
Effect of Carbon Sources in Carotenoid Production from Haloarcula sp. M1, Halolamina sp. M3 and Halorubrum sp. M5, Halophilic Archaea Isolated from Sonora Saltern, Mexico


 , , ,
, , ,  ,
, 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Organisms and Culture Media
2.2. DNA Extraction and Phylogenetic Analysis
2.3. Chemo-Taxonomic Identification
2.4. Carotenoid Production
2.5. Optimization of Carotenoid Production: Effect of Carbon Sources
2.6. Statistical Analysis
3. Results
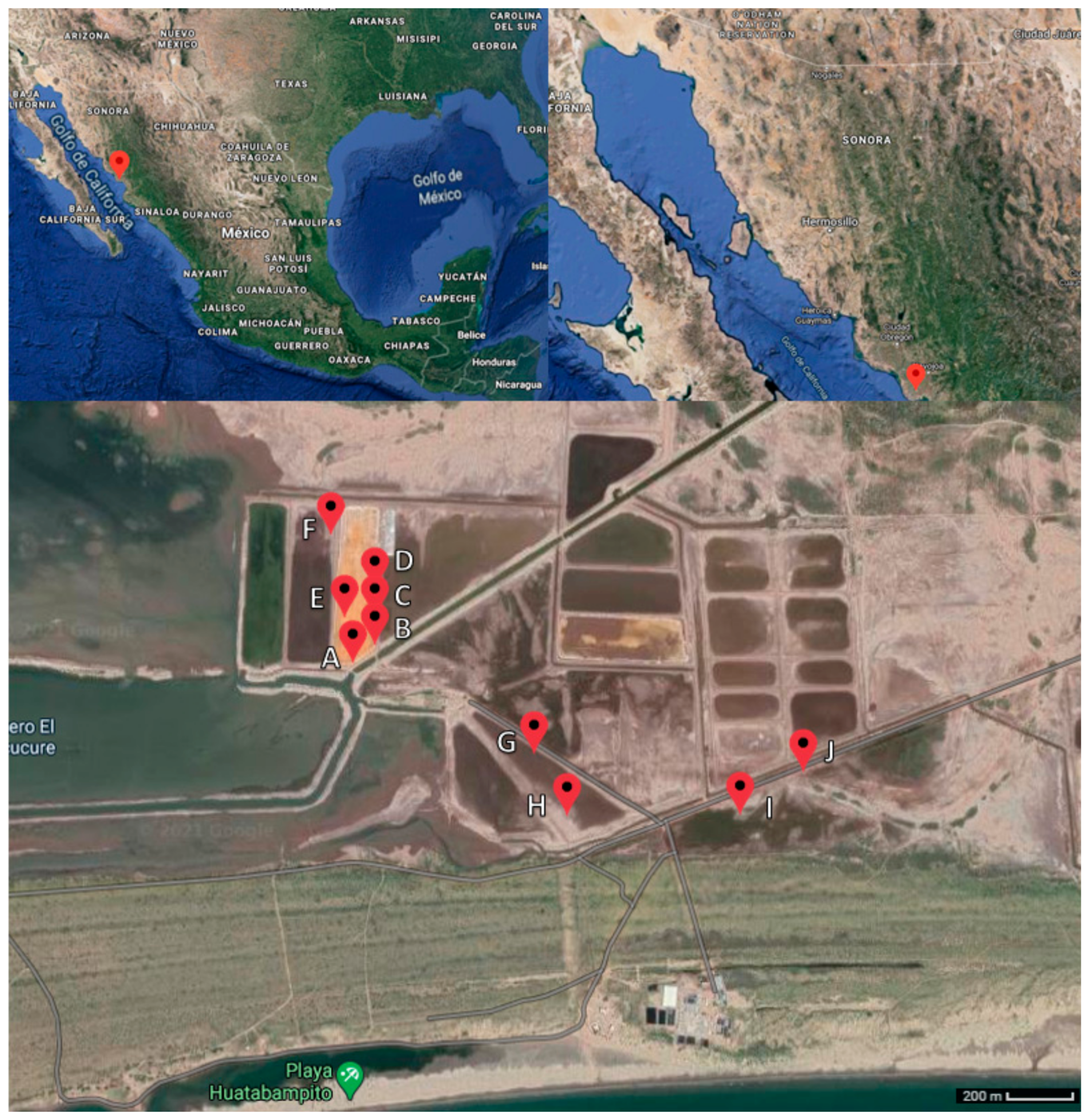
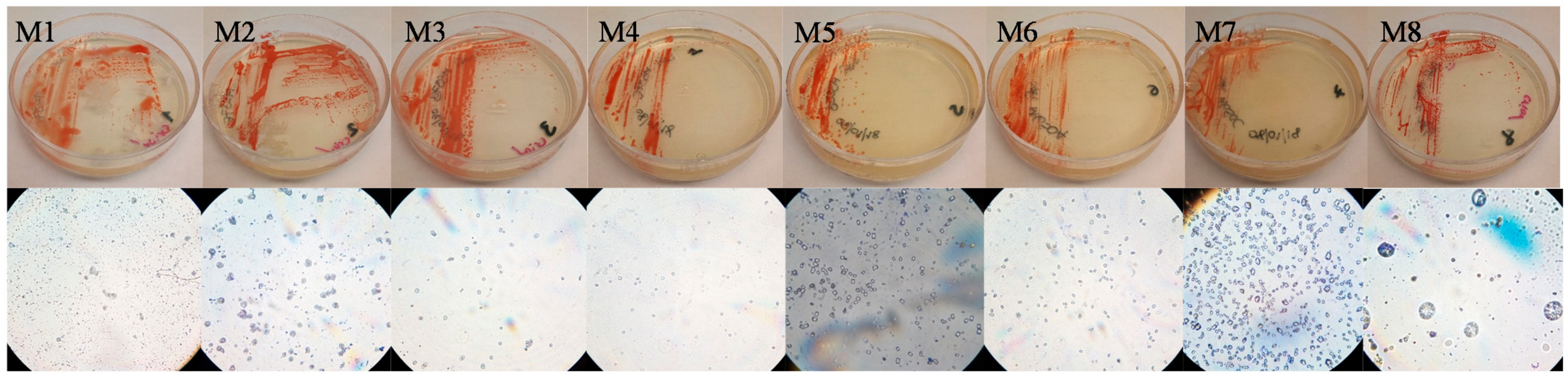
3.1. Sampling and Isolation
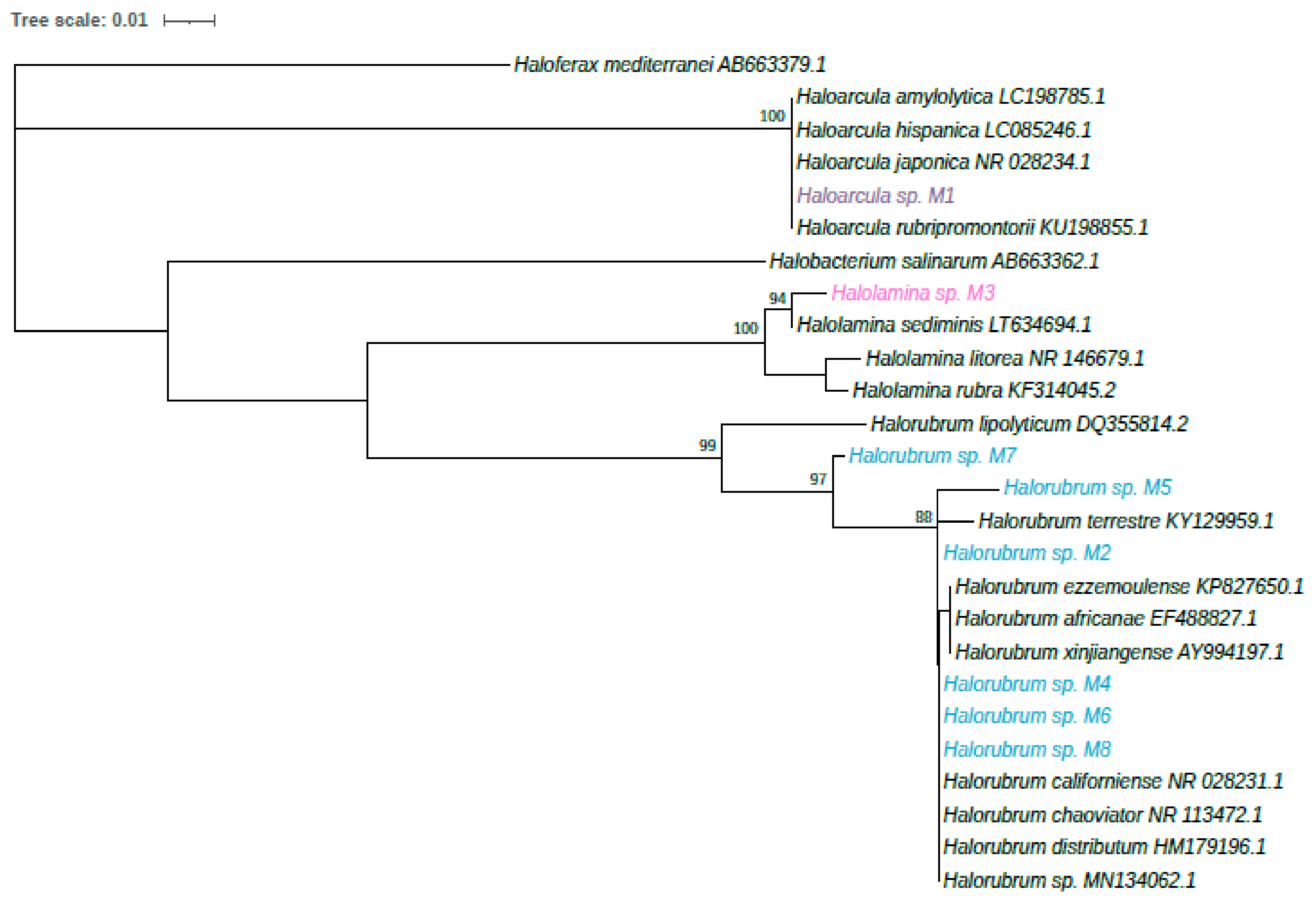
3.2. Identification of Halophilic Archaea Strains
3.3. Carotenoid Producers
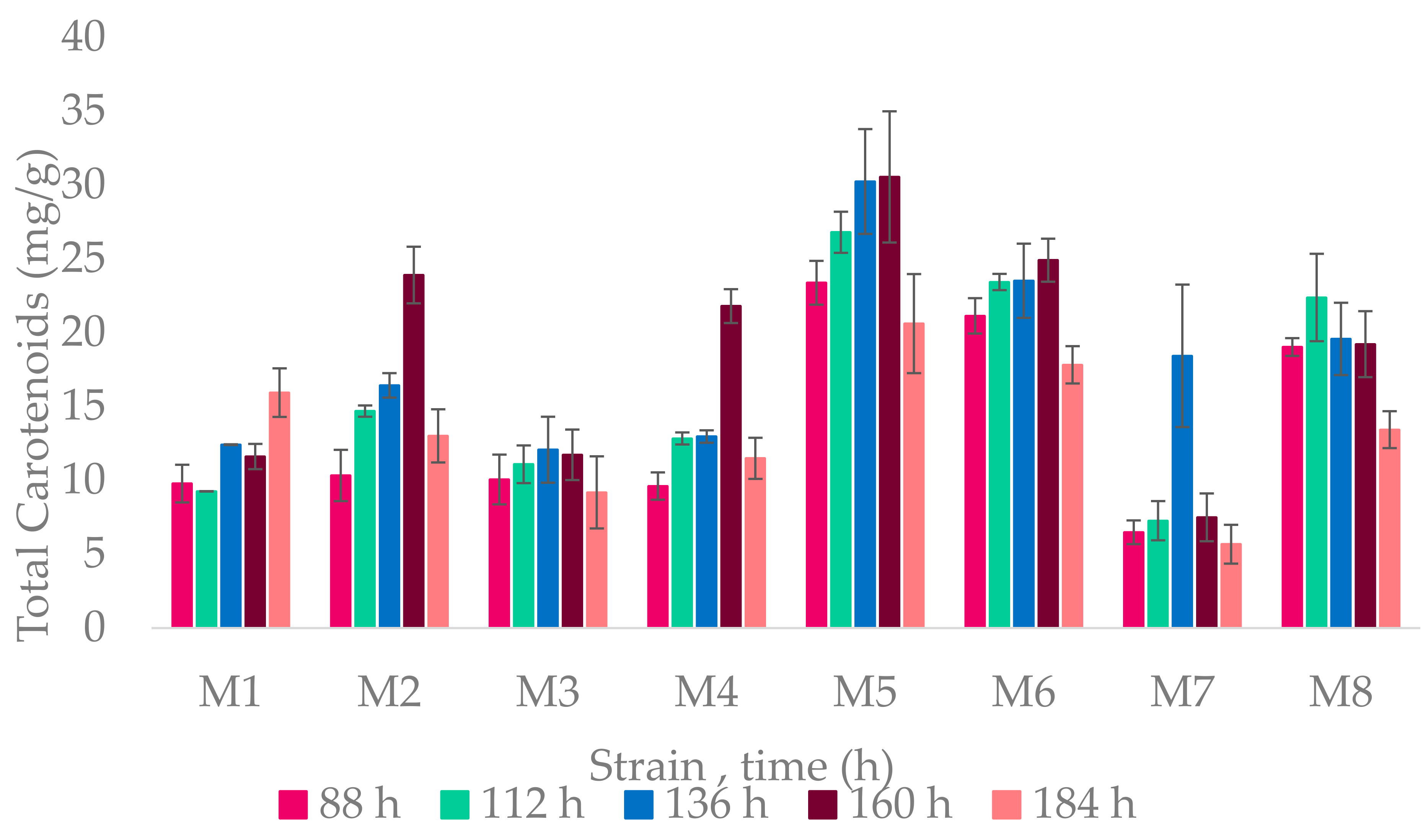
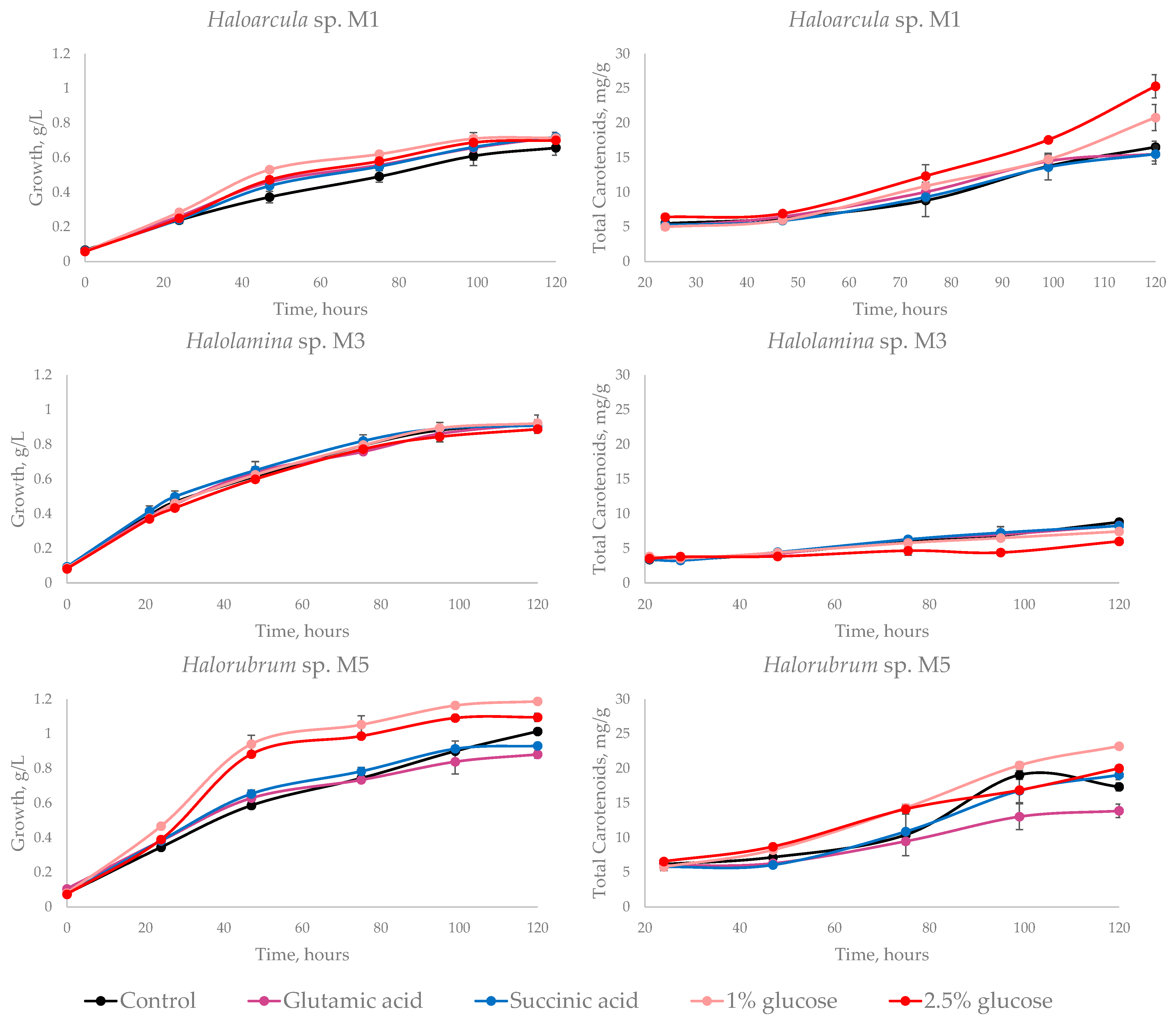
3.3.1. Carbon Source Influence in Carotenoid Production
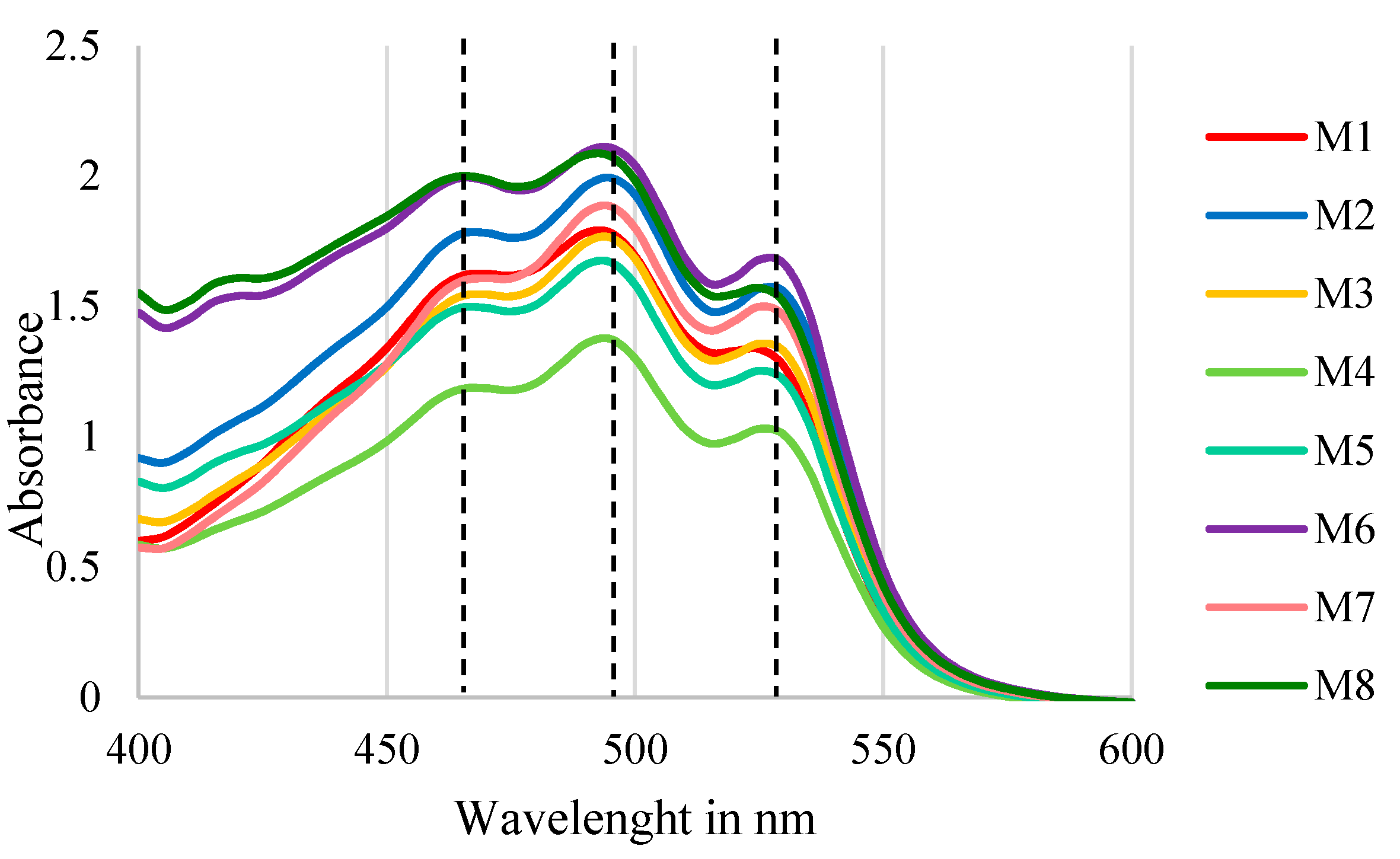
3.3.2. Pigments Identification
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stackebrandt, P.D.F.E.; Sanderson, K.; Volkman, K.; Cameron, D.E. Halobacterium lacusprofundi sp. Nov., a Halophilic Bacterium Isolated from Deep Lake, Antartica. Syst. Appl. Microbiol. 1988, 11, 20–27. [Google Scholar]
- Cayol, J.; Olivier, B.; Patel, B.K.C.; Prensier, G.; Guezennec, J.; Garcia, J. Isolation and Characterization of Halothermothrix orenii gen. sp nov, a Halophilic, Thermophilic, Fermentative Strictly Anaerobic Bacterium. Int. J. Syst. Evol. Microbiol. 1994, 44, 534–540. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Yatsunami, R.; Ando, A.; Miyoko, N.; Fukui, T.; Takaichi, S.; Nakamura, S. Complete biosynthetic pathway of the C50 carotenoid bacterioruberin from lycopene in the extremely halophilic archaeon Haloarcula japonica. J. Bacteriol. 2015, 197, 1614–1623. [Google Scholar] [CrossRef] [Green Version]
- Mandelli, F.; Miranda, V.S.; Rodrigues, E.; Mercadante, A.Z. Identification of carotenoids with high antioxidant capacity produced by extremophile microorganisms. World J. Microbiol. Biotechnol. 2012, 28, 1781–1790. [Google Scholar] [CrossRef] [PubMed]
- Calegari-Santos, R.; Diogo, R.A.; Fontana, J.D.; Bonfim, T.M.B. Carotenoid Production by Halophilic Archaea Under Different Culture Conditions. Curr. Microbiol. 2016, 72, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Souza, S.E.D.; Altekar, W.; D’Souza, S.F. Adaptive response of hfx mdi to low NaCl. Arch. Microbiol. 1997, 168, 68–71. [Google Scholar] [CrossRef]
- Yatsunami, R.; Ando, A.; Yang, Y.; Takaichi, S.; Kohno, M.; Matsumura, Y.; Ikeda, H.; Fukui, T.; Nakasone, K.; Fujita, N.; et al. Identification of carotenoids from extremely halophilic archaeon Haloarcula Japonica. Front. Microbiol. 2014, 5, 100. [Google Scholar] [CrossRef]
- Zalazar, L.; Pagola, P.; Miró, M.V.; Churio, M.S.; Cerletti, M.; Martínez, C.; Iniesta-Cuerda, M.; Soler, A.J.; Cesari, A.; De Castro, R. Bacterioruberin extracts from a genetically modified hyperpigmented Haloferax volcanii strain: Antioxidant activity and bioactive properties on sperm cells. J. Appl. Microbiol. 2019, 126, 796–810. [Google Scholar] [CrossRef] [PubMed]
- McGenity, T.J.; Oren, A. Hypersaline Environments. In Life at Extremes: Environments, Organisms and Strategies for Survival; Bell, M.E., Ed.; Cabi: Cambridge, MA, USA, 2016; pp. 402–437. [Google Scholar]
- Villa-Andrade, M.F. Humedales de Yavaros–Moroncarit. Ficha Informativa de los Humedales de Ramsar (FIR); Comisión de Ecología y Desarrollo Sustentable del Estado de Sonora (CEDES): Hermosillo, Mexico, 2011; Volume 2009–2012. [Google Scholar]
- Coronado, J.C.; De los Santos, S.; Prado, L.A.; Buenrostro, J.J.; Vásquez-Murrieta, M.S.; Estrada, M.I.; Cira, L.A. Isolation of moderately halphilic bacteria in saline environments of Sonora State searching for proteolylic hydrolases. Open Agric. 2018, 3, 207–213. [Google Scholar]
- Vigneron, A.; L’Haridon, S.; Godfroy, A.; Roussel, E.; Cragg, B.; Parkes, R.; Toffin, L. Evidence of active methanogen communities in shallow sediments of the Sonora margin cold seeps. Appl. Environ. Microbiol. 2015, 81, 3451–3459. [Google Scholar] [CrossRef] [Green Version]
- McGenity, T.J.; Grant, W.D. Transfer of Halobacterium saccharovorum, Halobacterium sodomense, Halobacterium trapanicum NRC 34021 and Halobacterium lacusprofundi to the Genus Halorubrum gen. nov., as Halorubrum saccharovorum comb. nov., Halorubrum sodomense comb. nov., Halorubrum. Syst. Appl. Microbiol. 1995, 18, 237–243. [Google Scholar] [CrossRef]
- Sabet, S.; Diallo, L.; Hays, L.; Jung, W.; Dillon, J.G. Characterization of halophiles isolated from solar salterns in Baja California, Mexico. Extremophiles 2009, 13, 643–656. [Google Scholar] [CrossRef]
- Peck, R.; Graham, S.; Gregory, A. Species widely distributed in halophilic archaea exhibit opsin-mediated inhibition of bacterioruberin biosynthesis. J. Bacteriol. 2019, 201, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Giani, M.; Miralles-Robledillo, J.M.; Peiró, G.; Pire, C.; Martínez-Espinoza, R.M. Deciphering Pathaways for carotenogenesis in haloarchaea. Molecules 2020, 25, 1197. [Google Scholar] [CrossRef] [Green Version]
- Ng, W.; Mahairas, G.; Pan, M.; Lasky, S.; Thorsson, V.; Swartzell, S.; Hall, J.; Dahl, T.; Welti, R.; Goo, Y.; et al. Genome sequence of Halobacterium species NRC-1. Proc. Natl. Acad. Sci. USA 2000, 97, 12176–12181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Moya, P.; Niehaus, K.; Alcaíno, J.; Baeza, M.; Cifuentes, V. Proteomic and metabolomic analysis of the carotenogenic yeast Xantophyllomyces dendrorhous using different carbon sources. BMC Genom. 2015, 16, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bräsen, C.; Esser, D.; Rauch, B.; Siebers, B. Carbohydrate Metabolism in Archaea: Current Insights into Unusual Enzymes and Pathaways and Their Regulation. Microbiol. Mol. Biol. Rev. 2014, 78, 89–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gochnauer, M.B.; Kushner, D.J. Growth and nutrition of extremely halophilic bacteria. Can. J. Microbiol 1969, 15, 1157–1165. [Google Scholar] [CrossRef]
- Gochnauer, M.B.; Kushwaha, S.C.; Kates, M.; Kushner, D.J. Nutritional control of pigment and isoprenoid compound formation in extremely halophilic bacteria. Archiv Für Mikrobiol. 1972, 84, 339–349. [Google Scholar] [CrossRef]
- Camacho, R.M.; Mateos-Diaz, J.C.; Diaz-Montaño, D.M.; González-Reynoso, O.; Córdova, J. Carboxyl ester hydrolases production and growth of a halophilic archaeon, Halobacterium sp. NRC-1. Extremophiles 2009, 14, 99–106. [Google Scholar] [CrossRef]
- Arahal, D.R.; Dewhirst, F.E.; Paster, B.J. Phylogenetic Analyses of Some Extremely Halophilic Archaea Isolated from Dead Sea Water, Determined on the Basis of Their 16S rRNA Secuences. Appl. Environ. Microbiol. 1996, 62, 3779–3786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Higgins, G. CLUSTAL V: Multiple alignment of DNA and protein sequences. Methods 634 Mol. Biol. 1994, 25, 307–318. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis 636 Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G+C content bases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar]
- Finore, I.; Gioello, A.; Leone, L.; Orlando, P.; Romano, I.; Nicolaus, B.; Poli, A. Aeribacillus composti sp. nov. a thermophilic bacillus isolated from olive mill pomace compost. Int. J. Syst. Evol. Microbiol. 2017, 67, 4830–4835. [Google Scholar] [CrossRef]
- Finore, I.; Orlando, P.; Di Donato, P.; Leone, L.; Nicolaus, B.; Poli, A. Nesterenkonia aurantiaca sp. Nov., an alkaliphilic actinobacterium isolated from Antarctica. Int. J. Syst. Evol. Microbiol. 2016, 66, 1554–1560. [Google Scholar] [CrossRef]
- Naziri, D.; Hamidi, M.; Hassanzadeh, S.; Tarhriz, V.; Zanjani, B.M.; Nazemyieh, H.; Hejazi, M.A.; Hejazi, M.S. Analysis of Carotenoid Production by Halorubrum sp. TBZ126; an Extremely Halophilic Archeon from Urmia Lake. Adv. Pharm. Bull. 2014, 4, 61–67. [Google Scholar] [PubMed] [Green Version]
- Britton, G.; Liaaen-Jensen, S.; Pfander, H. Carotenoids Handbook; Birkhauser Verlag: Basel, Switzerland, 2012; pp. 456–460. [Google Scholar]
- Instituto Nacional de Ecología y Cambio Climático. Available online: http://www2.inecc.gob.mx/publicaciones2/libros/421/cap2.html#:~:text=Se%20distribuyen%20principalmente%20en%20cuatro,la%20Sierra%20Madre%20del%20Sur (accessed on 26 January 2021).
- Ortiz, E. Características Edafológicas, Fisiográficas, Climáticas e Hidrográficas de México; Instituto Nacional de estadística y geografía INEGI: Aguascalientes, Mexico, 2008. [Google Scholar]
- Llanura Costera del Pacífico: Hidrología, Clima, Flora, Fauna. Available online: https://www.lifeder.com/llanura-costera-pacifico/ (accessed on 26 January 2021).
- Viver, T.; Cifuentes, A.; Díaz, S.; Rodríguez-Valdecantos, G.; González, B.; Antón, J.; Rosselló-Mora, R. Diversity of extremely halophilic cultivable prokaryotes in Mediterranean, Altlantic and Pacific solar salterns: Evidence that unexplored sites constitute sources of cultivable novelty. Syst. Appl. Microbiol. 2015, 38, 266–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Louyakis, A.; Makkay, A.; Guerrero, R.; Papke, T.; Gogarten, P. Complete Genome Sequence of Halorubrum ezzemoulense Strain Fb21. Microbiol. Resour. Announc. 2019, 8, e00096-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GenBank NCBI. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/nuccore/HM031393.1 (accessed on 11 May 2021).
- Atanasova, N.; Roine, E.; Oren, A.; Bamford, D.; Oksanen, H. Global network of specific virus-host interactions in hypersaline environments. Environ. Microbiol 2012, 14, 426–440. [Google Scholar] [CrossRef] [PubMed]
- Oren, A. The Microbiology of Red Brines. In Advances in Applied Microbiology, 1st ed.; Jackman, A.M., Johnson, H., Eds.; Elsevier: London, UK, 2020; Volume 113, pp. 57–110. [Google Scholar]
- Sahli, K.; Gomri, M.; Esclapez, J.; Gómez-Villegas, P.; Ghennai, O.; Bonete, M.; León, R.; Kharroub, K. Bioprospecting and characterization of pigmented halophilic archaeal strains from Algerian hypersaline environments with analysis of carotenoids produced by Halorubrum sp. BS2. J. Basic Microbiol. 2020, 60, 624–638. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Huo, Y.Y.; Zhang, X.Q. Halolamina salifodinae sp. nov. and Halolamina salina sp. nov. two extremely halophilic archaea isolated from a salt mine. Int. J. Syst. Evol. Microbiol. 2013, 63, 4380–4385. [Google Scholar] [PubMed]
- De la Vega, M.; Sayago, A.; Ariza, J.; Barneto, A.G.; León, R. Characterization of a bacterioruberin-producing Haloarchaea isolated from the marshlands of the Odiel river in the southwest of Spain. Biotechnol. Prog. 2016, 32, 592–600. [Google Scholar] [CrossRef]
- Cui, H.; Gao, X.; Yang, X.; Xu, X. Halolamina pelagica gen. nov., sp. nov., a new member of the family Halobacteriaceae. Int. J. Syst. Evol. Microbiol. 2011, 61, 1617–1621. [Google Scholar] [CrossRef]
- Parks, D.H.; Chuvochina, M.; Chaumeil, P.A.; Rinke, C.; Mussig, A.J.; Hugenholtz, P. A complete domain-to-species taxonomy for Bacteria and Archaea. Nat. Biotechnol. 2020, 38, 1079–1086. [Google Scholar] [CrossRef]
- Tang, L. Taxonomy of Bacteria and Archaea. Nat. Methods 2020, 17, 562. [Google Scholar] [CrossRef] [PubMed]
- Spang, A.; Caceres, E.; Ettema, T. Genomic exploration of the diversity, ecology, and evolution of the archaeal domain of life. Science 2017, 357, eaaf3883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Mu, Q.; Hu, K.; Chen, M.; Yang, J.; Chen, J.; Sun, Z. Characterization of MK 8 (H2) from Rhodococcus sp. B7740 and its potential antiglycation capacity measurements. Mar. Drugs 2018, 16, 391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barreteau, H.; Vandervennet, M.; Guèdon, L.; Point, V.; Canaan, S.; Rebuffat, S.; Peduzzi, J.; Carrè-Mlouka, A. Haloarcula sebkhae sp. nov., an extremely halophilic archaeon from Algerian hypersaline environment. Int. J. Syst. Evol. Microbiol. 2019, 69, 732–738. [Google Scholar] [CrossRef]
- Namwong, S.; Tanasupawat, S.; Kudo, T.; Itoh, T. Haloarcula salaria sp. Nov. and Haloarcula tradensis sp. nov., isolated from salt in Thai fish sauce. Int. J. Syst. Evol. Microbiol. 2011, 61, 231–236. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Cui, H.L.; Zhou, P.J.; Liu, S.J. Haloarcula amylolytica sp. nov., an extremely halophilic archaeon isolated from Aibi Salt Lake in Xin-Jiang, China. Int. J. Syst. Evol. Microbiol. 2007, 57, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.; Nurcahyanto, D.; Kawasaki, H.; Lisdiyanti, P.; Suzuki, K.I. Halobium palmae gen. nov., sp. nov., an extremely halophilic archaeon isolated from a solar saltern. Int. J. Syst. Evol. Microbiol. 2016, 66, 3799–3804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yim, K.; Cha, I.; Lee, H.; Song, H.; Kim, K.; Lee, S.; Nam, Y.; Hyun, D.; Bae, J.; Rhee, S.; et al. Halorubrum halophilum sp. nov., an extremely halophilic archaeon isolated from a salt-fermented seafood. Anton Leeuw 2014, 105, 603–612. [Google Scholar] [CrossRef] [PubMed]
- Collins, M.; Tindall, B. Ocurrence of menaquinones and some novel methylated menaquinones in the alkaliphilic, extremely halophilic archaebacterium Natronobacterium gregoryi. FEMS Microbiol. Lett. 1987, 43, 307–312. [Google Scholar] [CrossRef]
- Oren, A. Halophilic Microorganisms and Their Environments; Kluwer Academic Publishers: Norwell, MA, USA; Hebrew University of Jerusalem: Jerusalem, Israel, 2003; Volume 5, pp. 91–92. [Google Scholar]
- Oberwinkler, T. Metabolic and Genomic Annotations in Halophilic Archaea. Ph.D. Thesis, Fakultät für Chemie und Pharmazie der Ludwig-Maximilians-Universität München, München, Germany, 2011. [Google Scholar]
- Flores, N.; Hoyos, S.; Venegas, M.; Galetovic, A.; Zúñiga, L.; Fábrega, F.; Paredes, B.; Salazar-Ardilles, C.; Vilo, C.; Ascaso, C.; et al. Haloterrigena sp. strain SGH1, a bacterioruberin-rich, perchlorate-tolerant halophilic archaeon isolated from halite microbial communities, Atacama Desert, Chile. Front. Microbiol. 2020, 11, 1–18. [Google Scholar] [CrossRef]
- Falb, M.; Müller, K.; Königsmaier, L.; Oberwinkler, T.; Horn, P.; Von Gronau, S.; Oesterhelt, D. Metabolism of halophilic archaea. Extremophiles 2008, 12, 177–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torreblanca, M.; Rodriguez-Valera, F.; Juez, G.; Ventosa, A.; Kamekura, M.; Kates, M. Classification of Non-alkaliphilic Halobacteria Based on Numerical Taxonomy and Polar Lipid Composition, and Description of Haloarcula gen. nov. and Haloferax gen. nov. Syst. Appl. Microbiol. 1986, 8, 89–99. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains Code | Closest Strains in GenBank Database | Accession Numbers | Percentage of Similarity | Reference |
|---|---|---|---|---|
| M1 CA_13B53 | Haloarcula sp. M1 Haloarcula salaria | MW567147 LN649977.1 | 100% 99.4% | From this study [35] |
| M2 Fb21 | Halorubrum sp. M2 Halorubrum ezzemoulense | MW567148 CP034940.1 | 100% 99.3% | From this study [36] |
| M3 UAH-SP14 | Halolamina sp. M3 Haloarchaeon UA-SP14 | MW567149 HM031393.1 | 100% 99.46% | From this study [37] |
| M4 SD683 | Halorubrum sp. M4 Halorubrum sp. SD683 | MW567151 LT578362.2 | 100% 99.85% | From this study [38] |
| M5 Fb21 | Halorubrum sp. M5 Halorubrum ezzemoulense | MW567150 CP034940.1 | 100% 93.3% | From this study [36] |
| M6 Fb21 | Halorubrum sp. M6 Halorubrum ezzemoulense | MW567152 CP034940.1 | 100% 99.85% | From this study [36] |
| M7 Fb21 | Halorubrum sp. M7 Halorubrum ezzemoulense | MW56567153 CP034940.1 | 100% 100% | From this study [36] |
| M8 E302-1 | Halorubrum sp. M8 Halorubrum sp. E302-1 | MW567154 JN196504.1 | 100% 99.17% | From this study [38] |
| Strain ID | Quinone | Glycolipids | Phospholipids |
|---|---|---|---|
| Haloarcula sp. M1 | MK-8 | 2 UK, 1 GP | PG, PGP-Me, PI-Na, 3 UK |
| Halorubrum sp. M2 | MK-8, MK-8(H2) | 1 UK | PG, PGP-Me, PI-Na, 3 UK |
| Halolamina sp. M3 | MK-8, MK-8(H2) | S-DGD, 1 UK, 1 GP | PG, PGP-Me, PI-Na, 2 UK |
| Halorubrum sp. M4 | MK-8, MK-8(H2) | 2 UK, 1 GP | PG, PGP-Me, PI-Na, 3 UK |
| Halorubrum sp. M5 | MMK-8 | 1 UK | PG, PGP-Me, PI-Na, 3 UK |
| Halorubrum sp. M6 | MK-8, MK-8(H2) | 2 UK | PG, PGP-Me, PI-Na, 3 UK |
| Halorubrum sp. M7 | MK-8, MK-8(H2) | 1 UK | PG, PGP-Me, PI-Na, 3 UK |
| Halorubrum sp. M8 | MK-8 | 1 UK | PG, PGP-Me, PI-Na, 3 UK |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vázquez-Madrigal, A.S.; Barbachano-Torres, A.; Arellano-Plaza, M.; Kirchmayr, M.R.; Finore, I.; Poli, A.; Nicolaus, B.; De la Torre Zavala, S.; Camacho-Ruiz, R.M. Effect of Carbon Sources in Carotenoid Production from Haloarcula sp. M1, Halolamina sp. M3 and Halorubrum sp. M5, Halophilic Archaea Isolated from Sonora Saltern, Mexico. Microorganisms 2021, 9, 1096. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9051096
Vázquez-Madrigal AS, Barbachano-Torres A, Arellano-Plaza M, Kirchmayr MR, Finore I, Poli A, Nicolaus B, De la Torre Zavala S, Camacho-Ruiz RM. Effect of Carbon Sources in Carotenoid Production from Haloarcula sp. M1, Halolamina sp. M3 and Halorubrum sp. M5, Halophilic Archaea Isolated from Sonora Saltern, Mexico. Microorganisms. 2021; 9(5):1096. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9051096
Chicago/Turabian StyleVázquez-Madrigal, Ana Sofía, Alejandra Barbachano-Torres, Melchor Arellano-Plaza, Manuel Reinhart Kirchmayr, Ilaria Finore, Annarita Poli, Barbara Nicolaus, Susana De la Torre Zavala, and Rosa María Camacho-Ruiz. 2021. "Effect of Carbon Sources in Carotenoid Production from Haloarcula sp. M1, Halolamina sp. M3 and Halorubrum sp. M5, Halophilic Archaea Isolated from Sonora Saltern, Mexico" Microorganisms 9, no. 5: 1096. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9051096