
Nitric Oxide Induced stx2 Expression Is Inhibited by the Nitric Oxide Reductase, NorV, in a Clade 8 Escherichia coli O157:H7 Outbreak Strain


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, norV Detection, and Growth Conditions
2.2. Construction of a norV Mutant
2.3. Bacterial Gene Expression
2.4. Cell Culture and EHEC Infection
3. Results
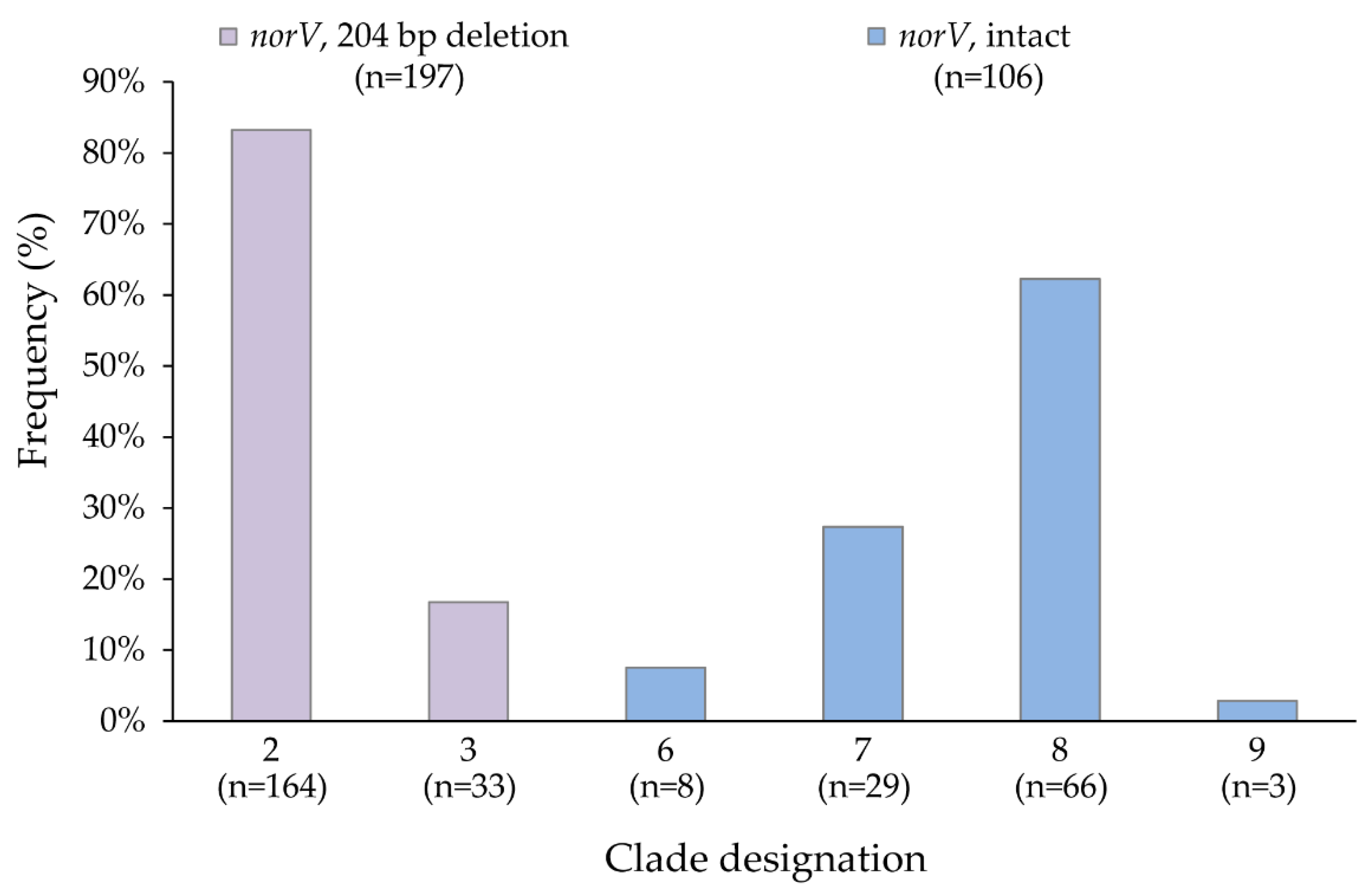
3.1. norV Distribution by Clade and Association with Stx Gene Profiles
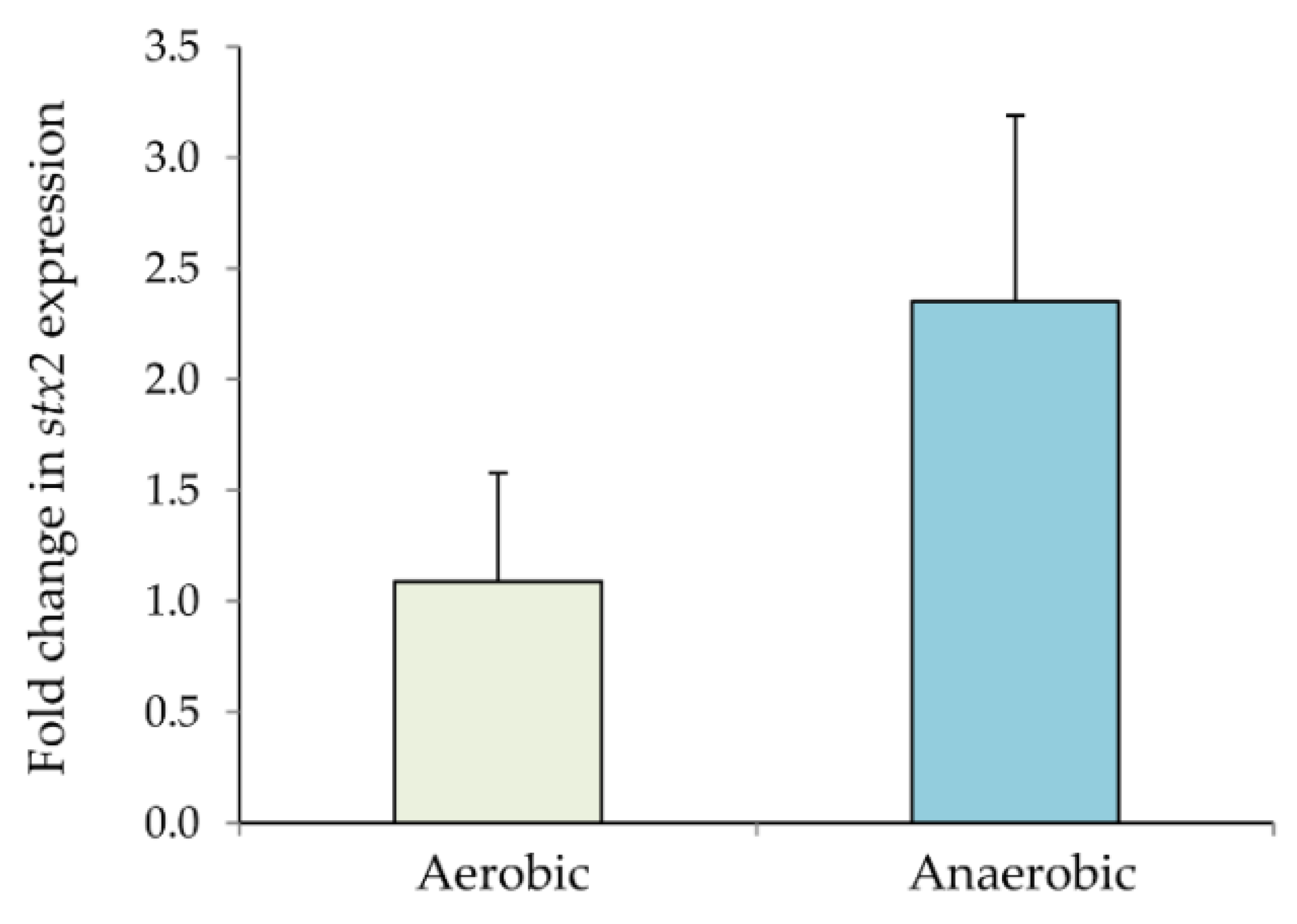
3.2. Impact of NO on stx2 Expression in the Clade 8 Outbreak Strain
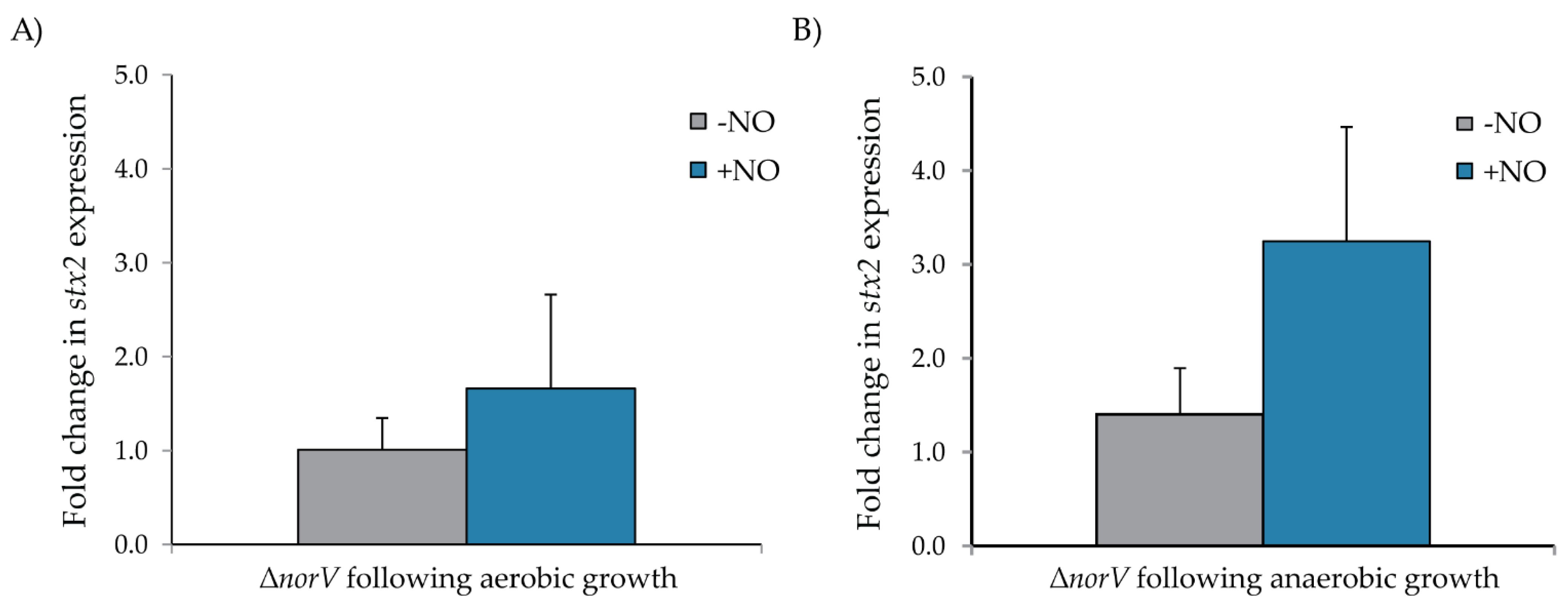
3.3. Role of norV in stx2 Expression in the Clade 8 Outbreak Strain across Growth Conditions
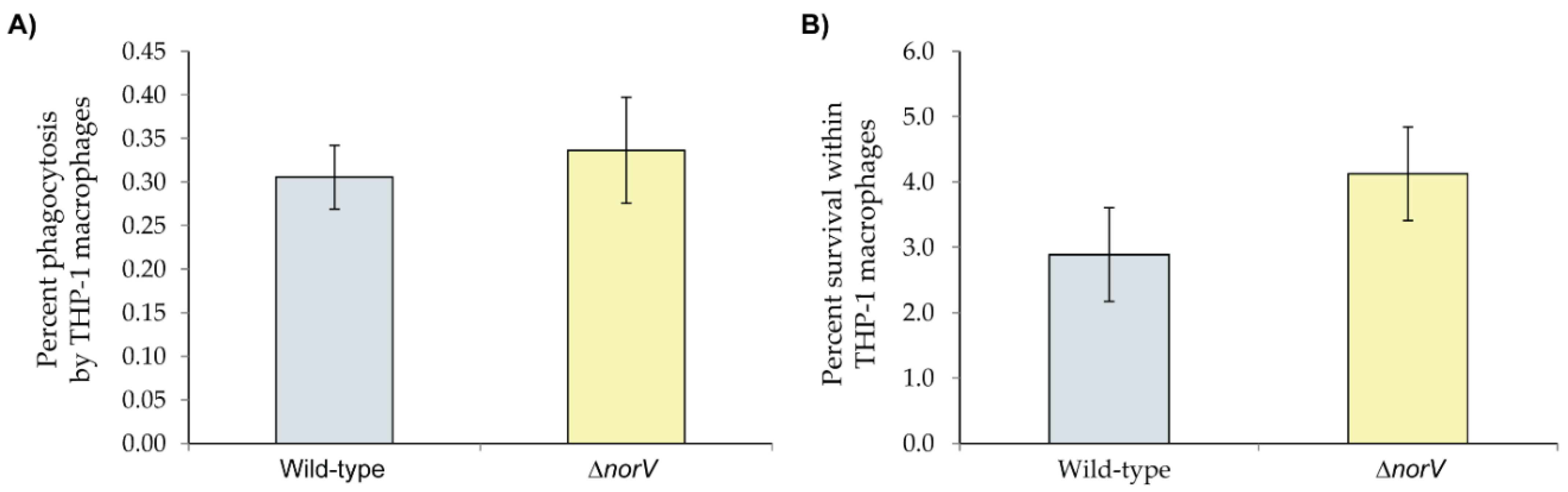
3.4. Role of norV in Uptake and Survival of the Clade 8 Outbreak Strain within Macrophages
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karmali, M.A.; Petric, M.; Lim, C.; McKeough, P.C.; Arbus, G.S.; Lior, H. The Association between Idiopathic Hemolytic Uremic Syndrome and Infection by Verotoxin-Producing Escherichia coli. J. Infect. Dis. 1985, 151, 775–782. [Google Scholar] [CrossRef]
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.A.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne Illness Acquired in the United States-Major Pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef]
- Tack, D.M.; Marder, E.P.; Griffin, P.M.; Cieslak, P.R.; Dunn, J.; Hurd, S.; Scallan, E.; Lathrop, S.; Muse, A.; Ryan, P.; et al. Preliminary Incidence and Trends of Infections with Pathogens Transmitted Commonly Through Food—Foodborne Diseases Active Surveillance Network, 10 U.S. Sites, 2015–2018. Morb. Mortal. Wkly. Rep. 2019, 68, 369–373. [Google Scholar] [CrossRef] [Green Version]
- Centers for Disease Control and Prevention. In National Shiga Toxin-Producing Escherichia coli (STEC) Surveillance Annual Report, 2016; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2018. Available online: www.cdc.gov/ecoli/surv2016/index.html (accessed on 30 December 2020).
- Strockbine, N.A.; Marques, L.R.M.; Newland, J.W.; Smith, H.W.; Holmes, R.K.; O’brien, A.D. Two Toxin-Converting Phages from Escherichia coli O157:H7 Strain 933 Encode Antigenically Distinct Toxins with Similar Biologic Activities. Infect. Immun. 1986, 53, 135–140. [Google Scholar] [CrossRef] [Green Version]
- McDaniel, T.K.; Kaper, J.B. A Cloned Pathogenicity Island from Enteropathogenic Escherichia coli Confers the Attaching and Effacing Phenotype on E. coli K-12. Mol. Microbiol. 1997, 23, 399–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nataro, J.P.; Kaper, J.B. Diarrheagenic Escherichia coli. Clin. Microbiol. Rev. 1998, 11, 142–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostroff, S.M.; Tarr, P.I.; Neill, M.A.; Lewis, J.H.; Hargrett-Bean, N.; Kobayashi, J.M. Toxin Genotypes and Plasmid Profiles as Determinants of Systemic Sequelae in Escherichia coli O157:H7 Infections. J. Infect. Dis. 1989, 160, 994–998. [Google Scholar] [CrossRef] [PubMed]
- Orth, D.; Grif, K.; Khan, A.B.; Naim, A.; Dierich, M.P.; Würzner, R. The Shiga Toxin Genotype Rather than the Amount of Shiga Toxin or the Cytotoxicity of Shiga Toxin in vitro Correlates with the Appearance of the Hemolytic Uremic Syndrome. Diagn. Microbiol. Infect. Dis. 2007, 59, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Manning, S.D.; Motiwala, A.S.; Springman, A.C.; Qi, W.; Lacher, D.W.; Ouellette, L.M.; Mladonicky, J.M.; Somsel, P.; Rudrik, J.T.; Dietrich, S.E.; et al. Variation in Virulence among Clades of Escherichia coli O157:H7 Associated with Disease Outbreaks. Proc. Natl. Acad. Sci. USA 2008, 105, 4868–4873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu-Ali, G.S.; Ouellette, L.M.; Henderson, S.T.; Lacher, D.W.; Riordan, J.T.; Whittam, T.S.; Manning, S.D. Increased Adherence and Expression of Virulence Genes in a Lineage of Escherichia coli O157:H7 Commonly Associated with Human Infections. PLoS ONE 2010, 5, e10167. [Google Scholar] [CrossRef] [PubMed]
- Henderson, S.T.; Singh, P.; Knupp, D.; Lacher, D.W.; Abu-Ali, G.S.; Rudrik, J.T.; Manning, S.D. Variability in the Occupancy of Escherichia coli O157 Integration Sites by Shiga Toxin-Encoding Prophages. Toxins 2021, 13, 433. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Ongoing Multistate Outbreak of Escherichia coli Serotype O157:H7 Infections Associated with Consumption of Fresh Spinach. Morb. Mortal. Wkly. Rep. 2006, 55, 1–2. [Google Scholar]
- Kulasekara, B.R.; Jacobs, M.; Zhou, Y.; Wu, Z.; Sims, E.; Saenphimmachak, C.; Rohmer, L.; Ritchie, J.M.; Radey, M.; McKevitt, M.; et al. Analysis of the Genome of the Escherichia coli O157:H7 2006 Spinach-Associated Outbreak Isolate Indicates Candidate Genes that May Enhance Virulence. Infect. Immun. 2009, 77, 3713–3721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, A.M.; Helmick, R.A.; Gardner, P.R. Flavorubredoxin, an Inducible Catalyst for Nitric Oxide Reduction and Detoxification in Escherichia coli. J. Biol. Chem. 2002, 277, 8172–8177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perna, N.T.; Plunkett, G.; Burland, V.; Mau, B.; Glasner, J.D.; Rose, D.J.; Mayhew, G.F.; Evans, P.S.; Gregor, J.; Kirkpatrick, H.A.; et al. Genome Sequence of Enterohaemorrhagic Escherichia coli O157:H7. Nature 2001, 409, 529–533. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, T.; Makino, K.; Ohnishi, M.; Kurokawa, K.; Ishii, K.; Yokoyama, K.; Han, C.G.; Ohtsubo, E.; Nakayama, K.; Murata, T.; et al. Complete Genome Sequence of Enterohemorrhagic Escherichia coli O157:H7 and Genomic Comparison with a Laboratory Strain K-12. DNA Res. 2001, 8, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimizu, T.; Tsutsuki, H.; Matsumoto, A.; Nakaya, H.; Noda, M. The Nitric Oxide Reductase of Enterohaemorrhagic Escherichia coli Plays an Important Role for the Survival within Macrophages. Mol. Microbiol. 2012, 85, 492–512. [Google Scholar] [CrossRef]
- Manning, S.D.; Madera, R.T.; Schneider, W.; Dietrich, S.E.; Khalife, W.; Brown, W.; Whittam, T.S.; Somsel, P.; Rudrik, J.T. Surveillance for Shiga Toxin-Producing Escherichia coli, Michigan, 2001–2005. Emerg. Infect. Dis. 2007, 13, 318. [Google Scholar] [CrossRef]
- Murphy, K.C.; Campellone, K.G. Lambda Red-Mediated Recombinogenic Engineering of Enterohemorrhagic and Enteropathogenic E. Coli. BMC Mol. Biol. 2003, 4, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanaja, S.K.; Bergholz, T.M.; Whittam, T.S. Characterization of the Escherichia coli O157:H7 Sakai GadE Regulon. J. Bacteriol. 2009, 191. [Google Scholar] [CrossRef] [Green Version]
- Al Safadi, R.; Abu-Ali, G.S.; Sloup, R.E.; Rudrik, J.T.; Waters, C.M.; Eaton, K.A.; Manning, S.D. Correlation between in vivo Biofilm Formation and Virulence Gene Expression in Escherichia coli O104:H4. PLoS ONE. 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Datsenko, K.A.; Wanner, B.L. One-Step Inactivation of Chromosomal Genes in Escherichia coli K-12 Using PCR Products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef] [Green Version]
- Bergholz, T.M.; Wick, L.M.; Qi, W.; Riordan, J.T.; Ouellette, L.M.; Whittam, T.S. Global Transcriptional Response of Escherichia coli O157:H7 to Growth Transitions in Glucose Minimal Medium. BMC Microbiol. 2007, 7, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative CT Method. Nat. Prot. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Schwende, H.; Fitzke, E.; Ambs, P.; Dieter, P. Differences in the State of Differentiation of THP-1 Cells Induced by Phorbol Ester and 1,25-Dihydroxyvitamin D3. J. Leukoc. Biol. 1996, 59, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Korir, M.L.; Laut, C.; Rogers, L.M.; Plemmons, J.A.; Aronoff, D.M.; Manning, S.D. Differing Mechanisms of Surviving Phagosomal Stress among Group B Streptococcus Strains of Varying Genotypes. Virulence 2017, 8, 924–937. [Google Scholar] [CrossRef] [Green Version]
- Zangari, T.; Melton-Celsa, A.R.; Panda, A.; Smith, M.A.; Tatarov, I.; de Tolla, L.; O’Brien, A.D. Enhanced Virulence of the Escherichia coli O157:H7 Spinach-Associated Outbreak Strain in Two Animal Models Is Associated with Higher Levels of Stx2 Production after Induction with Ciprofloxacin. Infect. Immun. 2014, 82, 4968–4977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaikh, N.; Tarr, P.I. Escherichia coli O157:H7 Shiga Toxin-Encoding Bacteriophages: Integrations, Excisions, Truncations, and Evolutionary Implications. J. Bacteriol. 2003, 185, 3596–3605. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, T.; Hirai, S.; Yokoyama, E.; Ichimura, K.; Noda, M. An Evolutionary Analysis of Nitric Oxide Reductase Gene NorV in Enterohemorrhagic Escherichia coli O157. Infect. Genet. Evol. 2015, 33, 176–181. [Google Scholar] [CrossRef]
- Gardette, M.; Daniel, J.; Loukiadis, E.; Jubelin, G. Role of the Nitric Oxide Reductase NorVW in the Survival and Virulence of Enterohaemorrhagic Escherichia coli during Infection. Pathogens 2020, 9, 683. [Google Scholar] [CrossRef]
- Flint, A.; Butcher, J.; Stintzi, A. Stress Responses, Adaptation, and Virulence of Bacterial Pathogens during Host Gastrointestinal Colonization. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Vareille, M.; de Sablet, T.; Hindré, T.; Martin, C.; Gobert, A.P. Nitric Oxide Inhibits Shiga-Toxin Synthesis by Enterohemorrhagic Escherichia coli. Proc. Natl. Acad. Sci. USA 2007, 104, 10199–10204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branchu, P.; Matrat, S.; Vareille, M.; Garrivier, A.; Durand, A.; Crépin, S.; Harel, J.; Jubelin, G.; Gobert, A.P. NsrR, GadE, and GadX Interplay in Repressing Expression of the Escherichia coli O157:H7 LEE Pathogenicity Island in Response to Nitric Oxide. PLoS Pathog. 2014, 10, e1003874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ichimura, K.; Shimizu, T.; Matsumoto, A.; Hirai, S.; Yokoyama, E.; Takeuchi, H.; Yahiro, K.; Noda, M. Nitric Oxide-Enhanced Shiga Toxin Production Was Regulated by Fur and RecA in Enterohemorrhagic Escherichia coli O157. Microbiol. Open 2017, 6, e00461. [Google Scholar] [CrossRef]
- McGannon, C.M.; Fuller, C.A.; Weiss, A.A. Different Classes of Antibiotics Differentially Influence Shiga Toxin Production. Antimicrob. Agents Chemother. 2010, 54, 3790–3798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacMicking, J.; Xie, Q.W.; Nathan, C. Nitric Oxide and Macrophage Function. Annu. Rev. Immunol. 1997, 15, 323–350. [Google Scholar] [CrossRef]
- Gardner, A.M.; Gardner, P.R. Flavohemoglobin Detoxifies Nitric Oxide in Aerobic, but Not Anaerobic, Escherichia coli: Evidence for a Novel Inducible Anaerobic Nitric Oxide-Scavenging Activity. J. Biol. Chem. 2002, 277. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Vine, C.E.; Balasiny, B.K.; Rizk, J.; Bradley, C.L.; Tinajero-Trejo, M.; Poole, R.K.; Bergaust, L.L.; Bakken, L.R.; Cole, J.A. The Roles of the Hybrid Cluster Protein, Hcp and Its Reductase, Hcr, in High Affinity Nitric Oxide Reduction That Protects Anaerobic Cultures of Escherichia coli against Nitrosative Stress. Mol. Microbiol. 2016, 100. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, T.; Matsumoto, A.; Noda, M. Cooperative Roles of Nitric Oxide-Metabolizing Enzymes to Counteract Nitrosative Stress in Enterohemorrhagic Escherichia coli. Infect. Immun. 2019, 87, e00334-19. [Google Scholar] [CrossRef] [Green Version]
- Pullan, S.T.; Gidley, M.D.; Jones, R.A.; Barrett, J.; Stevanin, T.M.; Read, R.C.; Green, J.; Poole, R.K. Nitric Oxide in Chemostat-Cultured Escherichia coli Is Sensed by Fnr and Other Global Regulators: Unaltered Methionine Biosynthesis Indicates Lack of S Nitrosation. J. Bacteriol. 2007, 189, 1845–1855. [Google Scholar] [CrossRef] [Green Version]
- Bang, I.S.; Liu, L.; Vazquez-Torres, A.; Crouch, M.L.; Stamler, J.S.; Fang, F.C. Maintenance of Nitric Oxide and Redox Homeostasis by the Salmonella Flavohemoglobin Hmp. J. Biol. Chem. 2006, 281, 28039–28047. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Hao, Z.; Wu, J.; Ma, C.; Xu, Y.; Li, J.; Lan, R.; Zhu, B.; Ren, P.; Fan, D.; et al. Comparative Proteomic Analysis of Polarized Human THP-1 and Mouse RAW264.7 Macrophages. Front. Immunol. 2021, 12, 700009. [Google Scholar] [CrossRef] [PubMed]
- Abu-Ali, G.S.; Ouellette, L.M.; Henderson, S.T.; Whittam, T.S.; Manning, S.D. Differences in Adherence and Virulence Gene Expression between Two Outbreak Strains of Enterohaemorrhagic Escherichia coli O157:H7. Microbiology 2010, 156, 408–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neupane, M.; Abu-Ali, G.S.; Mitra, A.; Lacher, D.W.; Manning, S.D.; Riordan, J.T. Shiga Toxin 2 Overexpression in Escherichia coli O157:H7 Strains Associated with Severe Human Disease. Microb. Pathog. 2011, 51, 466–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) | Target | Amplicon Size |
|---|---|---|---|
| norv698_F | ATAACCCGACGCAAATTGT | norV | 675 bp (intact) 470 bp (deletion) |
| norv1373_R | TATCCGGGACTTCACTCCA | ||
| norV-del1 | GCAATTAGCAAGACATCTTTTTAGAACACGCTGAATAAATTGAGGTTGCTGTGTAGGCTGGAGCTGCTTC | Kanamycin marker flanked by 50 bp of norV | 1577 bp |
| norV-del2 | CACCAGTTGGCGGGCGGCGAAGCCCGAACCAATGATCACAATGCCGTTGCCATATGAATATCCTCCTTAG | ||
| norV-del3 | GATATTCGCCAGCACATCAA | ΔnorV | 1868 bp (TW14359) 1905 bp (ΔnorV) |
| norV-del4 | AAACTGCTCGGCAAATTCAC | ||
| norvcompl_F | AGATATTGTCATATCGACCATTGGA | norV + flanking region and norV promoter | 1620 bp |
| norvcompl_R | AATGATCACAATGCCGTTGC |
| Clade | norV † | stx1 | stx1, stx2 | stx1, stx2, stx2c | stx1, stx2c | stx2 | stx2, stx2c | stx2c | Total * |
|---|---|---|---|---|---|---|---|---|---|
| 2 | Deletion | 2 (1.2) | 152 (92.7) | − | − | 10 (6.1) | − | − | 164 |
| 3 | Deletion | 1 (3.0) | 32 (97.0) | − | − | − | − | − | 33 |
| 6 | Intact | − | − | − | − | 1 (12.5) | 1 (12.5) | 6 (75.0) | 8 |
| 7 | Intact | 1 (3.5) | − | 1 (3.5) | 7 (24.1) | − | 6 (20.7) | 14 (48.3) | 29 |
| 8 | Intact | − | 1 (1.5) | 1 (1.5) | − | 30 (45.5) | 33 (50.0) | 1 (1.5) | 66 |
| 9 | Intact | − | 1 (33.3) | − | 2 (66.7) | − | − | − | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al Safadi, R.; Korir, M.L.; Manning, S.D. Nitric Oxide Induced stx2 Expression Is Inhibited by the Nitric Oxide Reductase, NorV, in a Clade 8 Escherichia coli O157:H7 Outbreak Strain. Microorganisms 2022, 10, 106. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10010106
Al Safadi R, Korir ML, Manning SD. Nitric Oxide Induced stx2 Expression Is Inhibited by the Nitric Oxide Reductase, NorV, in a Clade 8 Escherichia coli O157:H7 Outbreak Strain. Microorganisms. 2022; 10(1):106. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10010106
Chicago/Turabian StyleAl Safadi, Rim, Michelle L. Korir, and Shannon D. Manning. 2022. "Nitric Oxide Induced stx2 Expression Is Inhibited by the Nitric Oxide Reductase, NorV, in a Clade 8 Escherichia coli O157:H7 Outbreak Strain" Microorganisms 10, no. 1: 106. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10010106