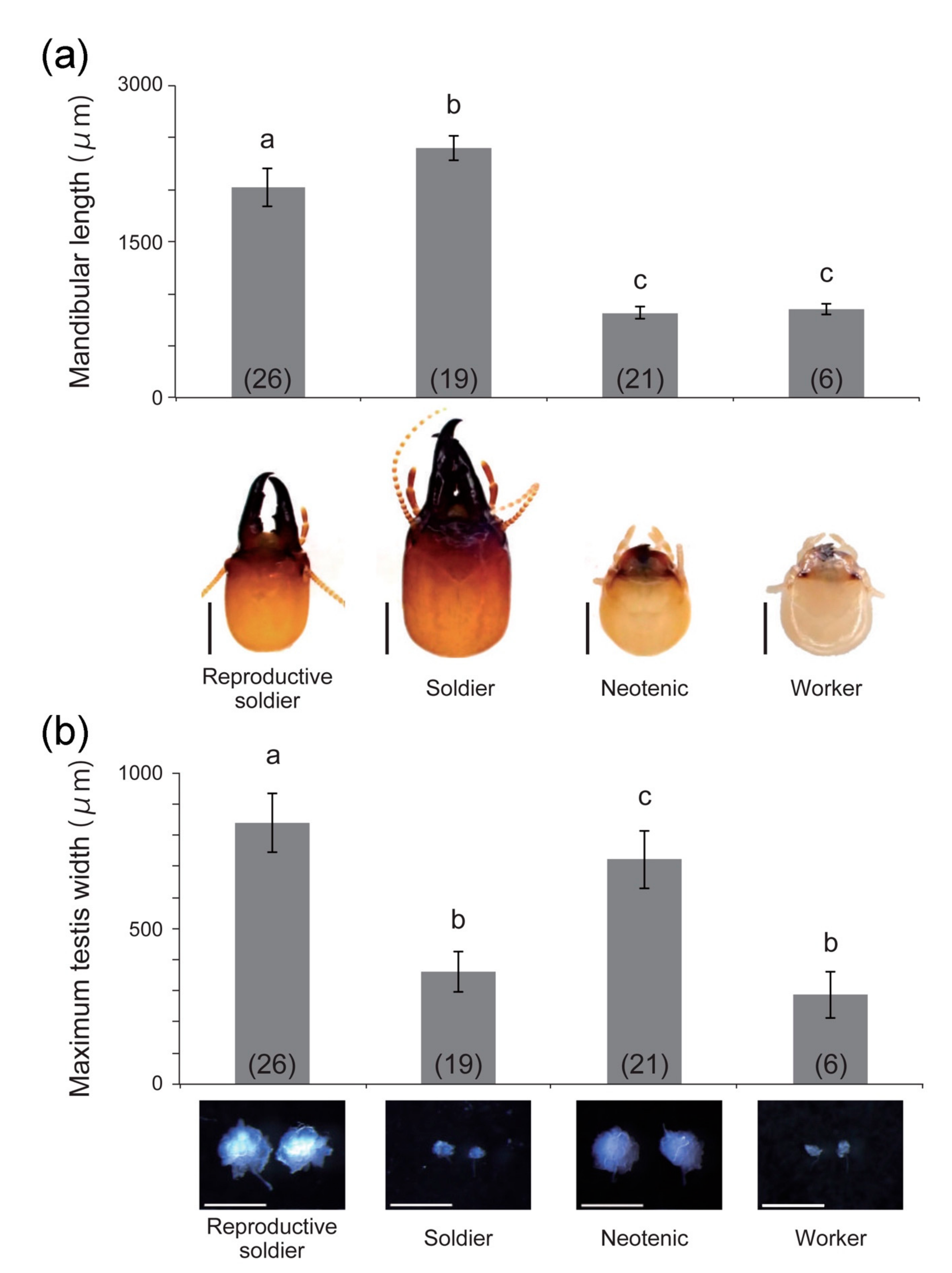
3.2. Behavior and Morphology of Reproductive Soldiers
3.3. Effects of Queen and Soldier Removal Treatment on Reproductive Soldier Development
3.4. Effects of Direct Contact with Reproductives and Soldiers
4. Discussion
4.1. Morphological Changes during Reproductive Soldier Differentiation
4.2. Sexual Differences in Reproductive Soldier Differentiation
4.3. Environmental Cues for Male Reproductive Soldier Differentiation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Nalepa, C.A. Altricial development in wood-feeding cockroaches: The key antecedent of termite eusociality. In Biology of Termites: A Modern Synthesis; Bignell, D.E., Roisin, Y., Lo, N., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 69–96. [Google Scholar]
- Tian, L.; Zhou, X. The soldiers in societies: Defense, regulation, and evolution. Int. J. Biol. Sci. 2014, 10, 296–308. [Google Scholar] [CrossRef] [Green Version]
- Miura, T.; Maekawa, K. The making of the defensive caste: Physiology, development, and evolution of the soldier differentiation in termites. Evol. Dev. 2020, 22, 425–437. [Google Scholar] [CrossRef]
- Myles, T.G. Reproductive soldiers in the Termopsidae (Isoptera). Pan-Pac. Entomol. 1986, 62, 293–299. [Google Scholar]
- Thorne, B.L.; Breisch, N.L.; Muscedere, M.L. Evolution of eusociality and the soldier caste in termites: Influence of intraspecific competition and accelerated inheritance. Proc. Natl. Acad. Sci. USA 2003, 100, 12808–12813. [Google Scholar] [CrossRef] [Green Version]
- Johnson, S.E.; Breisch, N.L.; Momen, B.; Thorne, B.L. Morphology and gonad development of normal soldiers and reproductive soldiers of the termite Zootermopsis nevadensis nevadensis (Isoptera, Archotermopsidae). ZooKeys 2011, 148, 15–30. [Google Scholar]
- Roisin, Y. Diversity and evolution of caste patterns. In Termites: Evolution, Sociality, Symbioses, Ecology; Abe, T., Bignell, D.E., Higashi, M., Eds.; Spinger: Dordrecht, The Netherlands, 2000; pp. 95–119. [Google Scholar]
- Saiki, R.; Yaguchi, H.; Hashimoto, Y.; Kawamura, S.; Maekawa, K. Reproductive soldier-like individuals induced by juvenile hormone analog treatment in Zootermopsis nevadensis (Isoptera, Archotermopsidae). Zool. Sci. 2014, 31, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, D.; Gotoh, H.; Miura, T.; Maekawa, K. Social interactions affecting caste development through physiological actions in termites. Front. Physiol. 2014, 5, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimoji, H.; Oguchi, K.; Hayashi, Y.; Hojo, M.K.; Miura, T. Regulation of neotenic differentiation through direct physical contact in the damp-wood termite Hodotermopsis sjostedti. Insect. Soc. 2017, 64, 393–401. [Google Scholar] [CrossRef]
- Maekawa, K.; Nakamura, S.; Watanabe, D. Termite soldier differentiation in incipient colonies is related to parental proctodeal trophallactic behavior. Zool. Sci. 2012, 29, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Masuoka, Y.; Yaguchi, H.; Suzuki, R.; Maekawa, K. Knockdown of the juvenile hormone receptor gene inhibits soldier-specific morphogenesis in the damp-wood termite Zootermopsis nevadensis (Isoptera: Archotermopsidae). Insect. Biochem. Mol. Biol. 2015, 64, 25–31. [Google Scholar] [CrossRef]
- Yashiro, T.; Mitaka, Y.; Nozaki, T.; Matsuura, K. Chemical and molecular identification of the invasive termite Zootermopsis nevadensis (Isoptera: Archotermopsidae) in Japan. Appl. Entmol. Zool. 2018, 53, 215–221. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015; Available online: https://www.R-project.org (accessed on 10 November 2020).
- Koshikawa, S.; Matsumoto, T.; Miura, T. Morphometric changes during soldier differentiation of the damp-wood termite Hodotermopsis japonica (Isoptera, Termopsidae). Insect. Soc. 2002, 49, 245–250. [Google Scholar] [CrossRef]
- Masuoka, Y.; Miyazaki, S.; Saiki, R.; Tsuchida, T.; Maekawa, K. High Laccase2 expression is likely involved in the formation of specific cuticular structures during soldier differentiation of the termite Reticulitermes speratus. Arthropod. Struct. Dev. 2013, 42, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Itano, H.; Maekawa, K. Soldier differentiation and larval juvenile hormone sensitivity in an incipient colony of the damp-wood termite Zootermopsis nevadensis (Isoptera, Termopsidae). Sociobiology 2008, 51, 151–162. [Google Scholar]
- Simmons, L.W.; Emlen, D.J. Evolutionary trade-off between weapons and testes. Proc. Natl. Acad. Sci. USA 2006, 103, 16346–16351. [Google Scholar] [CrossRef] [Green Version]
- Fry, C.L. Juvenile hormone mediates a trade-off between primary and secondary sexual traits in stalk-eyed flies. Evol. Dev. 2006, 8, 191–201. [Google Scholar] [CrossRef]
- Kelly, C.D. Sperm investment in relation to weapon size in a male trimorphic insect? Behav. Ecol. 2008, 19, 1018–1024. [Google Scholar] [CrossRef] [Green Version]
- Miura, T.; Koshikawa, S.; Matsumoto, T. Winged presoldiers induced by a juvenile hormone analog in Zootermopsis nevadensis: Implications for plasticity and evolution of caste differentiation in termites. J. Morphol. 2003, 257, 22–32. [Google Scholar] [CrossRef]
- Watanabe, D.; Shirasaki, I.; Maekawa, K. Effects of juvenile hormone III on morphogenetic changes during a molt from each nymphal instar in the termite Reticulitermes speratus (Isoptera: Rhinotermitidae). Appl. Entomol. Zool. 2010, 45, 377–386. [Google Scholar] [CrossRef]
- Oguchi, K.; Shimoji, H.; Hayashi, Y.; Miura, T. Reproductive organ development along the caste differentiation pathways in the dampwood termite Hodotermopsis sjostedti. Insect. Soc. 2016, 63, 519–529. [Google Scholar] [CrossRef]
- Watanabe, D.; Gotoh, H.; Miura, T.; Maekawa, K. Soldier presence suppresses presoldier differentiation through a rapid decrease of JH in the termite Reticulitermes speratus. J. Insect. Physiol. 2011, 57, 791–795. [Google Scholar] [CrossRef] [PubMed]
- Penick, C.A.; Trobaugh, B.; Brent, C.S.; Liebig, J. Head-butting as an early indicator of reproductive disinhibition in the termite Zootermopsis nevadensis. J. Insect. Behav. 2013, 26, 23–34. [Google Scholar] [CrossRef]
- Sun, Q.; Haynes, K.F.; Hampton, J.D.; Zhou, X. Sex-specific inhibition and stimulation of worker-reproductive transition in a termite. Naturwissenschaften 2017, 104, 79. [Google Scholar] [CrossRef] [PubMed]
- Funaro, C.F.; Böröczky, K.; Vargo, E.L.; Schal, C. Identification of a queen and king recognition pheromone in the subterranean termite Reticulitermes flavipe. Proc. Natl. Acad. Sci. USA 2018, 115, 3888–3893. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| King Removal Timing | Soldier Removal Timing | Colony Number 1 | Reproductive Soldier Development 2 | Days Until Reproductive Soldier Development (n) | Normal Male Neotenic Development |
|---|---|---|---|---|---|
| - | - | 14 | 0% (0/14) a | - | 0% (0/14) |
| First-soldier emergence | - | 21 | 4.8% (1/21) a | 90.0 (1) | N/A 3 |
| First-soldier emergence | First-soldier emergence | 63 | 3.2% (2/63) a | 91.5 ± 21.5 (2) | N/A 3 |
| +1 month | - | 15 | 33.3% (5/15) ab | 89.4 ± 34.9 (5) | 30% (3/10) 4 |
| +1 month | +1 month | 16 | 25.0% (4/16) ab | 51.0 ± 13.0 (4) | 60% (6/10) 4 |
| +2 months | - | 47 | 19.1% (9/47) a | 58.9 ± 22.1 (9) | 31.9% (15/47) |
| +2 months | +2 months | 47 | 51.1% (24/47) b | 61.2 ± 19.4 (11) | 31.9% (15/47) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masuoka, Y.; Nuibe, K.; Hayase, N.; Oka, T.; Maekawa, K. Reproductive Soldier Development Is Controlled by Direct Physical Interactions with Reproductive and Soldier Termites. Insects 2021, 12, 76. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12010076
Masuoka Y, Nuibe K, Hayase N, Oka T, Maekawa K. Reproductive Soldier Development Is Controlled by Direct Physical Interactions with Reproductive and Soldier Termites. Insects. 2021; 12(1):76. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12010076
Chicago/Turabian StyleMasuoka, Yudai, Keigo Nuibe, Naoto Hayase, Takateru Oka, and Kiyoto Maekawa. 2021. "Reproductive Soldier Development Is Controlled by Direct Physical Interactions with Reproductive and Soldier Termites" Insects 12, no. 1: 76. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12010076