
Analysis of Sanitizer Rotation on the Susceptibility, Biofilm Forming Ability and Caco-2 Cell Adhesion and Invasion of Listeria



Abstract
:1. Introduction
2. Materials and Methods
2.1. Selection of Bacterial Strains and Inoculum Preparation
2.2. Preparation of Sanitizer Solutions
2.3. Single Exposure, Re-Exposure, and Repeated Exposure Treatments
2.4. Biofilm Production Assay
2.5. Caco-2 Cell Interaction Assay
2.5.1. Inoculum Preparation
2.5.2. Preparation of Mammalian Cell Culture
2.5.3. Adhesion and Invasion Assay
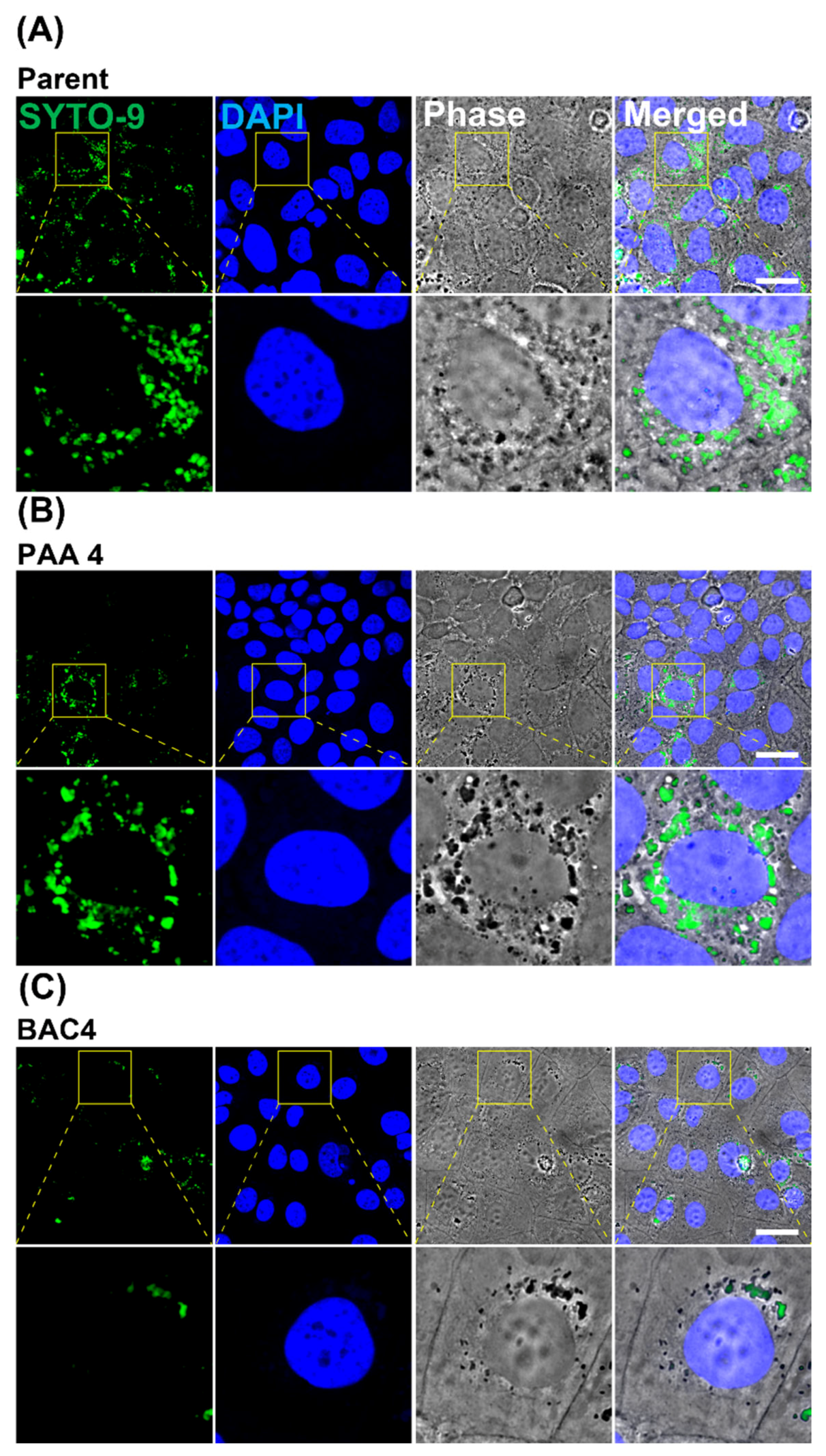
2.5.4. Confocal Imaging of Invaded L. innocua in Caco-2 Cells
2.6. Statistical Analysis
3. Results and Discussion
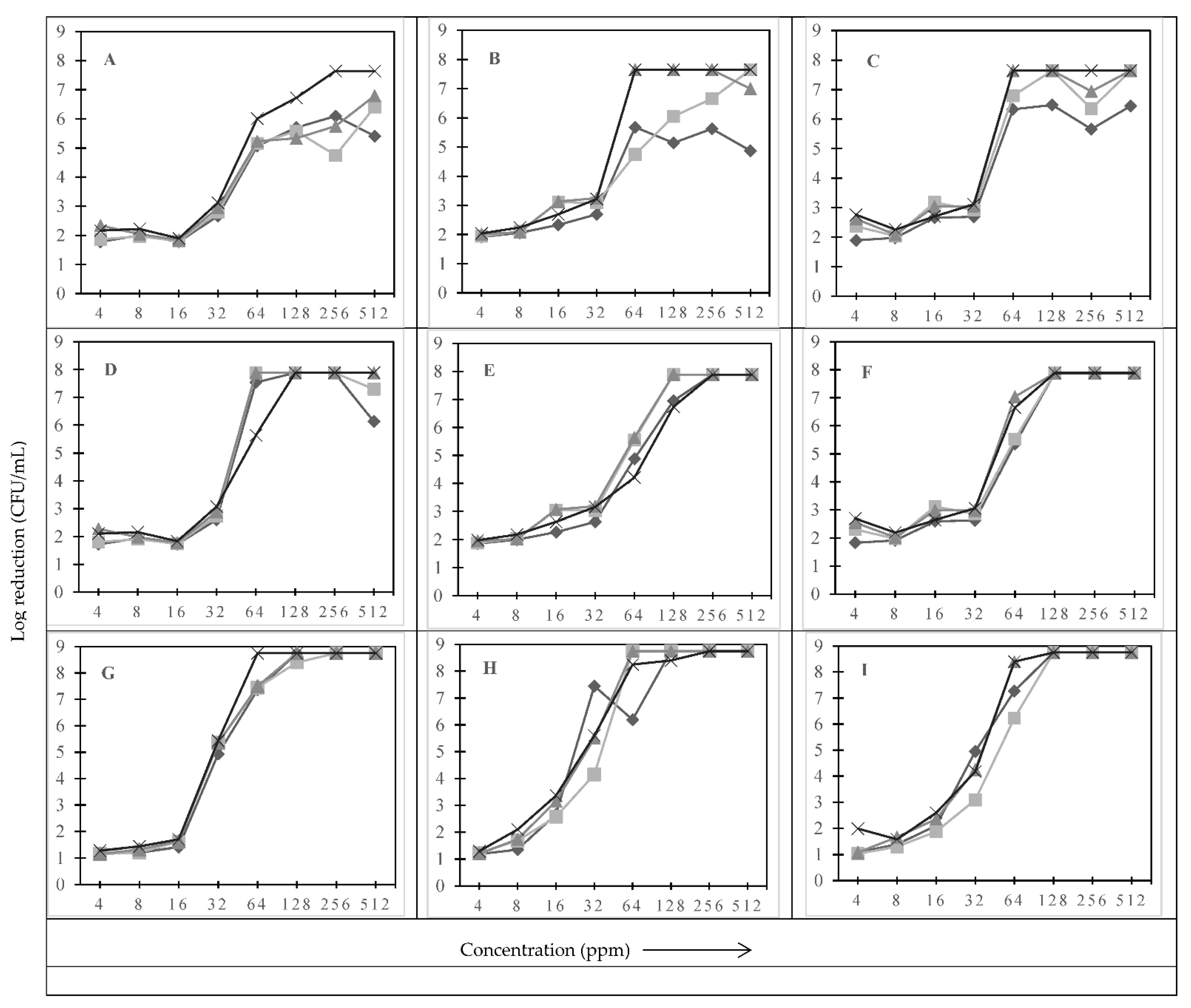
3.1. Effect of Type of Sanitizer, Sanitizer Concentration, and Treatment Time on the Log Reduction
3.2. Effect of Re-Exposure to the Same and/or Different Sanitizers
3.3. Effect of Repeated Sanitizer Exposure on the Survivability of L. monocytogenes and L. innocua
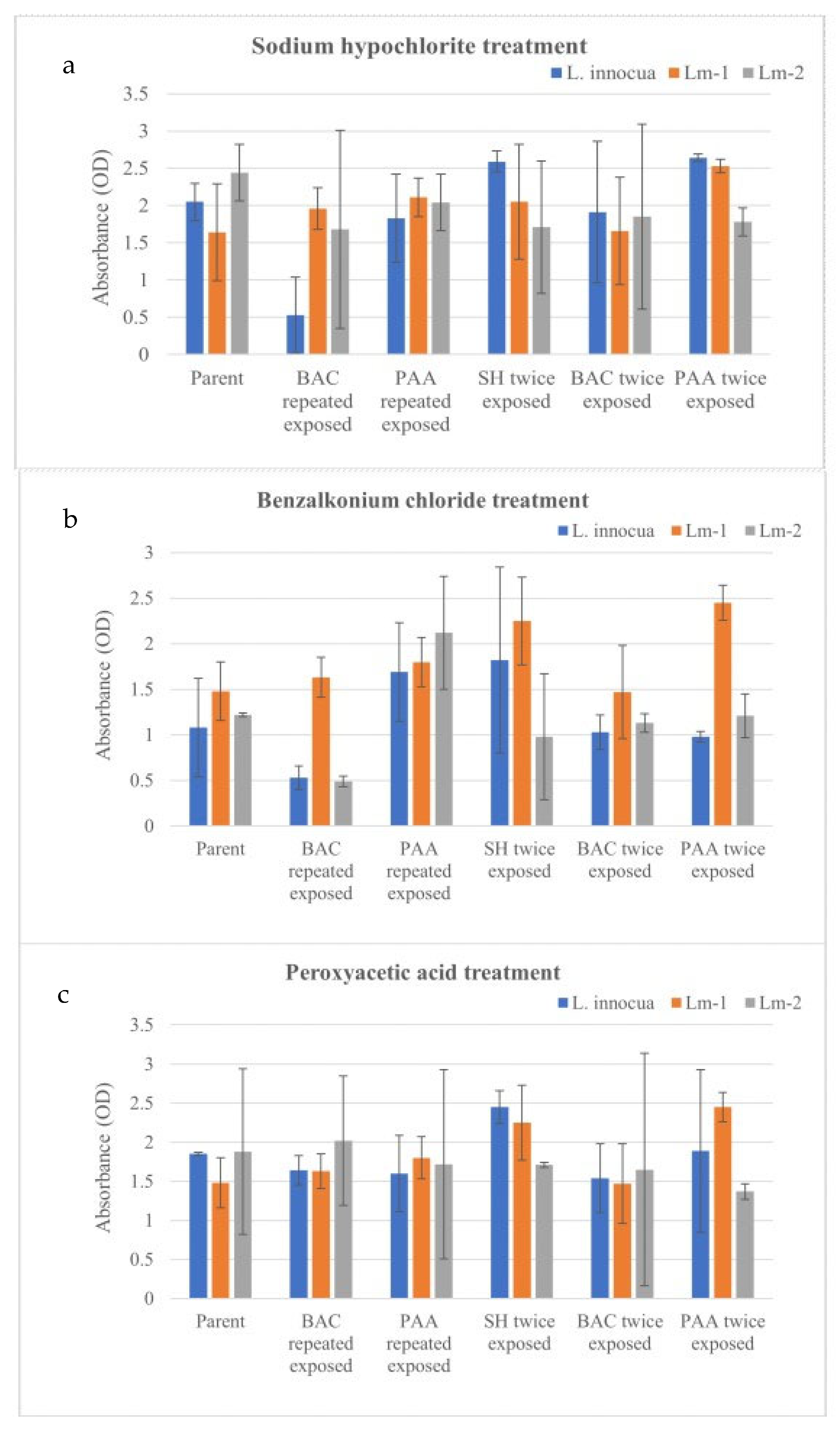
3.4. Effect of Sublethal Exposure to Sanitizers on Biofilm Forming Ability of Listeria spp.
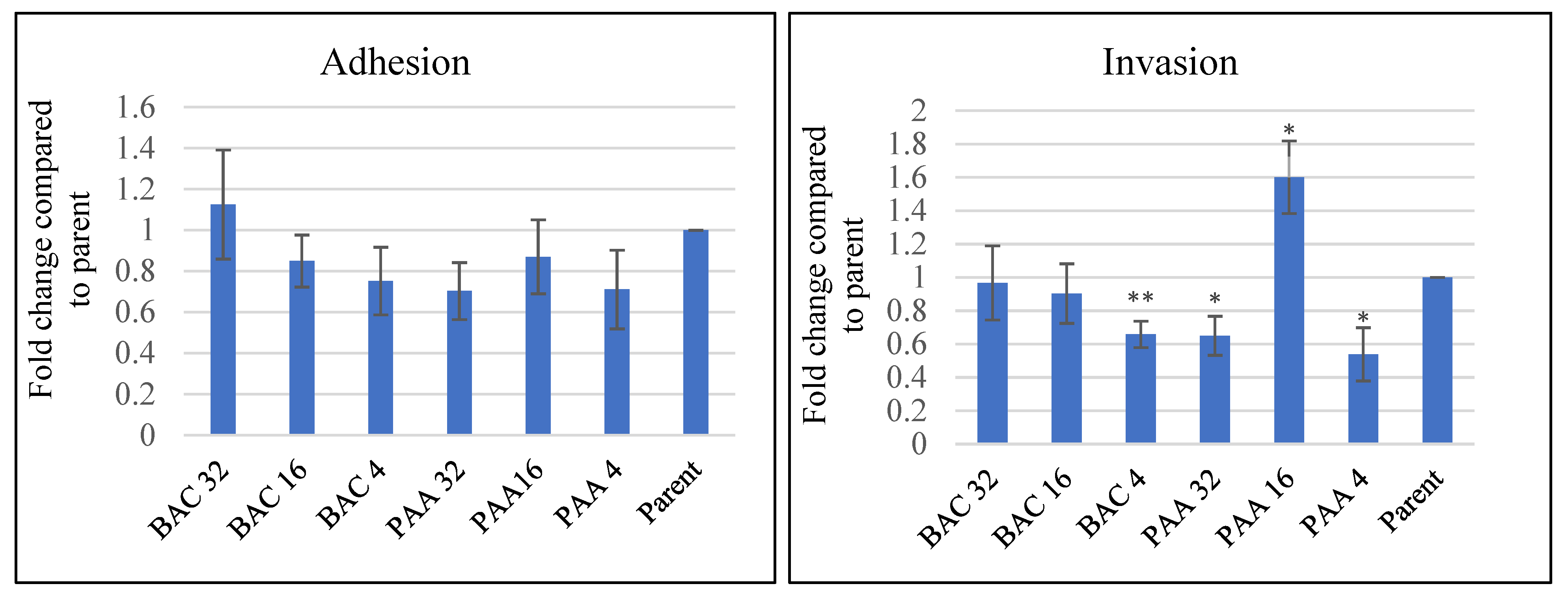
3.5. Effect of Sublethal Sanitizer Exposure on Intestinal Cell Interaction with L. innocua
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Swaminathan, B.; Gerner-Smidt, P. The epidemiology of human listeriosis. Microbes Infect. 2007, 9, 1236–1243. [Google Scholar] [CrossRef] [PubMed]
- Matle, I.; Mbatha, K.R.; Madoroba, E. A review of Listeria monocytogenes from meat and meat products: Epidemiology, virulence factors, antimicrobial resistance, and diagnosis. Onderstepoort J. Vet. Res. 2020, 87, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, V.; Wiedmann, M.; Teixeira, P.; Stasiewicz, M.J. Listeria monocytogenes persistence in food-associated environments: Epidemiology, strain characteristics, and implications for public health. J. Food Prot. 2014, 77, 150–170. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Doyle, M.P.; Zhao, P. Control of Listeria monocytogenes in a Biofilm by competitive-exclusion microorganisms. Appl. Environ. Microbiol. 2004, 70, 3996–4003. [Google Scholar] [CrossRef]
- Adrião, A.; Vieira, M.; Fernandes, I.; Barbosa, M.; Sol, M.; Tenreiro, R.P.; Chambel, L.; Barata, B.; Zilhao, I.; Shama, G.; et al. Marked intra-strain variation in response of Listeria monocytogenes dairy isolates to acid or salt stress and the effect of acid or salt adaptation on adherence to abiotic surfaces. Int. J. Food Microbiol. 2008, 123, 142–150. [Google Scholar] [CrossRef]
- Fox, E.M.; Leonard, N.; Jordan, K. Physiological and transcriptional characterization of persistent and nonpersistent Listeria monocytogenes isolates. Appl. Environ. Microbiol. 2011, 77, 6559–6569. [Google Scholar] [CrossRef]
- Aarnisalo, K.; Sheen, S.; Raaska, L.; Tamplin, M. Modelling transfer of Listeria monocytogenes during slicing of gravad salmon. Int. J. Food Microbiol. 2007, 118, 69–78. [Google Scholar] [CrossRef]
- Leriche, V.; Sibille, P.; Carpentier, B. Use of an enzyme-linked lectin sorbent assay to monitor the shift in polysaccharide composition in bacterial biofilms. Appl. Environ. Microbiol. 2000, 66, 1851–1856. [Google Scholar] [CrossRef]
- Frank, J.F.; Chmielewski, R.A.N. Effectiveness of sanitation with quaternary ammonium compound or chlorine on stainless steel and other domestic food preparation surfaces. J. Food Prot. 1997, 60, 43–47. [Google Scholar] [CrossRef]
- Pan, Y.; Breidt, F.; Kathariou, S. Resistance of Listeria monocytogenes biofilms to sanitizing agents in a simulated food processing environment. Appl. Environ. Microbiol. 2006, 72, 7711–7717. [Google Scholar] [CrossRef] [Green Version]
- Dell’Erba, A.; Falsanisi, D.; Liberti, L.; Notarnicola, M.; Santoroa, D. Disinfection by-products formation during wastewater disinfection with peracetic acid. Desalination 2007, 215, 177–186. [Google Scholar] [CrossRef]
- Schmidt, R.H. Basic Elements of Equipment Cleaning and Sanitizing in Food Processing and Handling Operations; University of Florida Extension Document FS14: Gainesville, FL, USA, 2003. [Google Scholar]
- Waghmare, R.B.; Annapure, U.S. Integrated effect of sodium hypochlorite and modified atmosphere packaging on quality and shelf life of fresh-cut cilantro. Food Packag. Shelf Life 2015, 3, 62–69. [Google Scholar] [CrossRef]
- Ortiz-Sola, J.; Colas-Meda, P.; Nicolau-Lapena, I.; Alegre, I.; Abadias, M.; Vinas, I. Pathogenic potential of the surviving Salmonella Enteritidis on strawberries after disinfection treatments based on ultraviolet-C light and peracetic acid. Int. J. Food Microbiol. 2022, 364, 109536. [Google Scholar] [CrossRef] [PubMed]
- Langsrud, S.; Sundheim, G.; Borgmann-Strahsen, R. Intrinsic and acquired resistsnce to quaternary ammonium compounds in food-related Pseudomonas spp. J. Appl. Microbiol. 2003, 95, 874–882. [Google Scholar] [CrossRef]
- Potenski, C.J.; Gandhi, M.; Matthews, K.R. Exposure of Salmonella Enteritidis to chlorine or food preservatives increases susceptibility to antibiotics. FEMS Microbiol. Lett. 2003, 220, 181–186. [Google Scholar] [CrossRef]
- Russell, A.D. Similarities and differences in the responses of microorganisms to biocides. J. Antimicrob. Chemother. 2003, 52, 750–763. [Google Scholar] [CrossRef]
- Poimenidou, S.V.; Chrysadakou, M.; Tzakoniati, A.; Bikouli, V.C.; Nychas, G.-J.; Skandamis, P.N. Variability of Listeria monocytogenes strains in biofilm formation on stainless steel and polystyrene materials and resistance to peracetic acid and quaternary ammonium compounds. Int. J. Food Microbiol. 2016, 237, 164–171. [Google Scholar] [CrossRef]
- Moradali, M.F.; Ghods, S.; Rehm, B.H. Activation mechanism and cellular localization of membrane-anchored alginate polymerase in Pseudomonas aeruginosa. Appl. Environ. Microbiol. 2017, 83, e03499-16. [Google Scholar] [CrossRef]
- Bland, R.; Waite-Cusic, J.; Weisberg, A.; Riutta, E.; Chang, J.; Kovacevic, J. Adaptation to a commercial quaternary ammonium compound sanitizer leads to cross-resistance to select antibiotics in Listeria monocytogenes isolated from fresh produce environments. Front. Microbiol. 2021, 12, 782920. [Google Scholar] [CrossRef]
- Jeyaletchumi, P.; Tunung, R.; Margaret, S.P.; Son, R.; Ghazali, F.M.; Cheah, Y.K.; Malakar, P.K. Quantification of Listeria monocytogenes in salad vegetables by MPN-PCR. Int. Food Res. J. 2010, 17, 281–286. [Google Scholar]
- Carvalho, F.T.; Vieira, B.S.; Vallim, D.C.; Carvalho, L.A.; Carvalho, R.C.; Pereira, R.C.; Figueiredo, E.E. Genetic similarity, antibiotic resistance and disinfectant susceptibility of Listeria monocytogenes isolated from chicken meat and chicken-meat processing environment in Mato Grosso, Brazil. LWT 2019, 109, 77–82. [Google Scholar] [CrossRef]
- Vazquez-Boland, J.A.; Kuhn, M.; Berche, P.; Chakraborty, T.; Dominguez-Bernal, G.; Goebel, W.; Gonzalez-Zorn, B.; Wehland, J.; Kreft, J. Listeria pathogenesis and molecular virulence determinants. Clin. Microbiol. Rev. 2001, 14, 584–640. [Google Scholar] [CrossRef] [PubMed]
- CLSI—Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically. CLSI Document M7-A8, Approved Standard, 8th ed.; CLSI: Wayne, PA, USA, 2009; ISBN 1-56238-689-1. [Google Scholar]
- Riazi, S.; Matthews, K.R. Failure of foodborne pathogens to develop resistance to sanitizers following repeated exposure to common sanitizers. Int. Biodeterior. Biodegrad. 2011, 65, 374–378. [Google Scholar] [CrossRef]
- Ammendolia, M.G.; Iosi, F.; De Berardis, B.; Guccione, G.; Superti, F.; Conte, M.P.; Longhi, C. Listeria monocytogenes behaviour in presence of non-UV-irradiated titanium dioxide nanoparticles. PLoS ONE 2014, 9, e84986. [Google Scholar] [CrossRef]
- Reddy, S.; Austin, F. Adhesion and invasion assay procedure using Caco-2 cells for Listeria monocytogenes. Bio Protoc. 2017, 7, e2267. [Google Scholar] [CrossRef] [PubMed]
- Belessi, C.E.A.; Gounadaki, A.S.; Psomas, A.N.; Skandamis, P.N. Efficiency of different sanitation methods on Listeria monocytogenes biofilms formed under various environmental conditions. Int. J. Food Microbiol. 2011, 145, S46–S52. [Google Scholar] [CrossRef] [PubMed]
- Tamburro, M.; Ripabelli, G.; Vitullo, M.; Dallman, T.J.; Pontello, M.; Amar, C.F.L.; Sammarco, M.L. Gene expression in Listeria monocytogenes exposed to sublethal concentration of benzalkonium chloride. Comp. Immunol. Microbiol. Infect. Dis. 2015, 40, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, S. Understanding the contribution of environmental factors in the spread of antimicrobial resistance. Environ. Health Prev. Med. 2015, 20, 243–252. [Google Scholar] [CrossRef]
- Bland, R.; Brown, S.R.B.; Waite-Cusic, J.; Kovacevic, J. Probing antimicrobial resistance and sanitizer tolerance themes and their implications for the food industry through the Listeria monocytogenes lens. Compr. Rev. Food Sci. Food Saf. 2021, 21, 1777–1802. [Google Scholar] [CrossRef]
- Warth, A.D. Mechanism of action of benzoic acid on Zygosaccharomyces bailii: Effects on glycolytic metabolite levels, energy production, and intracellular pH. Appl. Environ. Microbiol. 1991, 57, 3410–3414. [Google Scholar] [CrossRef]
- Osek, J.; Lachtara, B.; Wieczorek, K. Listeria monocytogenes—How this pathogen survives in food-production environments? Front. Microbiol. 2022, 13, 866462. [Google Scholar] [CrossRef] [PubMed]
- Gullberg, E.; Albrecht, L.M.; Karlsson, C.; Sandegren, L.; Andersson, D.I. Selection of a multidrug resistance plasmid by sublethal levels of antibiotics and heavy metals. MBio 2014, 5, e01918-14. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, J.; Bland, R.; Waite-Cusic, J.; Kovacevic, J. Diversity and antimicrobial resistance of Listeria spp. and L. monocytogenes clones from produce handling and processing facilities in the Pacific Northwest. Food Control 2021, 123, 107665. [Google Scholar] [CrossRef]
- Li, J.; Xie, S.; Ahmed, S.; Wang, F.; Gu, Y.; Zhang, C.; Chai, X.; Wu, Y.; Cai, J.; Cheng, G. Antimicrobial activity and resistance: Influencing factors. Front. Pharmacol. 2017, 8, 364. [Google Scholar] [CrossRef]
- Gao, H.; Liu, C. Biochemical and morphological alteration of Listeria monocytogenes under environmental stress caused by chloramine-T and sodium hypochlorite. Food Control 2014, 46, 455–461. [Google Scholar] [CrossRef]
- Kostaki, M.; Chorianopoulos, N.; Braxou, E.; Nychas, G.J.; Giaouris, E. Differential biofilm formation and chemical disinfection resistance of sessile cells of Listeria monocytogenes strains under monospecies and dual-species (with Salmonella enterica) conditions. Appl. Environ. Microbiol. 2012, 78, 2586–2595. [Google Scholar] [CrossRef]
- Sharma, D.; Misba, L.; Khan, A.U. Antibiotics versus biofilm: An emerging battleground in microbial communities. Antimicrob. Resist. Infect. Control 2019, 8, 76. [Google Scholar] [CrossRef]
- Luque-Sastre, L.; Fox, E.M.; Jordan, K.; Fanning, S. Comparative study of the susceptibility of Listeria species to sanitizer treatments when grown under planktonic and biofilm conditions. J. Food Prot. 2018, 81, 1481–1490. [Google Scholar] [CrossRef]
- Hua, Z.; Korany, A.M.; El-Shinawy, S.H.; Zhu, M.-J. Comparative evaluation of different sanitizers against Listeria monocytogenes biofilms on major food-contact surfaces. Front. Microbiol. 2019, 10, 2462. [Google Scholar] [CrossRef]
- Feliciano, L.; Lee, J.; Pacall, M.A. Transmission electron microscopic analysis showing structural changes to bacterial cells treated with electrolyzed water and an acidic sanitizer. J. Food Sci. 2012, 77, M182–M187. [Google Scholar] [CrossRef]
- McGoverin, C.; Robertson, J.; Jonmohamadi, Y.; Swift, S.; Vanholsbeeck, F. Species dependence of SYTO 9 staining of bacteria. Front. Microbiol. 2020, 11, 545419. [Google Scholar] [CrossRef] [PubMed]
- Meunier, V.; Bourrie, M.; Berger, Y.; Fabre, G. The human intestinal epithelial cell line Caco-2; pharmacological and pharmacokinetic applications. Cell Biol. Toxicol. 1995, 11, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Jaradat, Z.W.; Bhunia, A.K. Adhesion, invasion, and translocation characteristics of Listeria monocytogenes serotypes in Caco-2 cell and mouse models. Appl. Environ. Microbiol. 2003, 69, 3640–3645. [Google Scholar] [CrossRef] [PubMed]
- Moroni, O.; Kheadr, E.; Boutin, Y.; Lacroix, C.; Fliss, I. Inactivation of adhesion and invasion of food-borne Listeria monocytogenes by bacteriocin-producing Bifidobacterium strains of human origin. Appl. Environ. Microbiol. 2006, 72, 6894–6901. [Google Scholar] [CrossRef]
- Ortiz, S.; López, V.; Martínez-Suárez, J. The influence of subminimal inhibitory concentrations of benzalkonium chloride on biofilm formation by Listeria monocytogenes. Int. J. Food Microbiol. 2014, 189, 106–112. [Google Scholar] [CrossRef]
 represent treatment times of 30 s, 1, 2.5, and 5 min, respectively.
represent treatment times of 30 s, 1, 2.5, and 5 min, respectively.
represent treatment times of 30 s, 1, 2.5, and 5 min, respectively.
represent treatment times of 30 s, 1, 2.5, and 5 min, respectively.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sanitizer Concentration (ppm) | Treatment Time (min) | Log Reduction (CFU/mL) of L. innocua when Exposed to Same or Different Sanitizers either Once (Parent) or Repeatedly (Survived Cells Re-Exposed) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sodium Hypochlorite (SHC) Treatment | Peroxyacetic Acid (PAA) Treatment | Benzalkonium Chloride (BAC) Treatment | |||||||||||
| 1 Parent | 2 SHC Exposed | 3 BAC Exposed | 4 PAA Exposed | 5 Parent | 6 SHC Exposed | 7 BAC Exposed | 8 PAA Exposed | 9 Parent | 10 SHC Exposed | 11 BAC Exposed | 12 PAA Exposed | ||
| 512 | 0.5 | 5.41 ± 1.05 aA | 4.32 ± 1.67 aA | 4.49 ± 0.35 aA | 7.66 ± 0.89 bB | 6.45 ± 0.21 bB | 5.58 ± 0.21 aA | 4.89 ± 0.21 aA | 7.66 ± 0.28 bB | 4.88 ± 0.5 aA | 6.08 ± 0.01 bB | 5.08 ± 0.01 aA | 7.66 ± 1.04 cB |
| 1 | 6.41 ± 0.35 bB | 4.93 ± 0.35 aA | 5.37 ± 0.35 aA | 7.66 ± 0.35 cB | 7.65 ± 0.21 bB | 5.31 ± 0.21 aA | 5.37 ± 0.21 aA | 7.66 ± 0.62 bB | 7.65 ± 0.01 bB | 6.08 ± 0.01 aB | 5.22 ± 0.01 aA | 7.66 ± 0.23 bB | |
| 2.5 | 6.80 ± 0.30 aB | 6.08 ± 1.1 aB | 6.26 ± 0.09 aB | 7.66 ± 0.81 bB | 7.65 ± 0.21 bB | 6.08 ± 0.21 aB | 5.56 ± 0.21 aA | 7.66 ± 0.28 bB | 7.00 ± 0.01 bB | 5.73 ± 0.01 aA | 5.32 ± 0.01 aA | 7.66 ± 0.01 bB | |
| 5 | 7.65 ± 1.18 bB | 6.08 ± 0.35 aB | 6.26 ± 0.35 aB | 7.66 ± 0.35 bB | 7.65 ± 0.21 bB | 6.08 ± 0.21 aB | 6.26 ± 0.21 aB | 7.66 ± 0.21 bB | 7.65 ± 0.01 bB | 6.08 ± 0.01 aB | 5.56 ± 0.00 aA | 7.66 ± 0.83 bB | |
| 256 | 0.5 | 6.10 ± 0.24 bB | - | 4.41 ± 1.30 aA | 7.66 ± 0.40 bB | 5.66 ± 0.21 aA | 5.73 ± 0.21 aA | 4.46 ± 0.21 aA | - | 5.63 ± 0.01 aA | 6.08 ± 0.49 aB | - | 7.66 ± 0.09 bB |
| 1 | 4.75 ± 1.03 aA | - | 6.26 ± 0.35 bB | 7.66 ± 0.10 bB | 6.36 ± 0.21 aB | 5.73 ± 0.28 aA | 5.46 ± 0.21 aA | - | 6.66 ± 0.01 aB | 6.08 ± 1.14 aB | - | 7.66 ± 0.10 bB | |
| 2.5 | 5.75 ± 0.33 aA | - | 6.26 ± 1.51 bB | 7.66 ± 0.68 bB | 6.95 ± 2.06 aB | 6.08 ± 1.64 aB | 5.52 ± 0.71 aA | - | 7.65 ± 0.50 bB | 6.08 ± 0.86 aB | - | 7.66 ± 0.27 bB | |
| 5 | 7.65 ± 1.15 aB | - | 6.26 ± 0.35 aB | 7.66 ± 0.16 aB | 7.65 ± 0.21 bB | 5.25 ± 0.49 aA | 6.26 ± 0.28 aB | - | 7.65 ± 1.54 bB | 6.08 ± 0.76 aB | - | 7.66 ± 0.72 bB | |
| 128 | 0.5 | 5.70 ± 0.03 aA | - | 4.45 ± 0.17 aA | 7.16 ± 0.14 bB | 6.48 ± 2.75 bB | 4.37 ± 2.55 aA | 4.54 ± 0.62 aA | - | 5.14 ± 0.03 aA | 5.06 ± 0.46 aA | - | 6.66 ± 1.08 aB |
| 1 | 5.58 ± 0.01 aA | - | 6.26 ± 0.05 aB | 7.66 ± 0.19 bB | 7.65 ± 2.63 bB | 4.32 ± 2.55 aA | 4.76 ± 0.28 aA | - | 6.05 ± 0.04 aB | 5.58 ± 0.16 aA | - | 7.66 ± 0.21 bB | |
| 2.5 | 5.34 ± 0.08 aA | - | 6.26 ± 0.78 aB | 7.66 ± 0.06 bB | 7.65 ± 0.43 bB | 4.25 ± 0.87 aA | 5.33 ± 1.69 aA | - | 7.65 ± 0.14 bB | 4.93 ± 0.74 aA | - | 7.66 ± 0.21 bB | |
| 5 | 6.72 ± 0.02 aB | - | 6.26 ± 0.15 aB | 7.66 ± 0.01 bB | 7.65 ± 0.39 bB | 4.35 ± 1.41 aA | 5.67 ± 1.83 aA | - | 7.65 ± 0.08 bB | 4.93 ± 0.11 aA | - | 7.66 ± 0.21 bB | |
| 64 | 0.5 | 5.07 ± 0.12 aA | - | 4.02 ± 0.04 aA | 6.25 ± 0.01 aB | 6.34 ± 0.86 bB | 3.55 ± 1.15 aA | 4.4 ± 0.62 aA | - | 5.68 ± 0.18 aA | 4.46 ± 0.20 aA | - | 5.23 ± 0.21 aA |
| 1 | 5.16 ± 0.04 aA | - | 5.26 ± 0.21 aA | 7.66 ± 0.01 bB | 6.8b ± 0.60 bB | 3.75 ± 1.17 aA | 5.33 ± 0.9 aA | - | 4.75 ± 0.02 aA | 4.38 ± 0.25 aA | - | 6.36 ± 0.21 bB | |
| 2.5 | 5.23 ± 0.01 aA | - | 6.26 ± 0.66 aB | 7.66 ± 0.01 bB | 7.65 ± 0.95 bB | 4.23 ± 0.89 aA | 6.26 ± 1.05 bB | - | 7.65 ± 0.07 bB | 3.43 ± 0.12 aA | - | 7.66 ± 0.21 bB | |
| 5 | 6.01 ± 0.23 aB | - | 6.26 ± 1.03 aB | 7.66 ± 0.18 bB | 7.65 ± 0.77 bB | 4.44 ± 0.91 aA | 6.26 ± 2.14 bB | - | 7.65 ± 0.29 bB | 4.88 ± 0.06 aA | - | 7.66 ± 0.71 bB | |
| Type of Sanitizer | Concentration of Sanitizer (ppm) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Strains | 1 | 2 | 4 | 8 | 16 | 32 | 64 | 128 | |
| BAC | L. innocua | + | + | + | + | + | + | − | − |
| Lm-1 | + | + | + | + | + | + | − | − | |
| Lm-2 | + | + | + | + | + | + | − | − | |
| PAA | L. innocua | + | + | + | + | + | + | + | − |
| Lm-1 | + | + | + | + | + | + | + | − | |
| Lm-2 | + | + | + | + | + | + | + | − | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahman, M.A.; Sahoo, N.; Yemmireddy, V. Analysis of Sanitizer Rotation on the Susceptibility, Biofilm Forming Ability and Caco-2 Cell Adhesion and Invasion of Listeria. Pathogens 2022, 11, 961. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11090961
Rahman MA, Sahoo N, Yemmireddy V. Analysis of Sanitizer Rotation on the Susceptibility, Biofilm Forming Ability and Caco-2 Cell Adhesion and Invasion of Listeria. Pathogens. 2022; 11(9):961. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11090961
Chicago/Turabian StyleRahman, Md Asfakur, Nirakar Sahoo, and Veerachandra Yemmireddy. 2022. "Analysis of Sanitizer Rotation on the Susceptibility, Biofilm Forming Ability and Caco-2 Cell Adhesion and Invasion of Listeria" Pathogens 11, no. 9: 961. https://0-doi-org.brum.beds.ac.uk/10.3390/pathogens11090961