
Differential Amino Acid Uptake and Depletion in Mono-Cultures and Co-Cultures of Streptococcus thermophilus and Lactobacillus delbrueckii subsp. bulgaricus in a Novel Semi-Synthetic Medium
 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Cultivation Conditions
2.2. Acidification Measurements
2.3. Medium Preparation
2.3.1. Complex Media
2.3.2. Semi-Synthetic Medium
2.4. Cell Dry Weight (DW)
2.5. Biomass Measurements Using Flow Cytometry
2.6. Quantification of Fermentation Products
2.7. Total Amino Acid Composition in the Supernatant
2.8. Calculation of Amino Acid Production Rates
2.9. Fitting of Gaussian Models to pH-Dependent Amino Acid Production Rate
2.10. Simulation of Amino Acid Concentrations
2.11. Uncertainty Analysis
3. Results
3.1. Medium Development
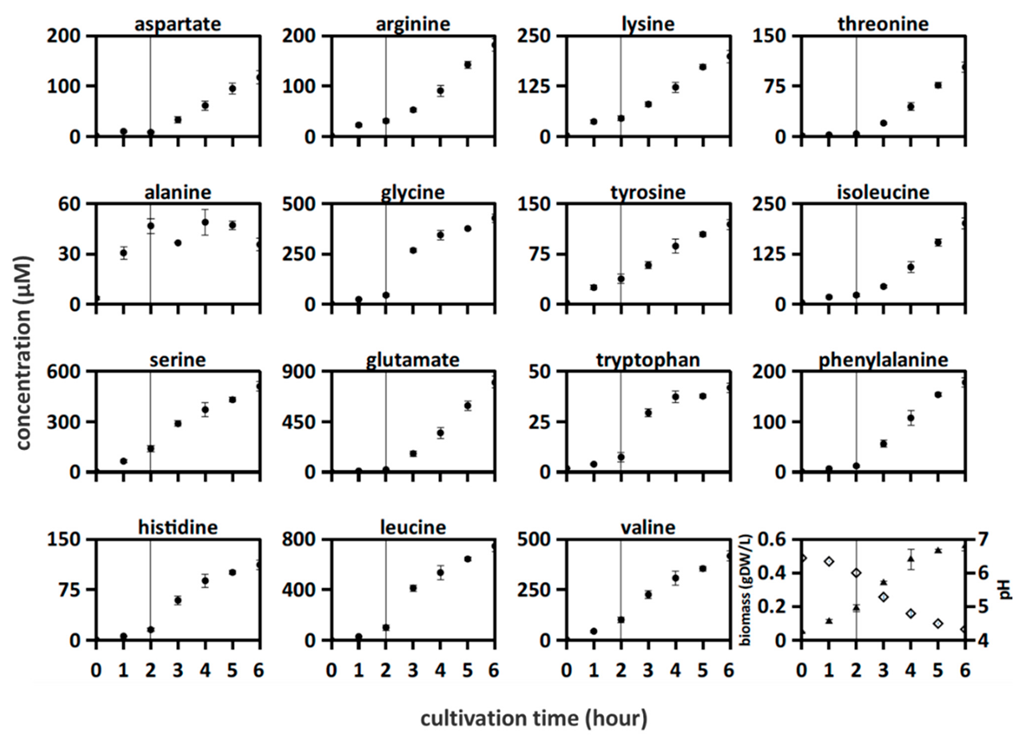
3.2. Growth and Amino Acid Release in L. bulgaricus Mono-Culture
3.3. Growth and Amino Acid Release in Proteinase-Positive S. thermophilus Mono-Culture
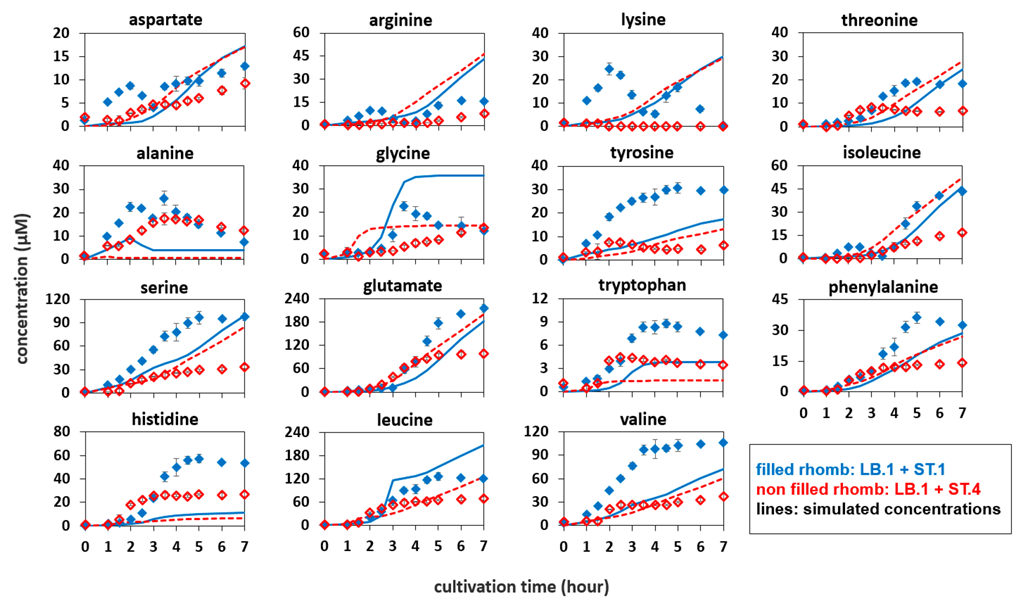
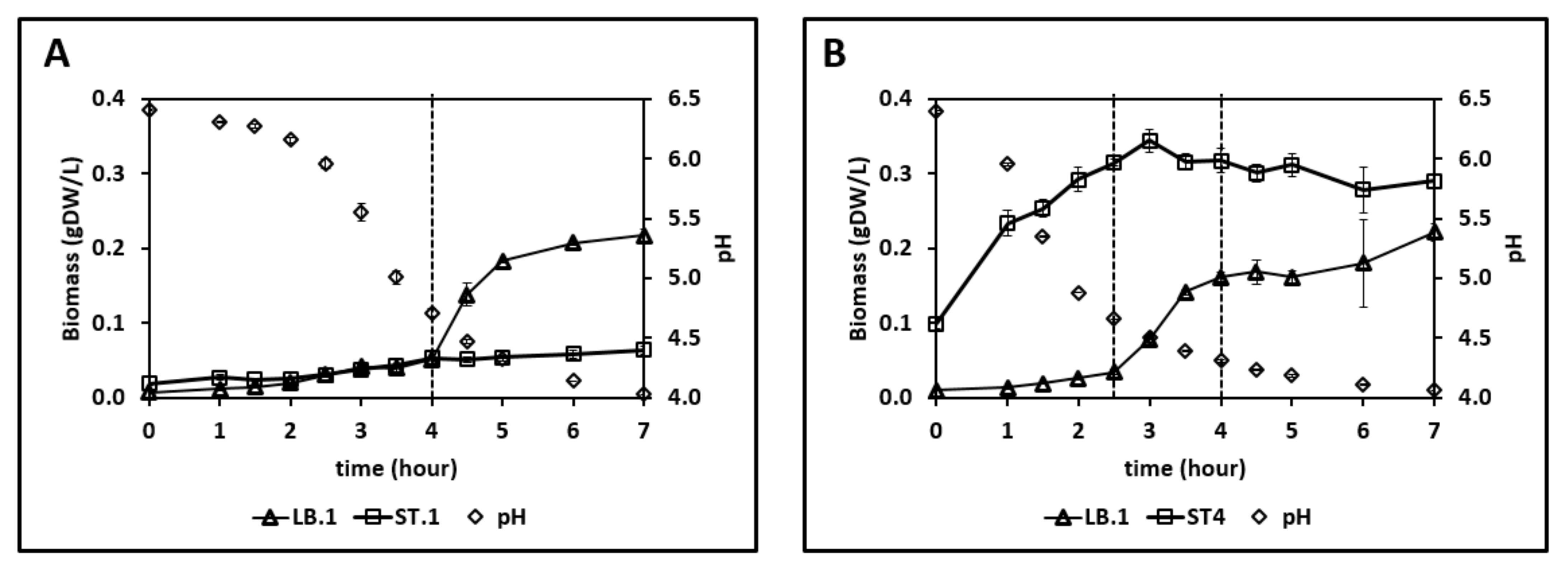
3.4. Growth and Amino Acid Release in the Co-Culture of Proteinase-Positive S. thermophilus and L. bulgaricus
3.5. Growth and Amino Acid Release in the Co-Culture of Proteinase-Negative S. thermophilus and L. bulgaricus
3.6. Simulation of Amino Acid Concentrations to Compare Mono- and Co-Culture Cultivations
4. Discussion
4.1. Amino Acids Are Consumed by L. bulgaricus and S. thermophilus
4.2. Amino Acids Can Accumulate in Cultivations with L. bulgaricus and S. thermophilus
4.3. Differences between Co-Cultures with Different S. thermophilus Strains
4.4. Co-Culture Is Not the Sum of Mono-Cultures
4.5. Stimulatory Effects of Branched-Chain Amino Acid (BCAA) Depletion
4.6. Arginine and Lysine Depletion in Co-Cultures
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marco, M.L.; Heeney, D.; Binda, S.; Cifelli, C.J.; Cotter, P.D.; Foligné, B.; Gänzle, M.; Kort, R.; Pasin, G.; Pihlanto, A.; et al. Health Benefits of Fermented Foods: Microbiota and Beyond. Curr. Opin. Biotechnol. 2017, 44, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Markakiou, S.; Gaspar, P.; Johansen, E.; Zeidan, A.A.; Neves, A.R. Harnessing the Metabolic Potential of Streptococcus thermophilus for New Biotechnological Applications. Curr. Opin. Biotechnol. 2020, 61, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Arioli, S.; della Scala, G.; Remagni, M.C.; Stuknyte, M.; Colombo, S.; Guglielmetti, S.; de Noni, I.; Ragg, E.; Mora, D. Streptococcus thermophilus Urease Activity Boosts Lactobacillus delbrueckii subsp. Bulgaricus Homolactic Fermentation. Int. J. Food Microbiol. 2017, 247, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Blasche, S.; Kim, Y.; Mars, R.A.T.; Machado, D.; Maansson, M.; Kafkia, E.; Milanese, A.; Zeller, G.; Teusink, B.; Nielsen, J.; et al. Metabolic Cooperation and Spatiotemporal Niche Partitioning in a Kefir Microbial Community. Nat. Microbiol. 2021, 6, 196–208. [Google Scholar] [CrossRef] [PubMed]
- Schöpping, M.; Gaspar, P.; Neves, A.R.; Fanzén, C.J.; Zeidan, A.A. Identifying the Essential Nutritional Requirements of the the Probiotic Bacteria Bifidobacterium animalis and Bifidobacterium longum Using Genome-Scale Modeling. NPJ Syst. Biol. Appl. 2021, 7, 1–15. [Google Scholar] [CrossRef]
- Settachaimongkon, S.; Nout, M.J.R.; Antunes Fernandes, E.C.; Hettinga, K.A.; Vervoort, J.M.; van Hooijdonk, T.C.M.; Zwietering, M.H.; Smid, E.J.; van Valenberg, H.J.F. Influence of Different Proteolytic Strains of Streptococcus thermophilus in Co-Culture with Lactobacillus delbrueckii subsp. Bulgaricus on the Metabolite Profile of Set-Yoghurt. Int. J. Food Microbiol. 2014, 177, 29–36. [Google Scholar] [CrossRef]
- Johansen, E. Use of Natural Selection and Evolution to Develop New Starter Cultures for Fermented Foods. Annu. Rev. Food Sci. Technol. 2018, 9, 411–428. [Google Scholar] [CrossRef]
- Chandan, R.C.; Gandhi, A.; Shah, N.P. Yogurt: Historical Background, Health Benefits, and Global Trade; Elsevier Inc.: Amsterdam, The Netherlands, 2017. [Google Scholar] [CrossRef]
- Liu, E.; Zheng, H.; Hao, P.; Konno, T.; Kume, H.; Oda, M.; Suzuki, K.; Ji, Z. Acquisition of Amino Acids by Lactobacillus delbrueckii subsp. Bulgaricus 2038 When Grown in the Presence of Casein. Int. Dairy J. 2014, 35, 145–152. [Google Scholar] [CrossRef]
- de Souza Oliveira, R.P.; Torres, B.R.; Perego, P.; de Oliveira, M.N.; Converti, A. Co-Metabolic Models of Streptococcus thermophilus in Co-Culture with Lactobacillus bulgaricus or Lactobacillus acidophilus. Biochem. Eng. J. 2012, 62, 62–69. [Google Scholar] [CrossRef]
- Herve-Jimenez, L.; Guillouard, I.; Guedon, E.; Gautier, C.; Boudebbouze, S.; Hols, P.; Monnet, V.; Rul, F.; Maguin, E. Physiology of Streptococcus thermophilus during the Late Stage of Milk Fermentation with Special Regard to Sulfur Amino-Acid Metabolism. Proteomics 2008, 8, 4273–4286. [Google Scholar] [CrossRef]
- Herve-Jimenez, L.; Guillouard, I.; Guedon, E.; Boudebbouze, S.; Hols, P.; Monnet, V.; Maguin, E.; Rul, F. Postgenomic Analysis of Streptococcus thermophilus Cocultivated in Milk with Lactobacillus delbrueckii subsp. Bulgaricus: Involvement of Nitrogen, Purine, and Iron Metabolism. Appl. Environ. Microbiol. 2009, 75, 2062–2073. [Google Scholar] [CrossRef]
- Sieuwerts, S.; Molenaar, D.; van Hijum, S.A.F.T.; Beerthuyzen, M.; Stevens, M.J.A.; Janssen, P.W.M.; Ingham, C.J.; de Bok, F.A.M.; de Vos, W.M.; van Hylckama Vlieg, J.E.T. Mixed-Culture Transcriptome Analysis Reveals the Molecular Basis of Mixed-Culture Growth in Streptococcus thermophilus and Lactobacillus bulgaricus. Appl. Environ. Microbiol. 2010, 76, 7775–7784. [Google Scholar] [CrossRef] [PubMed]
- Mendes, F.; Sieuwerts, S.; de Hulster, E.; Almering, M.J.H.; Luttik, M.A.H.; Pronk, J.T.; Smid, E.J.; Bron, P.A.; Daran-Lapujadea, P. Transcriptome-Based Characterization of Interactions between Saccharomyces cerevisiae and Lactobacillus delbrueckii subsp. Bulgaricus in Lactose-Grown Chemostat Cocultures. Appl. Environ. Microbiol. 2013, 79, 5949–5961. [Google Scholar] [CrossRef] [PubMed]
- Branco dos Santos, F.; de Vos, W.M.; Teusink, B. Towards Metagenome-Scale Models for Industrial Applications-the Case of Lactic acid bacteria. Curr. Opin. Biotechnol. 2013, 24, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Somerville, V.; Grigaitis, P.; Battjes, J.; Moro, F.; Teusink, B. Use and Limitations of Genome-Scale Metabolic Models in Food Microbiology. Curr. Opin. Food Sci. 2022, 43, 225–231. [Google Scholar] [CrossRef]
- Chen, S.; Niu, H.; Wu, Y.; Sun, J.; Han, X.; Zhang, L. Influence of Lactic Acid on Cell Cycle Progressions in Lactobacillus bulgaricus During Batch Culture. Appl. Biochem. Biotechnol. 2021, 193, 912–924. [Google Scholar] [CrossRef]
- Russell, J.B.; Diez-Gonzalez, F. The Effects of Fermentation Acids on Bacterial Growth. Adv. Microb. Physiol. 1998, 39, 228–234. [Google Scholar] [CrossRef]
- Douwenga, S.; Janssen, P.; Teusink, B. A Centrifugation-Based Clearing Method Allows High-Throughput Acidification and Growth-Rate Measurements in Milk. J. Dairy Sci. 2021, 8, 8530–8540. [Google Scholar] [CrossRef]
- Radke-Mitchell, L.; Sandine, W.E. Associative Growth and Differential Enumeration of Streptococcus thermophilus and Lactobacillus bulgaricus: A Review. J. Food Prot. 1984, 47, 245–248. [Google Scholar] [CrossRef]
- Grobben, G.J.; Sikkema, J.; Smith, M.R.; de Bont, J.A.M. Production of Extracellular Polysaccharides by Lactobacillus delbrueckii ssp. Bulgaricus NCFB 2772 Grown in a Chemically Defined Medium. J. Appl. Bacteriol. 1995, 79, 103–107. [Google Scholar] [CrossRef]
- Liu, E.; Zheng, H.; Shi, T.; Ye, L.; Konno, T.; Oda, M.; Shen, H.; Ji, Z.S. Relationship between Lactobacillus bulgaricus and Streptococcus thermophilus under Whey Conditions: Focus on Amino Acid Formation. Int. Dairy J. 2016, 56, 141–150. [Google Scholar] [CrossRef]
- Radke-Mitchell, L.C.; Sandine, W.E. Influence of Temperature on Associative Growth of Streptococcus thermophilus and Lactobacillus bulgaricus. J. Dairy Sci. 1986, 69, 2558–2568. [Google Scholar] [CrossRef]
- Buchholz, J.; Schwentner, A.; Brunnenkan, B.; Gabris, C.; Grimm, S.; Gerstmeir, R.; Takors, R.; Eikmanns, B.J.; Blombacha, B. Platform Engineering of Corynebacterium glutamicum with Reduced Pyruvate Dehydrogenase Complex Activity for Improved Production of L-Lysine, l-Valine, and 2-Ketoisovalerate. Appl. Environ. Microbiol. 2013, 79, 5566–5575. [Google Scholar] [CrossRef]
- Kunji, E.R.S. The Proteolytic Systems of Lactic acid bacteria. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 1996, 70, 187–221. [Google Scholar] [CrossRef]
- Rodríguez-Serrano, G.M.; García-Garibay, J.M.; Cruz-Guerrero, A.E.; Gómez-Ruiz, L.D.C.; Ayala-Niño, A.; Castañeda-Ovando, A.; González-Olivares, L.G. Proteolytic System of Streptococcus thermophilus. J. Microbiol Biotechnol. 2018, 28, 1581–1588. [Google Scholar] [CrossRef]
- Hetényi, K.; Németh, Á.; Sevella, B. Role of PH-Regulation in Lactic Acid Fermentation: Second Steps in a Process Improvement. Chem. Eng. Processing: Process Intensif. 2011, 50, 293–299. [Google Scholar] [CrossRef]
- Crittenden, R.G.; Martinez, N.R.; Playne, M.J. Synthesis and Utilisation of Folate by Yoghurt Starter Cultures and Probiotic Bacteria. Int. J. Food Microbiol. 2003, 80, 217–222. [Google Scholar] [CrossRef]
- Sieuwerts, S.; de Bok, F.A.M.; Hugenholtz, J.; van Hylckama Vlieg, J.E.T. Unraveling Microbial Interactions in Food Fermentations: From Classical to Genomics Approaches. Appl. Environ. Microbiol. 2008, 74, 4997–5007. [Google Scholar] [CrossRef]
- Otto, R.; Tenbrink, B.; Veldkamp, H.; Konings, W.N. The Relation between Growth-Rate and Electrochemical Proton Gradient of Streptococcus-Cremoris. FEMS Microbiol. Lett. 1983, 16, 69–74. [Google Scholar] [CrossRef]
- Letort, C.; Juillard, V. Development of a Minimal Chemically-Defined Medium for the Exponential Growth of Streptococcus thermophilus. J. Appl. Microbiol. 2001, 91, 1023–1029. [Google Scholar] [CrossRef]
- Chervaux, C.; Ehrlich, S.D.; Maguin, E. Physiological Study of Lactobacillus delbrueckii subsp. Bulgaricus Strains in a Novel Chemically Defined Medium. Society 2000, 66, 5306–5311. [Google Scholar] [CrossRef]
- Courtin, P.; Monnet, V.; Rul, F. Cell-Wall Proteinases PrtS and PrtB Have a Different Role in Streptococcus thermophilus/Lactobacillus bulgaricus Mixed Cultures in Milk. Microbiology (N Y) 2002, 148, 3413–3421. [Google Scholar] [CrossRef]
- Liu, E.; Hao, P.; Konno, T.; Yu, Y.; Oda, M.; Zheng, H.; Ji, Z. Amino Acid Biosynthesis and Proteolysis in Lactobacillus bulgaricus Revisited: A Genomic Comparison. Computat. Mol. Biosci. 2012, 2012, 61–77. [Google Scholar] [CrossRef]
- van de Guchte, M.; Penaud, S.; Grimaldi, C.; Barbe, V.; Bryson, K.; Nicolas, P.; Robert, C.; Oztas, S.; Mangenot, S.; Couloux, A.; et al. The Complete Genome Sequence of Lactobacillus bulgaricus Reveals Extensive and Ongoing Reductive Evolution. Proc. Natl. Acad. Sci. 2006, 103, 9274–9279. [Google Scholar] [CrossRef]
- Hols, P.; Hancy, F.; Fontaine, L.; Grossiord, B.; Prozzi, D.; Leblond-Bourget, N.; Decaris, B.; Bolotin, A.; Delorme, C.; Ehrlich, S.D.; et al. New Insights in the Molecular Biology and Physiology of Streptococcus thermophilus Revealed by Comparative Genomics. FEMS Microbiol. Rev. 2005, 29, 435–463. [Google Scholar] [CrossRef]
- Zheng, H.; Liu, E.; Hao, P. In Silico Analysis of Amino Acid Biosynthesis and Proteolysis in Lactobacillus delbrueckii subsp. Bulgaricus 2038 and the Implications for Bovine Milk Fermentation. Biotechnol. Lett. 2012, 34, 1545–1551. [Google Scholar] [CrossRef] [PubMed]
- Smid, E.J.; Lacroix, C. Microbe-Microbe Interactions in Mixed Culture Food Fermentations. Curr. Opin. Biotechnol. 2013, 24, 148–154. [Google Scholar] [CrossRef]
- Rau, M.H.; Gaspar, P.; Jensen, M.L.; Geppel, A.; Neves, A.R.; Zeidan, A.A. Genome-Scale Metabolic Modeling Combined with Transcriptome Profiling Provides Mechanistic Understanding of Streptococcus thermophilus CH8 Metabolism. Appl. Environ. Microbiol. 2022, 88, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Li, Y. Soybean Peptides Promote Yoghurt Fermentation and Quality. Biotechnol. Lett. 2020, 42, 1927–1937. [Google Scholar] [CrossRef]
- Kliche, T.; Li, B.; Bockelmann, W.; Habermann, D.; Klempt, M.; de Vrese, M. Screening for Proteolytically Active Lactic acid bacteria and Bioactivity of Peptide Hydrolysates Obtained with Selected Strains. Appl. Microbiol. Biotechnol. 2017, 101, 7621–7633. [Google Scholar] [CrossRef] [PubMed]
- Liu, E.; Zheng, H.; Hao, P.; Konno, T.; Yu, Y.; Kume, H.; Oda, M.; Ji, Z.S. A Model of Proteolysis and Amino Acid Biosynthesis for Lactobacillus delbrueckii subsp. Bulgaricus in Whey. Curr. Microbiol. 2012, 65, 742–751. [Google Scholar] [CrossRef] [PubMed]
- Hutkins, R.W.; Nannen, N.L. pH Homeostasis in Lactic acid bacteria. J. Dairy Sci. 1993, 76, 2354–2365. [Google Scholar] [CrossRef]
- Rajagopal, S.N.; Sandine, W.E. Associative Growth and Proteolysis of Streptococcus thermophilus and Lactobacillus bulgaricus in Skim Milk. J. Dairy Sci. 1990, 73, 894–899. [Google Scholar] [CrossRef]
- Beshkova, D.M.; Simova, E.D.; Frengova, G.I.; Simov, Z.I.; Adilov, E.F. Production of Amino Acids by Yogurt Bacteria. Biotechnol. Prog. 1998, 14, 963–965. [Google Scholar] [CrossRef]
- Pastink, M.I.; Teusink, B.; Hols, P.; Visser, S.; de Vos, W.M.; Hugenholtz, J. Genome-Scale Model of Streptococcus thermophilus LMG18311 for Metabolic Comparison of Lactic acid bacteria. Appl. Environ. Microbiol. 2009, 75, 3627–3633. [Google Scholar] [CrossRef] [PubMed]
- Wintermute, E.H.; Silver, P.A. Dynamics in the Mixed Microbial Concourse. Genes Dev. 2010, 24, 2603–2614. [Google Scholar] [CrossRef]
- Hao, P.; Zheng, H.; Yu, Y.; Ding, G.; Gu, W.; Chen, S.; Yu, Z.; Ren, S.; Oda, M.; Konno, T.; et al. Complete Sequencing and Pan-Genomic Analysis of Lactobacillus delbrueckii subsp. Bulgaricus Reveal Its Genetic Basis for Industrial Yogurt Production. PLoS ONE 2011, 6, e15964. [Google Scholar] [CrossRef]
- Garault, P.; Letort, C.; Juillard, V.; Monnet, V. Branched-Chain Amino Acids and Purine Biosynthesis: Two Pathways Essential for Optimal Growth of Streptococcus thermophilus in Milk. Lait 2001, 81, 83–90. [Google Scholar] [CrossRef]
- Arioli, S.; Roncada, P.; Salzano, A.M.; Deriu, F.; Corona, S.; Guglielmetti, S.; Bonizzi, L.; Scaloni, A.; Mora, D. The Relevance of Carbon Dioxide Metabolism in Streptococcus thermophilus. Microbiology 2009, 155, 1953–1965. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Compound | Concentration [g L−1] | CAS Number |
|---|---|---|---|
| - | Di-potassium hydrogen phosphate | 2.5 | 7758-11-4 |
| Potassium dihydrogen phosphate | 3 | 7778-77-0 | |
| Sodium acetate | 1 | 127-09-3 | |
| Ammonium citrate tribasic | 0.6 | 3458-72-8 | |
| Manganese sulfate monohydrate | 0.02 | 10034-96-5 | |
| Iron(II) sulfate heptahydrate | 0.00132 | 7782-63-0 | |
| Calcium chloride dihydrate | 0.08745 | 10035-04-8 | |
| Tween 80 | 1 mL L−1 | 9005-65-6 | |
| D-Lactose monohydrate | 15.75 | 10039-26-6 | |
| Magnesium sulfate heptahydrate | 0.2 | 10034-99-8 | |
| Urea | 0.12 | 57-13-6 | |
| nucleobases | Adenine | 0.01 | 73-24-5 |
| Guanine | 0.01 | 73-40-5 | |
| Uracil | 0.01 | 66-22-8 | |
| Xanthine | 0.01 | 69-89-6 | |
| vitamins | Biotin | 0.0002 | 58-85-5 |
| Folic acid | 0.0002 | 59-30-3 | |
| Pyridoxal hydrochloride | 0.001 | 65-22-5 | |
| Riboflavin | 0.0005 | 83-88-5 | |
| Thiamine chloride hydrochloride | 0.0005 | 67-03-8 | |
| Nicotinamide | 0.0005 | 98-92-0 | |
| Cyanocobalamin | 0.0005 | 68-19-9 | |
| 4-Aminobenzoic acid | 0.0005 | 150-13-0 | |
| D-Pantothenic acid hemicalcium salt | 0.004 | 137-08-6 | |
| DL-6,8-thioctic acid | 0.0005 | 1077-28-7 | |
| trace elements | Ammonium molybdate tetrahydrate | 0.0000037 | 12054-85-2 |
| Cobalt(II) chloride hexahydrate | 0.000007 | 7791-13-1 | |
| Boric acid | 0.000025 | 10043-35-3 | |
| Copper(II) sulfate pentahydrate | 0.0000025 | 7758-99-8 | |
| Zinc sulfate heptahydrate | 0.0000029 | 7446-20-0 | |
| amino acids | L-Alanine | 0.1 | 56-41-7 |
| L-Arginine | 0.317 | 74-79-3 | |
| L-Asparagine monohydrate | 0.343 | 5794-13-8 | |
| L-Aspartic acid | 0.499 | 56-84-8 | |
| L-Cysteine hydrochloride monohydrate | 0.3 | 7048-04-6 | |
| L-Glutamic acid | 0.331 | 56-86-0 | |
| L-Glutamine | 0.29 | 56-85-9 | |
| Glycine | 0.16 | 56-40-6 | |
| L-Histidine monohydrochloride monohydrate | 0.273 | 5934-29-2 | |
| L-Isoleucine | 0.361 | 73-32-5 | |
| L-Leucine | 0.6 | 61-90-5 | |
| L-Lysine | 0.351 | 56-87-1 | |
| L-Methionine | 0.119 | 63-68-3 | |
| L-Phenylalanine | 0.34 | 63-91-2 | |
| L-Proline | 0.921 | 147-85-3 | |
| L-Serine | 0.359 | 56-45-1 | |
| L-Threonine | 0.3 | 72-19-5 | |
| L-Tryptophan | 0.102 | 73-22-3 | |
| L-Tyrosine | 0.12 | 60-18-4 | |
| L-Valine | 0.468 | 72-18-4 | |
| casein | Casein | 2 | 9005-46-3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ulmer, A.; Erdemann, F.; Mueller, S.; Loesch, M.; Wildt, S.; Jensen, M.L.; Gaspar, P.; Zeidan, A.A.; Takors, R. Differential Amino Acid Uptake and Depletion in Mono-Cultures and Co-Cultures of Streptococcus thermophilus and Lactobacillus delbrueckii subsp. bulgaricus in a Novel Semi-Synthetic Medium. Microorganisms 2022, 10, 1771. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10091771
Ulmer A, Erdemann F, Mueller S, Loesch M, Wildt S, Jensen ML, Gaspar P, Zeidan AA, Takors R. Differential Amino Acid Uptake and Depletion in Mono-Cultures and Co-Cultures of Streptococcus thermophilus and Lactobacillus delbrueckii subsp. bulgaricus in a Novel Semi-Synthetic Medium. Microorganisms. 2022; 10(9):1771. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10091771
Chicago/Turabian StyleUlmer, Andreas, Florian Erdemann, Susanne Mueller, Maren Loesch, Sandy Wildt, Maiken Lund Jensen, Paula Gaspar, Ahmad A. Zeidan, and Ralf Takors. 2022. "Differential Amino Acid Uptake and Depletion in Mono-Cultures and Co-Cultures of Streptococcus thermophilus and Lactobacillus delbrueckii subsp. bulgaricus in a Novel Semi-Synthetic Medium" Microorganisms 10, no. 9: 1771. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10091771