
Evaluation of Probiotic Properties of Pediococcus acidilactici M76 Producing Functional Exopolysaccharides and Its Lactic Acid Fermentation of Black Raspberry Extract

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and Growth Conditions
2.2. Acid and Bile Tolerance
2.3. Antimicrobial Activity
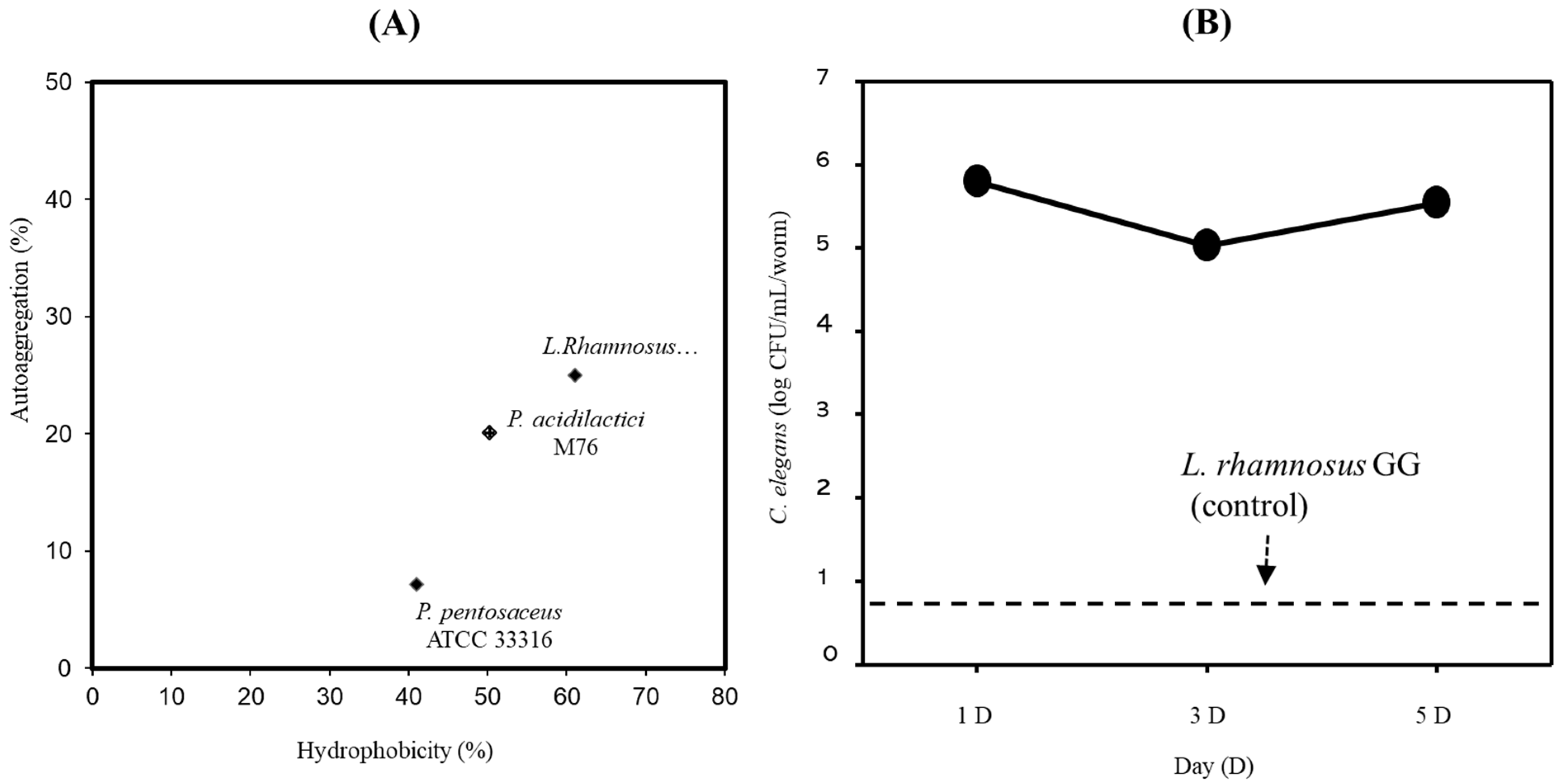
2.4. Cell Surface Hydrophobicity and Autoaggregation
2.5. Caenorhabditis elegans Intestinal Adhesion
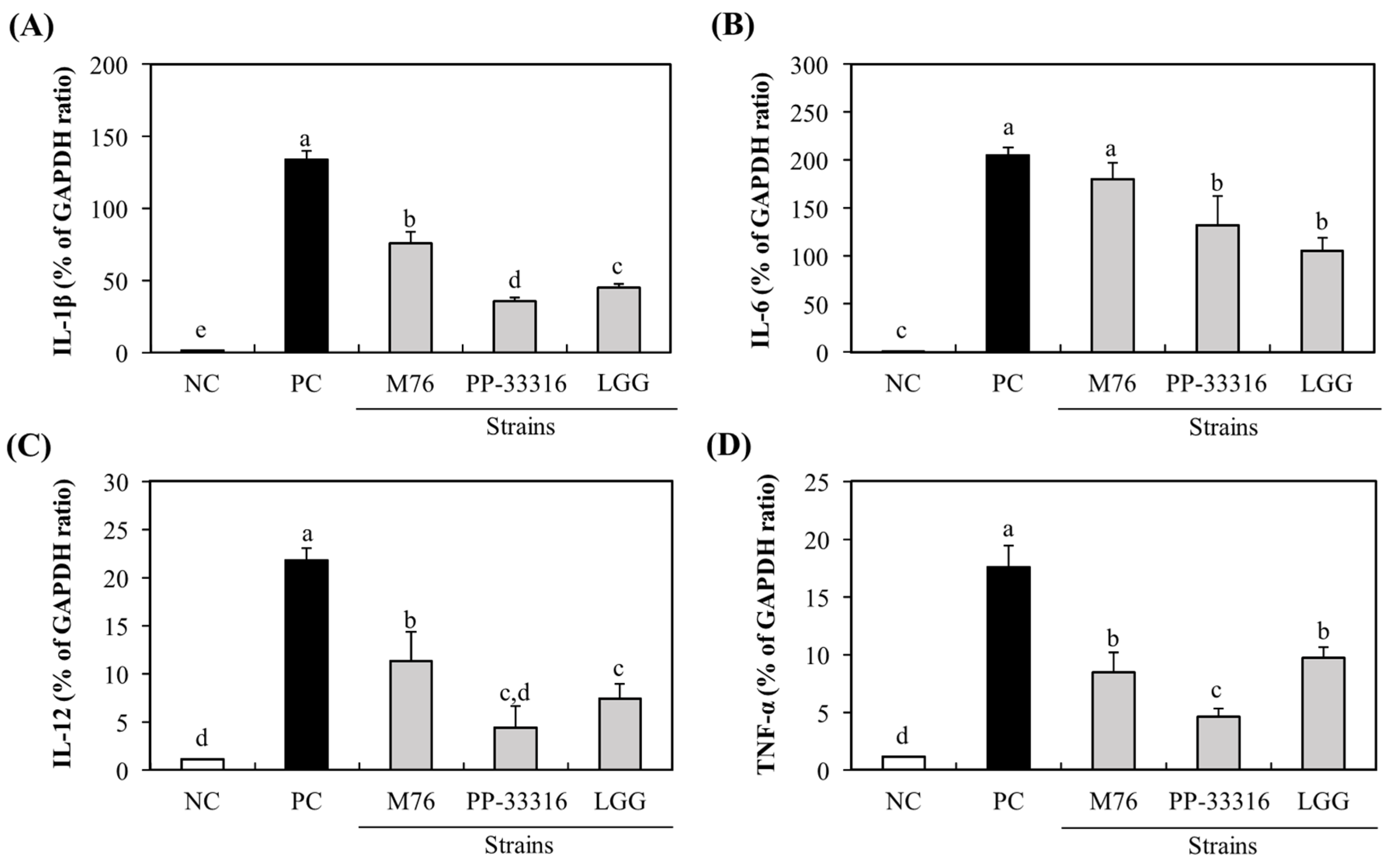
2.6. In Vitro Immuno-Stimulatory Effects
2.6.1. Cell Culture and Bacterial Stimulation
2.6.2. Cell Viability Assay
2.6.3. qRT-PCR Analysis
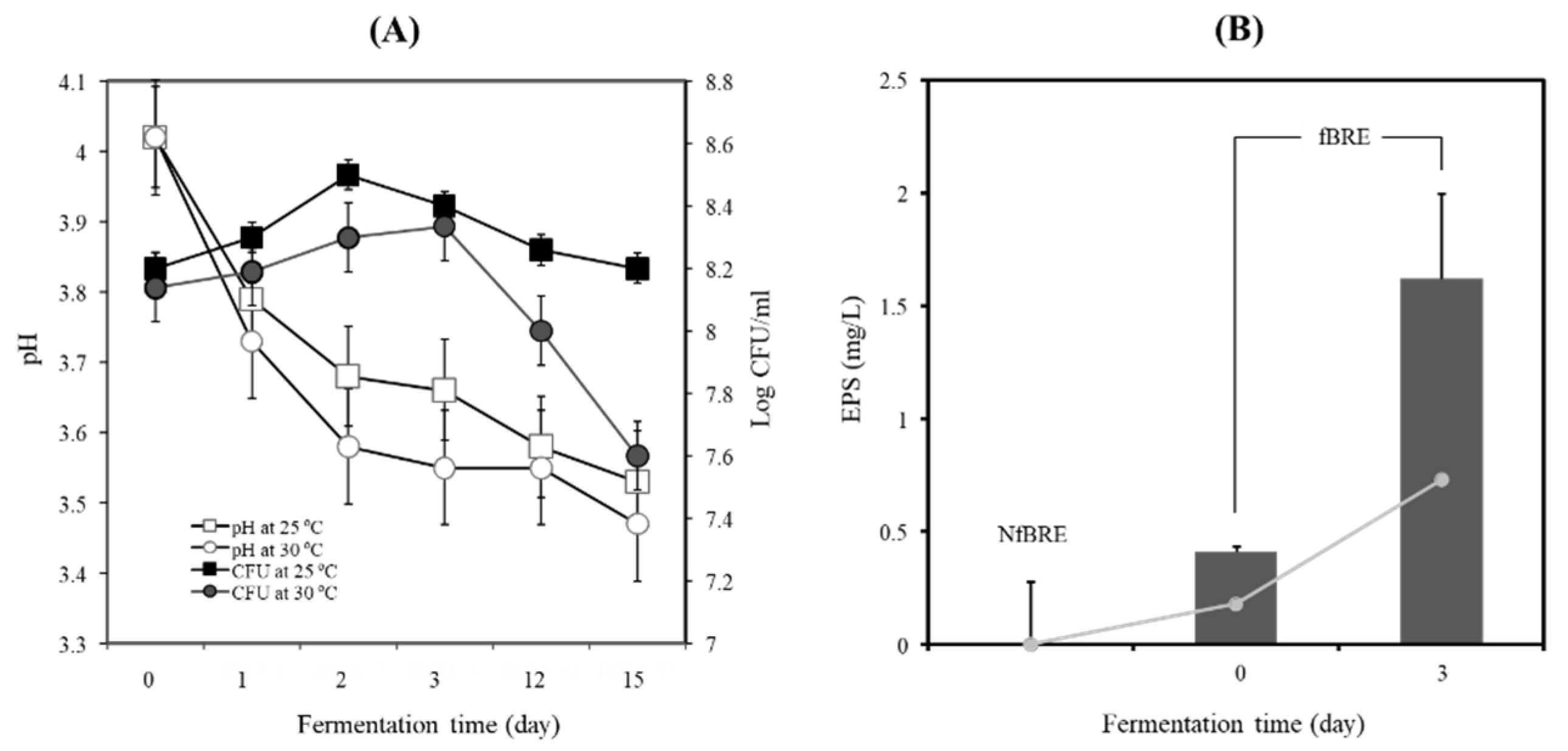
2.7. Fermentation of BRE with PA-M76
2.8. Analysis of Cell Biomass and EPS
2.9. Total Anthocyanin, Phenol, and Flavonoid Analysis
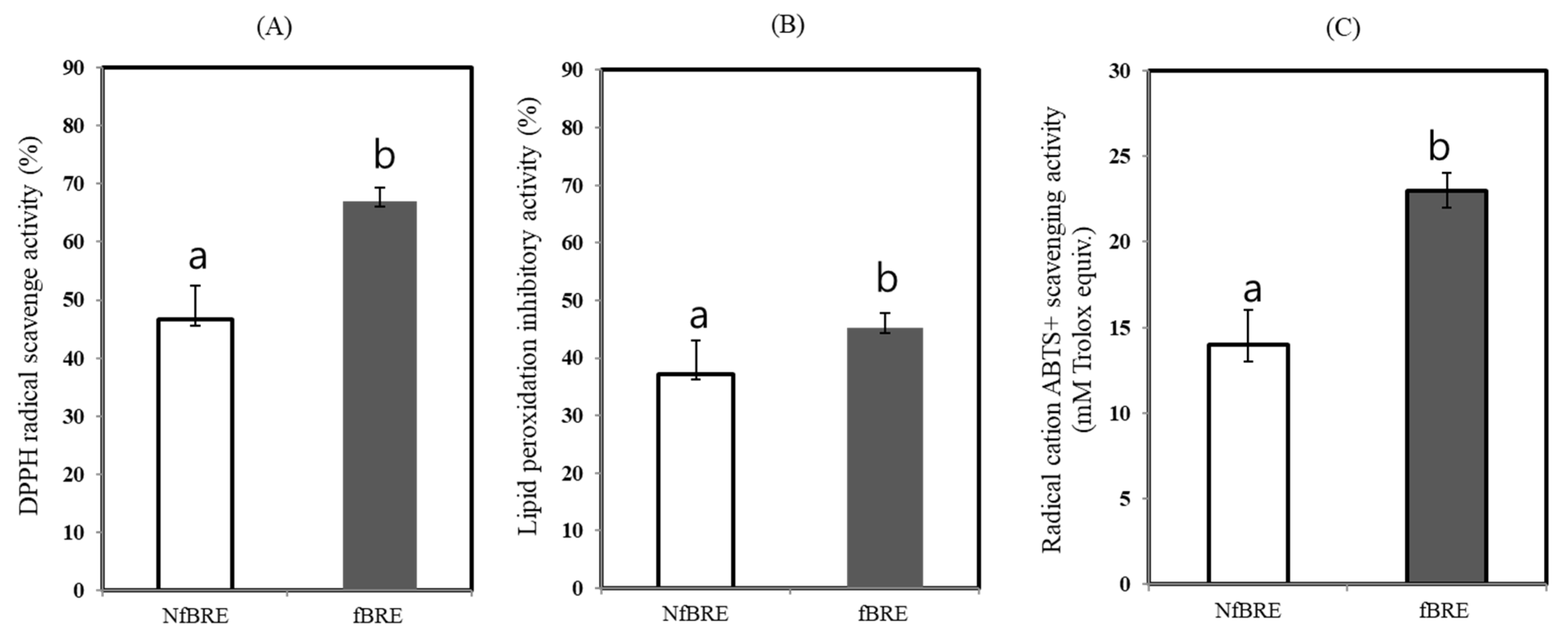
2.10. In Vitro Antioxidant Activity
2.10.1. DPPH Radical Scavenging Activity
2.10.2. Lipid Peroxidation Inhibitory Activity
2.10.3. Radical Cation ABTS+ Scavenging Activity
2.11. Headspace Solid-Phase Microextraction (SPME) and Gas Chromatography-Mass Spectrometry (GC-MS) Analysis
2.12. Statistical Analysis
3. Results and Discussion
3.1. Fundamental Probiotic Properties: Tolerance and Antimicrobial Activity
3.2. Adhesion Properties of PA-M76
3.3. Immuno-Stimulatory Activity of PA-M76 to RAW 264.7 Cells
3.4. Black Raspberry Fermentation by PA-M76 Enhanced Production of Functional EPS
3.5. Phenolic Compounds and Antioxidant Activity of fBRE
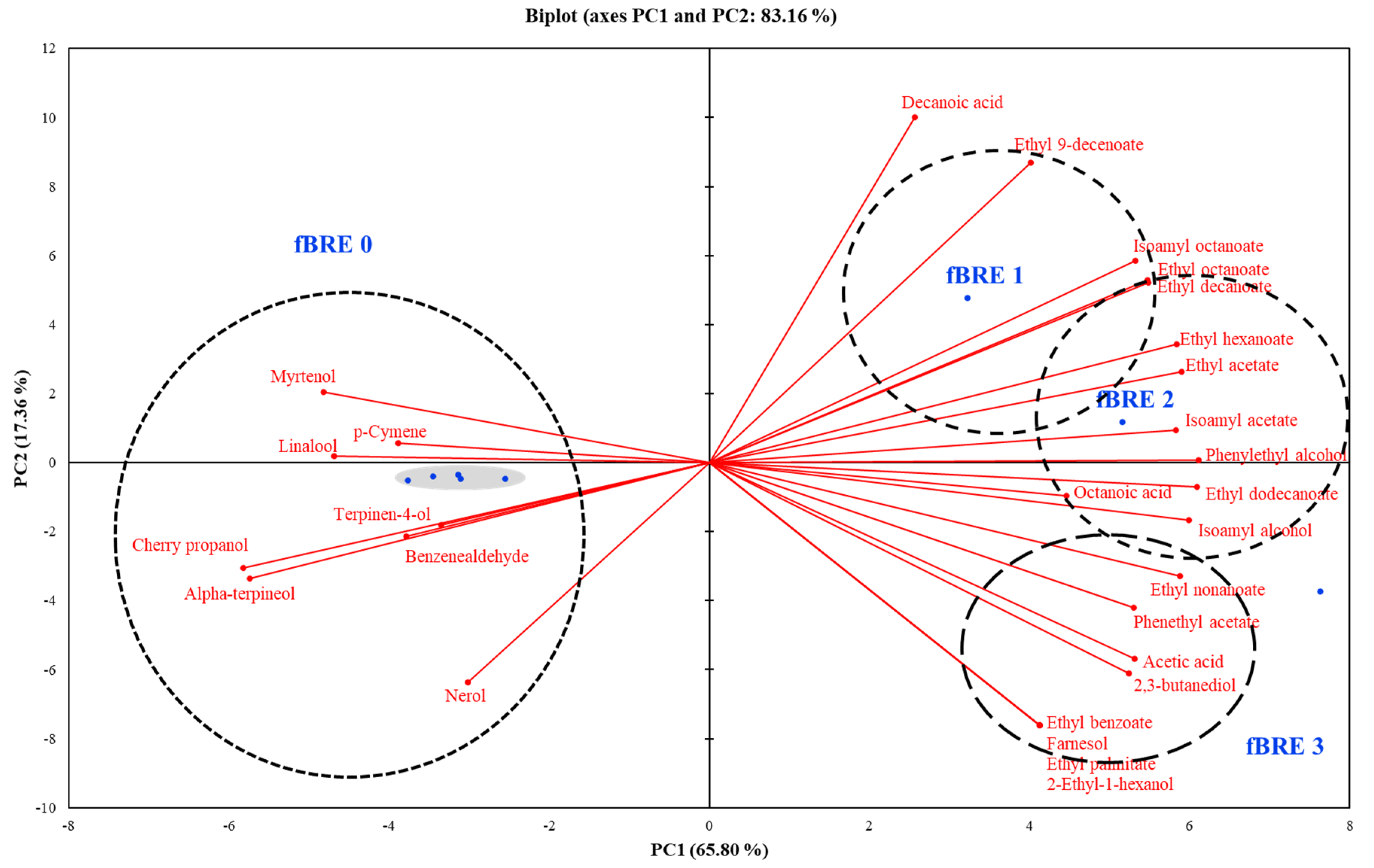
3.6. Production of Volatile Compounds during Fermentation of BRE with P. acidilactici M76
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviation
| BR | Black raspberry |
| BRE | Black raspberry extract |
| fBRE | Fermented black raspberry extract |
| NfBRE | Non-fermented black raspberry extract |
| EPS | Exopolysaccharide |
| GIT | Gut intestinal track |
| PA-M76 | Pediococcus acidilactici M76 |
| PP | P. pentosaceus ATCC 33316 |
| LGG | Lactobacillus rhamnosus GG |
| PCA | Principal component analysis |
References
- Ghoson, M.D.; Marwa, O.E.; Waill, A.E. Contributions of exopolysaccharides from lactic acid bacteria as biotechnological tools in food, pharmaceutical, and medical applications. Int. J. Biol. Macromol. 2021, 173, 79–89. [Google Scholar]
- Torino, M.; Valdez, G.F.; Mozzi, F. Biopolymers from lactic acid bacteria. Novel applications in foods and beverages. Front. Microbiol. 2015, 6, 834. [Google Scholar] [CrossRef] [Green Version]
- Wisselink, H.W.; Weusthuis, R.A.; Eggink, G.; Hugenholtz, J.; Grobben, G.J. Mannitol production by lactic acid bacteria: A review. Int. Dairy J. 2002, 12, 151–161. [Google Scholar] [CrossRef]
- Ogawa, J.; Kishino, S.; Ando, A.; Sugimoto, S.; Mihara, K.; Shimizu, S. Production of conjugated fatty acids by lactic acid bacteria. J. Biosci. Bioeng. 2005, 100, 355–364. [Google Scholar] [CrossRef]
- Cerning, J. Exocellular polysaccharides produced by lactic acid bacteria. FEMS Microbiol. Lett. 1990, 87, 113–130. [Google Scholar] [CrossRef]
- Nwodo, U.U.; Green, E.; Okoh, A.I. Bacterial exopolysaccharides: Functionality and prospects. Int. J. Mol. Sci. 2012, 13, 14002–14015. [Google Scholar] [CrossRef] [Green Version]
- Patel, A.; Lindström, C.; Patel, A.; Prajapati, J.; Holst, O. Probiotic properties of exopolysaccharide producing lactic acid bacteria isolated from vegetables and traditional Indian fermented foods. Int. J. Fermented Foods 2012, 1, 87–101. [Google Scholar]
- Oh, Y.J.; Jung, D.S. Evaluation of probiotic properties of Lactobacillus and Pediococcus strains isolated from Omegisool, a traditionally fermented millet alcoholic beverage in Korea. LWT Food Sci. Technol. 2015, 63, 437–444. [Google Scholar] [CrossRef]
- Almudena, G.R.; Dolores, G.L.; Adelaida, E.F.; Teresa, R.; Begona, B.M.; Victoria, M.A. Assessment of probiotic properties in lactic acid bacteria isolated from wine. Food Microbiol. 2014, 44, 220–225. [Google Scholar]
- Zannini, E.; Deborah, M.W.; Aidan, C.; Elke, K.A. Production, properties, and industrial food application of lactic acid bacteria-derived exopolysaccharides. Appl. Microbiol. Biotechnol. 2016, 100, 1121–1135. [Google Scholar] [CrossRef] [PubMed]
- Seeram, N.P. Berries. In Nutritional Oncology, 2nd ed.; Heber, D., Blackburn, G., Go, V.L.W., Milner, J., Eds.; Academic Press: London, UK, 2006; pp. 615–628. [Google Scholar]
- Puupponen-Pimia, R.; Nohynek, L.; Meier, C.; Kahkonen, M.; Heinonen, M.; Hopia, A.; Oksman-Caldentey, K.M. Antimicrobial properties of phenolic compounds from berries. J. Appl. Microbiol. 2001, 90, 494–507. [Google Scholar] [CrossRef] [PubMed]
- Moon, Y.J.; Baik, S.H.; Cha, Y.S. Lipid-lowering effects of Pediococcus acidilactici M76 isolated from Korean traditional Makgeolli in high fat diet-induced obese mice. Nutrients 2014, 6, 1016–1028. [Google Scholar] [CrossRef] [Green Version]
- Maria, D.; Aline, L.F.; Marguerite, D.L. Chapter 12: Polysaccharide production by grapes must and wine microorganisms. In Biology of Microorganisms on Grapes, in Must and in Wine; Springer International Publishing AG: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Rishikesh, S.; Arun, G. Probiotic potential of Pediococcus pentosaceus CRAG3: A new isolate from fermented cucumber probiotics and antimicrobial proteins. Probiotics Antimicrob. Proteins 2014, 6, 11–21. [Google Scholar]
- Sim, I.S.; Park, K.T.; Kwon, G.; Koh, J.H.; Lim, Y.H. Probiotic potential of Enterococcus faecium isolated from chicken cecum with immunomodulating activity and promoting longevity in Caenorhabditis elegans. J. Microbiol. Biotechnol. 2018, 28, 883–892. [Google Scholar] [CrossRef] [Green Version]
- Namiki, M.; Osawa, T. Antioxidants/antimutagens in foods. Basic Life Sci. 1986, 39, 131–142. [Google Scholar]
- Abushelaibi, A.; Al-Mahadin, S.; El-Tarabily, K.; Shah, N.P.; Ayyasha, M. Characterization of potential probiotic lactic acid bacteria isolated from camel milk. LWT Food Sci. Technol. 2017, 79, 316–325. [Google Scholar] [CrossRef]
- Mathara, J.M.; Schillinger, U.; Kutima, P.M.; Mbugua, S.K.; Guigas, C.; Franz, C.; Holzapfel, W.H. Functional properties of Lactobacillus plantarum strains isolated from maasai traditional fermented milk products in Kenya. Curr. Microbiol. 2008, 56, 315–321. [Google Scholar] [CrossRef]
- de Palencia, P.F.; Werning, M.L.; Sierra-Filardi, E.; Dueñas, M.T.; Irastorza, A.; Corbí, A.L.; López, P. Probiotic properties of the 2-substituted (1,3)-β-d-glucan-producing bacterium Pediococcus parvulus 2.6. Appl. Environ. Microbiol. 2009, 75, 4887–4891. [Google Scholar] [CrossRef] [Green Version]
- Park, M.R.; Yun, H.S.; Son, S.J.; Oh, S.; Kim, Y. Short communication: Development of a direct in vivo screening model to identify potential probiotic bacteria using Caenorhabditis elegans. J. Dairy Sci. 2014, 97, 6828–6834. [Google Scholar] [CrossRef] [Green Version]
- Poupet, C.; Saraoui, T.; Veisseire, P.; Bonnet, M.; Dausset, C.; Gachinat, M.; Camarès, O.; Chassard, C.; Nivoliez, A.; Bornes, S. Lactobacillus rhamnosus Lcr35 as an effective treatment for preventing Candida albicans infection in the invertebrate model Caenorhabditis elegans: First mechanistic insights. PLoS ONE 2019, 14, e0216184. [Google Scholar] [CrossRef]
- Lee, H.K.; Choi, S.H.; Lee, C.R.; Lee, S.H.; Park, M.R.; Kim, Y.H.; Lee, M.K.; Kim, G.B. Screening and characterization of Lactic Acid Bacteria Strains with anti-inflammatory activities through in vitro and Caenorhabditis elegans model testing. Korean J. Food Sci. Anim. Resour. 2015, 35, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.R.; Jeong, D.Y.; Cha, Y.S.; Baik, S.H. Exopolysaccharides produced by Pediococcus acidilactici M76 isolated from the traditional rice wine, Makgeolli. J. Microbiol. Biotechnol. 2013, 23, 681–688. [Google Scholar] [CrossRef]
- Osmanagaoglu, O.; Kiran, F.; Yagci, F.C.; Gursel, I. Immunomodulatory function and in vivo properties of Pediococcus pentosaceus OZF, a promising probiotic strain. Ann. Microbiol. 2013, 63, 1311–1318. [Google Scholar] [CrossRef]
- Ashraf, R.; Shah, N.P. Immune System Stimulation by Probiotic Microorganisms. Crit. Rev. Food Sci. Nutri. 2014, 54, 938–956. [Google Scholar] [CrossRef]
- Jonganurakkun, B.; Wang, Q.; Xu, S.H.; Tada, Y.; Minamida, K.; Yasokawa, D.; Sugi, M.; Hara, H.; Asano, K. Pediococcus pentosaceus NB-17 for probiotic use. J. Biosci. Bioeng. 2008, 106, 69–73. [Google Scholar] [CrossRef]
- Cao, Z.; Pan, H.; Tong, H.; Gu, D.; Li, S.; Xu, Y.; Ge, C.; Lin, Q. In vitro evaluation of probiotic potential of Pediococcus pentosaceus L1 isolated from paocai a Chinese fermented vegetable. Ann. Microbiol. 2016, 66, 963–971. [Google Scholar] [CrossRef]
- Cerning, J. Production of exopolysaccharides by lactic acid bacteria and dairy propionibacteria. Le Lait 1995, 75, 463–472. [Google Scholar] [CrossRef]
- Zisu, B.; Shah, N.P. Effects of pH, temperature, supplementation with whey protein concentrate, and adjunct cultures on the production of exopolysaccharides by Streptococcus thermophilus 1275. J. Dairy Sci. 2003, 86, 3405–3415. [Google Scholar] [CrossRef] [Green Version]
- Vaningelgem, F.; Zamfir, M.; Mozzi, F.; Adriany, T.; Vancanneyt, M.; Swings, J.; De Vuyst, L. Biodiversity of exopolysaccharides produced by Streptococcus thermophiles strains is reflected in their production and their molecular and functional characteristics. Appl. Environ. Microbiol. 2004, 70, 900–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, H.; Curiel, J.A.; Landete, J.M.; de las Rivas, B.; de Felipe, F.L.; Gómez-Cordovés, C.; Mancheño, J.M.; Muñoz, R. Food phenolics and lactic acid bacteria. Int. J. Food Microbiol. 2009, 132, 79–90. [Google Scholar] [CrossRef] [Green Version]
- Kwaw, E.; Ma, Y.; Tchabo, W.; Apaliya, M.T.; Wu, M.; Sackey, A.S.; Xiao, L.; Tahir, H.E. Effect of lactobacillus strains on phenolic profile, color attributes and antioxidant activities of lactic-acid-fermented mulberry juice. Food Chem. 2018, 250, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wang, R.; Zhang, Y.; Yang, Y.; Sun, X.H.; Zhang, Q.; Yang, N. Biotransformation of phenolics and metabolites and the change in antioxidant activity in kiwi fruit induced by Lactobacillus plantarum fermentation. J. Sci. Food Agric. 2020, 100, 3283–3290. [Google Scholar] [CrossRef]
- Qian, Z.J.; Jung, W.K.; Kim, S.K. Free radical scavenging activity of a novel antioxidative peptide purified from hydrolysate of bullfrog skin, Rana catesbeiana Shaw. Bioresour. Technol. 2008, 6, 1690–1698. [Google Scholar] [CrossRef] [PubMed]
- Escudero, A.; Gogorza, B.; Melus, M.A.; Ortin, N.; Cacho, J.; Ferreira, V. Characterization of the aroma of a wine from Maccabeo. Key role played by compounds with low odor activity values. J. Agric. Food Chem. 2004, 52, 3516–3524. [Google Scholar] [CrossRef]
- Duarte, W.F.; Dias, D.R.; Oliveira, J.M.; Vilanova, M.; Teixeira, J.A.; e Silva, J.B.A.; Schwan, R.F. Raspberry (Rubus idaeus L.) wine: Yeast selection, sensory evaluation and instrumental analysis of volatile and other compounds. Food Res. Int. 2010, 43, 2303–2314. [Google Scholar] [CrossRef]
- Guth, H. Quantitation and sensory studies of character impact odorants of different white wine varieties. J. Agric. Food Chem. 1997, 45, 3027–3032. [Google Scholar] [CrossRef]
- Vilanova, M.; Campo, E.; Escudero, A.; Graña, M.; Masa, A.; Cacho, J. Volatile composition and sensory properties of Vitis vinifera red cultivars from North West Spain: Correlation between sensory and instrumental analysis. Anal. Chim. Acta 2012, 720, 104–111. [Google Scholar] [CrossRef]
- Lee, S.J.; Lee, J.E.; Kim, H.W.; Kim, S.S.; Koh, K.H. Development of Korean red wines using Vitis labrusca varieties: Instrumental and sensory characterization. Food Chem. 2006, 94, 385–393. [Google Scholar] [CrossRef]
- Choi, H.S. Headspace analysis of Robus coreanus berry by solid-phase microextraction and its sniffing test by gas chromatography-olfactometry. Food Sci. Biotechnol. 2002, 11, 355–360. [Google Scholar]
- Nykänen, L.; Suomalainen, H. Aroma of Beer, Wine and Distilled Alcoholic Beverages; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1983; Volume 3. [Google Scholar]
- Liberatore, M.T.; Pati, S.; Del Nobile, M.A.; La Notte, E. Aroma quality improvement of Chardonnay white wine by fermentation and ageing in barrique on lees. Food Res. Int. 2010, 43, 996–1002. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tolerance | PA-M76 | LGG | P. pentosaceus 33316 |
|---|---|---|---|
| pH 7.0 | 111.7 ± 0.58 | 110.7 ± 0.76 | 111.4 ± 0.35 |
| pH 2.5 | 71.7 ± 0.22 | 72.5 ± 0.42 | 74.0 ± 0.08 |
| Bile salt 0.3% | 101.8 ± 0.92 | 97.9 ± 0.78 | 95.9 ± 0.61 |
| Bile salt 1% | 104.0 ± 1.11 | 93.3 ± 0.53 | 94.1 ± 0.78 |
| Bile salt 3% | 101.4 ± 0.15 | 80.9 ± 1.73 | 89.1 ± 1.33 |
| Temperature | 35~50 | 35~50 | 35~50 |
| NaCl (%, w/v) | 4~8 | 4~7 | 4~8 |
| Glucose (%, w/v) | 4~14 | 4~14 | 4~14 |
| Strain | Pathogenic Microorganisms | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| S. aureus | S. epidermidis | S. xylosus | P. aeruginosa | P. putida | B. cereus | B. subtilis | B. vallismortis | E. coli | P. acnes | |
| L. Rhamnosus GG * | ++++ | + | +++ | ++ | ++ | ++ | + | +++ | + | ++ |
| P. acidilactici M76 | ++++ | +++ | +++ | +++ | +++ | ++++ | ++ | +++ | ++++ | +++ |
| P. pentosaceus ATCC 33316 | ++++ | + | +++ | ++ | +++ | ++ | ++ | +++ | +++ | ++ |
| Phenolic compounds | NfBRE | fBRE |
|---|---|---|
| Total polyphenols (mg GAE/100g) | 349.4 ± 3.99 a | 379.3 ± 4.38 b |
| Total flavonoids (mg CE/100g) | 99.9 ± 2.37 a | 81 ± 0.82 a |
| Total anthocyanins (TAC mg/L) | 43 ± 0.67 a | 45 ± 0.14 a |
| Aroma Compound | RT (1) | Non Fermented Black Raspberry Extract (Day) | Fermented Black Raspberry Extract (Day) | Odor Description (2) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 0 | 1 | 2 | 3 | |||
| Acids | ||||||||||
| Acetic acid | 2.77 | nd (3) | nd | nd | nd | nd | nd | 0.03 ± 0.01 ab | 0.05 ± 0.03 a | Sour |
| Octanoic acid | 6.63 | nd | nd | nd | nd | nd | nd | 0.18 ± 0.16 a | 0.08 ± 0.07 ab | Fatty, waxy, cheesy |
| Decanoic acid | 8.01 | nd | 0.01 ± 0.01 | 0.01 ± 0.00 | nd | nd | 0.32 ± 0.28 | 0.07 ± 0.06 | 0.02 ± 0.01 | Rancid, sour, fatty, citrus |
| Alcohols | ||||||||||
| Isoamyl alcohol | 2.93 | nd | nd | nd | nd | nd | 0.65 ± 0.19 b | 1.48 ± 0.25 a | 1.75 ± 0.34 a | Alcoholic, whiskey, fruity |
| 2,3-Butanediol | 3.62 | nd | nd | nd | nd | nd | nd | 0.01 ± 0.01 ab | 0.02 ± 0.01 a | Fruity, creamy, buttery |
| 2-Ethyl-1-hexanol | 5.48 | nd | nd | nd | nd | nd | nd | nd | 0.05 ± 0.00 | Citrus, floral, oily, sweet |
| Phenylethyl alcohol | 6.25 | 0.02 ± 0.00 b | 0.02 ± 0.01 Ba | 0.02 ± 0.01 Ba | 0.02 ± 0.01 Ba | 0.05 ± 0.02 Ab | 0.40 ± 0.25 Aab | 0.48 ± 0.21 Aa | 0.64 ± 0.31 Aa | Floral, rose, dried rose |
| Cherry propanol | 6.79 | 0.15 ± 0.01 | 0.17 ± 0.03 | 0.16 ± 0.04 | 0.15 ± 0.02 | 0.12 ± 0.02 | nd | nd | nd | Sweet, fruity, cherry |
| Aldehyde | ||||||||||
| Benzealdehyde | 4.95 | 0.01 ± 0.01 Ab | 0.03 ± 0.00 Aa | 0.04 ± 0.00 a | 0.04 ± 0.00 a | nd | nd | nd | nd | Sharp, sweet, bitter, almond |
| Esters | ||||||||||
| Ethyl acetate | 2.11 | nd | nd | nd | nd | nd | 1.99 ± 0.30 | 1.60 ± 0.85 | 2.08 ± 0.54 | Ethereal, fruity, sweet, grape |
| Isoamyl acetate | 4.14 | nd | nd | nd | nd | nd | 0.30 ± 0.07 b | 0.63 ± 0.11 a | 0.49 ± 0.11 a | Sweet, banana, fruity |
| Ethyl hexanoate | 5.22 | nd | nd | nd | nd | nd | 0.15 ± 0.12 | 0.13 ± 0.10 | 0.14 ± 0.12 | Sweet, fruity, pineapple |
| Ethyl benzoate | 6.67 | nd | nd | nd | nd | nd | nd | nd | 0.05 ± 0.01 | Fruity, sweet, wintergreen, minty |
| Ethyl octanoate | 6.82 | nd | nd | nd | nd | nd | 1.11 ± 0.94 | 0.89 ± 0.73 | 0.75 ± 0.60 | Fruity, winey, waxy, sweet |
| Phenethyl acetate | 7.31 | nd | nd | nd | nd | nd | nd | 0.17 ± 0.15 | 0.18 ± 0.16 | Floral, rose, sweet, honey |
| Ethyl nonanoate | 7.53 | nd | nd | nd | nd | nd | 0.02 ± 0.02 | 0.04 ± 0.04 | 0.07 ± 0.05 | Fruity, rose, waxy, rummy |
| Ethyl 9-decenoate | 8.16 | nd | nd | nd | nd | nd | 0.50 ± 0.44 | 0.23 ± 0.20 | 0.13 ± 0.11 | Fruity, fatty |
| Ethyl decanoate | 8.21 | nd | 0.04 ± 0.05 | 0.03 ± 0.04 | 0.01 ± 0.02 | nd | 0.52 ± 0.44 | 0.45 ± 0.38 | 0.36 ± 0.30 | Sweet, waxy, fruity, apple |
| Isoamyl octanoate | 8.55 | nd | nd | nd | nd | nd | 0.05 ± 0.04 | 0.04 ± 0.04 | 0.03 ± 0.02 | Sweet, oily, fruity, green |
| Ethyl dodecanoate | 9.46 | nd | nd | nd | nd | nd | 0.05 ± 0.05 | 0.08 ± 0.06 | 0.10 ± 0.08 | Sweet, waxy, floral, soapy |
| Ethyl palmitate | 11.6 | nd | nd | nd | nd | nd | nd | nd | 0.01 ± 0.00 | Waxy, fruity, creamy, milky |
| Terpenes | ||||||||||
| p-Cymene | 5.5 | 0.07 ± 0.01 a | 0.03 ± 0.00 Ab | 0.02 ± 0.02 b | nd | 0.05 ± 0.01 a | 0.02 ± 0.00 Bb | nd | nd | Fresh, citrus, terpenic |
| Linalool | 6.08 | 0.09 ± 0.01 a | 0.06 ± 0.01 Ab | 0.05 ± 0.01 Abc | 0.03 ± 0.00 Ac | 0.07 ± 0.02 a | 0.04 ± 0.01 Bb | 0.02 ± 0.01 Bb | 0.02 ± 0.00 Bb | Citrus, floral, sweet |
| Terpinen-4-ol | 6.76 | nd | 0.18 ± 0.01 a | 0.16 ± 0.02 b | 0.12 ± 0.01 c | nd | nd | nd | nd | Peppery, woody, earthy |
| alpha-terpineol | 6.85 | 0.35 ± 0.02 a | 0.34 ± 0.02 Aab | 0.31 ± 0.03 Abc | 0.27 ± 0.02 Ac | 0.29 ± 0.03 a | 0.14 ± 0.13 Bb | 0.13 ± 0.04 Bb | 0.15 ± 0.02 Bb | Pine, terpenic, lilac, citrus |
| Myrtenol | 6.91 | 0.18 ± 0.01 a | 0.16 ± 0.01 ab | 0.13 ± 0.01 b | 0.11 ± 0.01 c | 0.18 ± 0.02 a | 0.13 ± 0.06 ab | 0.10 ± 0.03 b | 0.08 ± 0.02 b | Woody, pine, balsamic |
| Nerol | 7.25 | 0.02 ± 0.00 a | 0.01 ± 0.00 b | 0.01 ± 0.00 c | 0.01 ± 0.00 a | 0.02 ± 0.00 a | nd | 0.01 ± 0.00 b | 0.01 ± 0.00 b | Fresh, citrus, floral |
| Farnesol | 10.24 | nd | nd | nd | nd | nd | nd | nd | 0.01 ± 0.01 | Sweet, floral |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, Y.-R.; Lee, C.-M.; Lee, S.-H.; Baik, S.-H. Evaluation of Probiotic Properties of Pediococcus acidilactici M76 Producing Functional Exopolysaccharides and Its Lactic Acid Fermentation of Black Raspberry Extract. Microorganisms 2021, 9, 1364. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071364
Song Y-R, Lee C-M, Lee S-H, Baik S-H. Evaluation of Probiotic Properties of Pediococcus acidilactici M76 Producing Functional Exopolysaccharides and Its Lactic Acid Fermentation of Black Raspberry Extract. Microorganisms. 2021; 9(7):1364. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071364
Chicago/Turabian StyleSong, Young-Ran, Chan-Mi Lee, Seon-Hye Lee, and Sang-Ho Baik. 2021. "Evaluation of Probiotic Properties of Pediococcus acidilactici M76 Producing Functional Exopolysaccharides and Its Lactic Acid Fermentation of Black Raspberry Extract" Microorganisms 9, no. 7: 1364. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms9071364