
Assessment of Fish Abundance, Biodiversity and Movement Periodicity Changes in a Large River over a 20-Year Period

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Fishway Monitoring and Environmental Factors
2.3. Data and Statistical Analysis
3. Results
3.1. Fluctuations in Environmental Factors
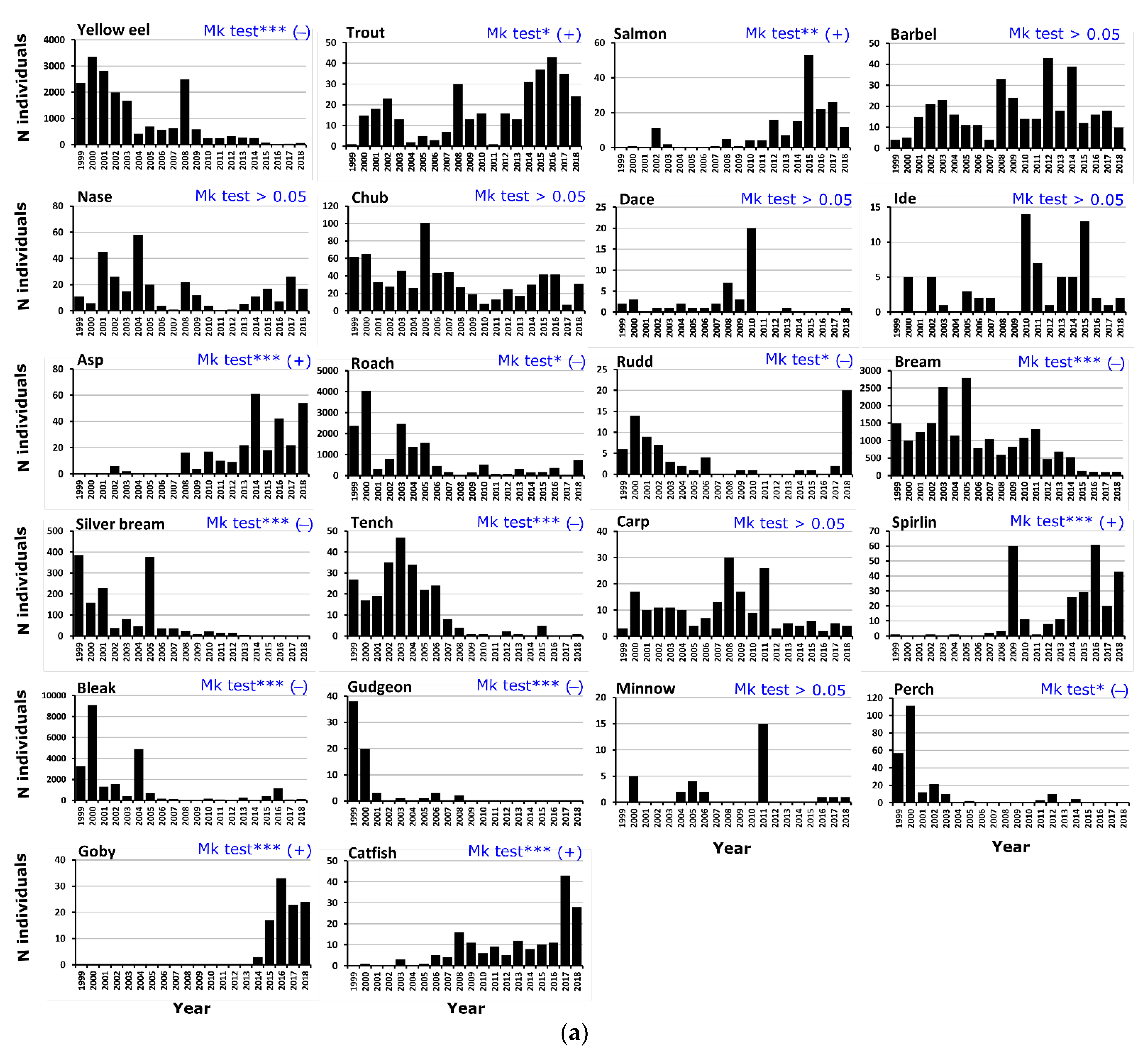
3.2. Overall Biodiversity and Change over Time
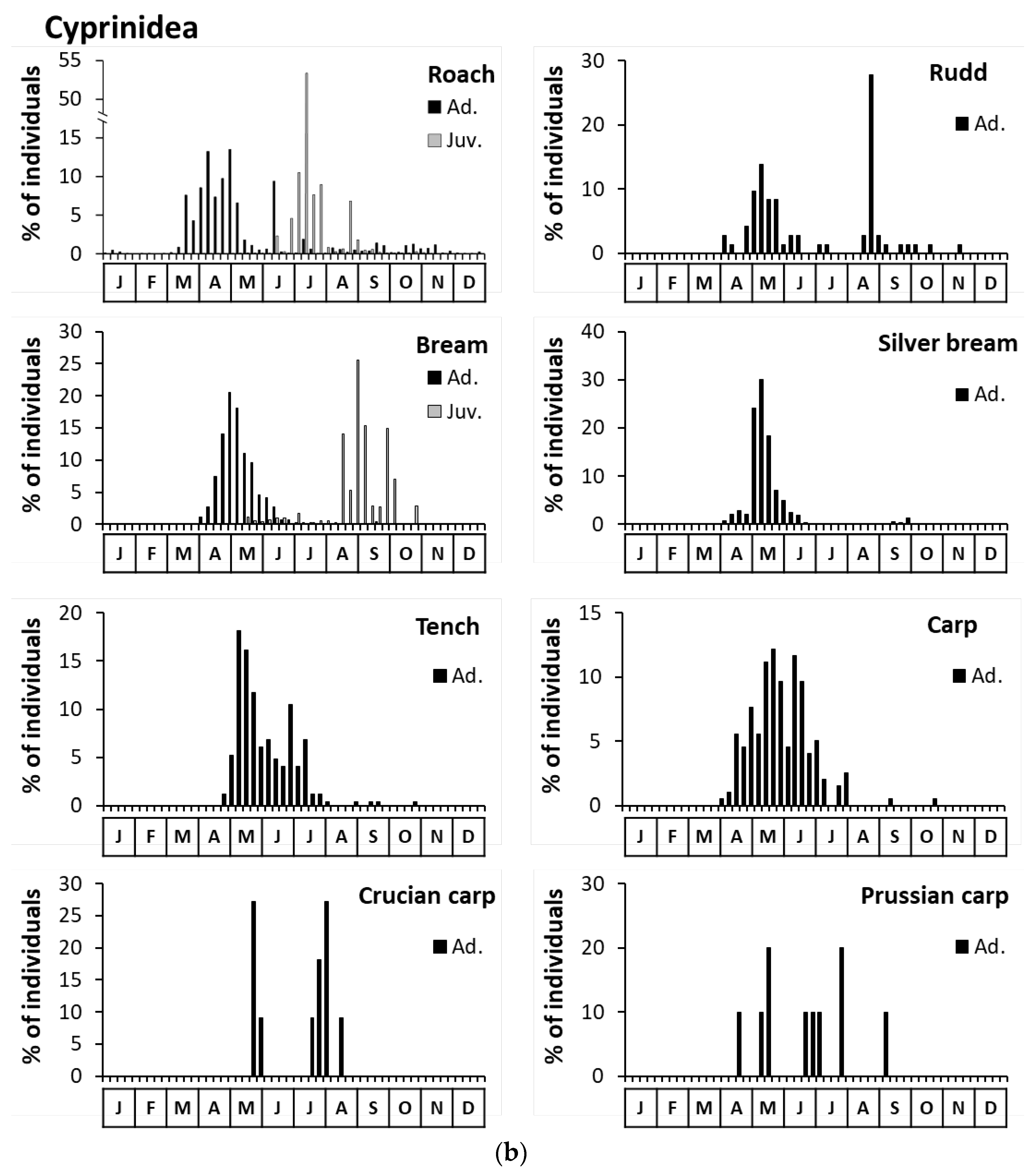
3.3. Overall Fish Movement Indicators and Their Changes over Time
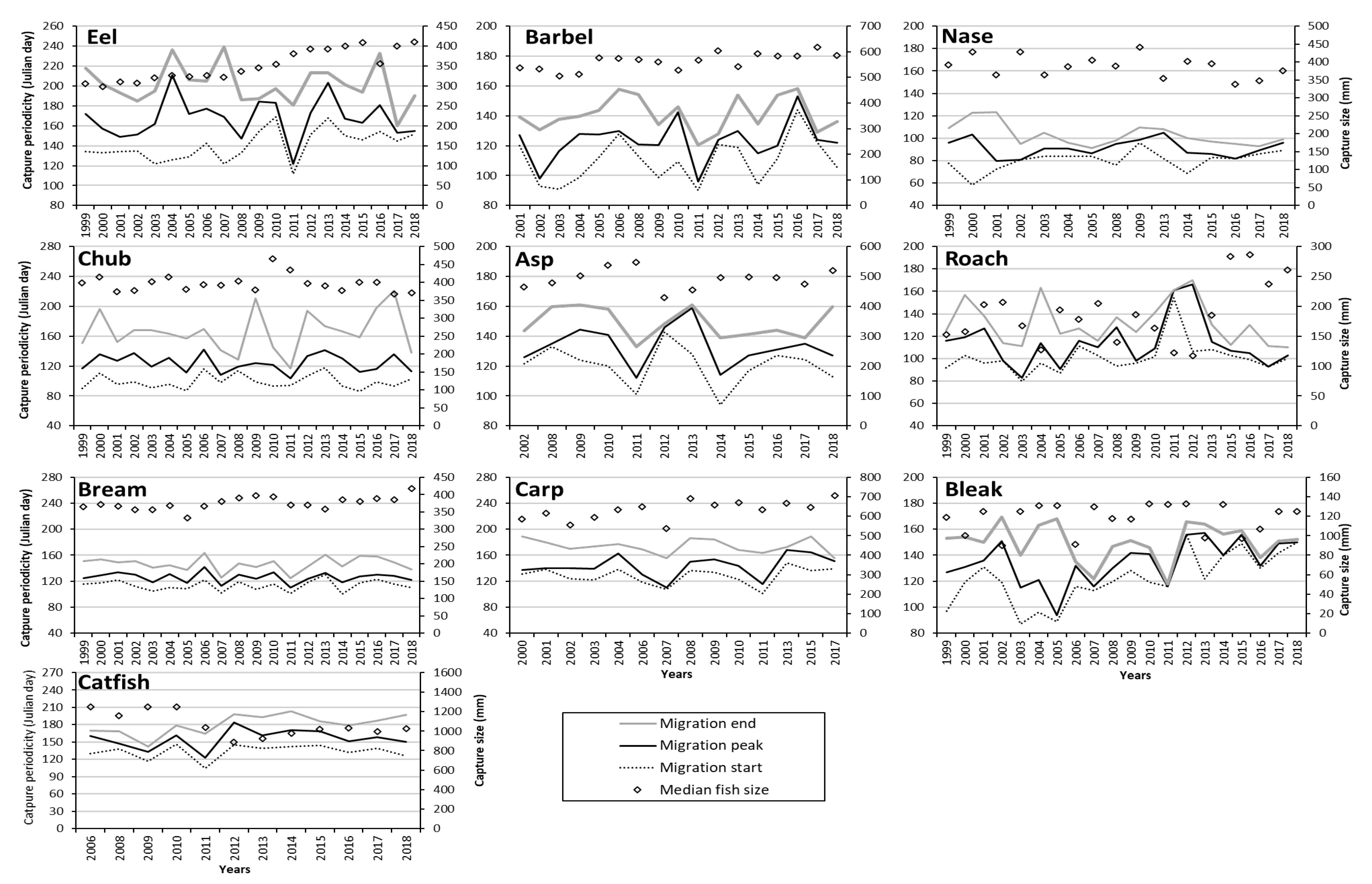
3.3.1. Annual Capture Periodicities by Species
3.3.2. Fish Size and Environmental Factors Associated with Captures
3.3.3. Temporal Trends in Fish Movement Indicators and Fish Size
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nilsson, C.; Reidy, C.A.; Dynesius, M.; Revenga, C. Fragmentation and flow regulation of the world’s large river systems. Science 2005, 308, 405–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capra, H.; Plichard, L.; Bergé, J.; Pella, H.; Ovidio, M.; McNeil, E.; Lamouroux, N. Fish habitat selection in a large hydropeaking river: Strong individual and temporal variations revealed by telemetry. Sci. Total Environ. 2017, 578, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Petts, G.E.; Gurnell, A.M. Dams and geomorphology: Research progress and future directions. Geomorphology 2005, 71, 27–47. [Google Scholar] [CrossRef]
- Silva, A.T.; Lucas, M.C.; Castro-Santos, T.; Katopodis, C.; Baumgartner, L.; Thiem, J.; Aarestrup, K.; Pompeu, P.S.; O’Brien, G.; Braun, D.C.; et al. The future of fish passage science, engineering, and practice. Fish Fish. 2018, 19, 340–362. [Google Scholar] [CrossRef] [Green Version]
- Benitez, J.-P.; Nzau Matondo, B.; Dierckx, A.; Ovidio, M. An overview of potamodromous fish upstream movements in medium-sized rivers, by means of fish passes monitoring. Aquat. Ecol. 2015, 49, 481–497. [Google Scholar] [CrossRef]
- Lucas, M.C.; Baras, E. Migration of Freshwater Fishes; Blackwell Science: Oxford, UK, 2001. [Google Scholar]
- Pringle, C.M. Hydrologic connectivity and the management of biological reserves: A global perspective. Ecol. Appl. 2001, 11, 981–998. [Google Scholar] [CrossRef]
- Birnie-Gauvin, K.; Nielsen, J.; Frandsen, S.B.; Olsen, H.-M.; Aarestrup, K. Catchment-scale effects of river fragmentation: A case study on restoring connectivity. J. Environ. Manag. 2020, 264, 110408. [Google Scholar] [CrossRef]
- Ovidio, M.; Sonny, D.; Watthez, Q.; Goffaux, D.; Detrait, O.; Orban; Nzau Matondo, B.; Renardy, S.; Dierckx, A.; Benitez, J.-P. Evaluation of the performance of successive multispecies improved fishways to reconnect a rehabilitated river. Wetl. Ecol. Manag. 2020, 28, 641–654. [Google Scholar] [CrossRef]
- Gardner, C.J.; Deeming, D.C.; Eady, P.E. Seasonal movements with shifts in lateral and longitudinal habitat use by common bream, Abramis brama, in a heavily modified lowland river. Fish. Manag. Ecol. 2013, 20, 315–325. [Google Scholar] [CrossRef]
- Benitez, J.-P.; Dierckx, A.; Nzau Matondo, B.; Rollin, X.; Ovidio, M. Movement behaviours of potamodromous fish within a large anthropised river after the reestablishment of the longitudinal connectivity. Fish. Res. 2018, 207, 140–149. [Google Scholar] [CrossRef]
- Roscoe, D.W.; Hinch, S.G. Effectiveness monitoring of fish passage facilities: Historical trends, geographic patterns and future directions. Fish Fish. 2010, 11, 12–33. [Google Scholar] [CrossRef]
- Dudgeon, D.; Smith, R.E.W. Exotic species, fisheries and conservation of freshwater biodiversity in tropical Asia: The case of the Sepik River, Papua New Guinea. Aquat. Conserv. 2006, 16, 203–215. [Google Scholar] [CrossRef]
- Poulet, N.; Beaulaton, L.; Dembski, S. Time trends in fish populations in metropolitan France: Insights from national monitoring data. J. Fish Biol. 2011, 79, 1436–1452. [Google Scholar] [CrossRef] [PubMed]
- Kruk, A.; Ciepłucha, M.; Zięba, G.; Tybulczuk, S.; Tszydel, M.; Marszał, L.; Błońska, D.; Galicka, W.; Przybylski, M. Recovery of fish fauna in the upper Warta River, Poland: Long-term (1963–2012) monitoring study. Ecol. Inform. 2016, 33, 109–118. [Google Scholar] [CrossRef]
- Belliard, J.; Beslagic, S.; Delaigue, O.; Tales, E. Reconstructing long-term trajectories of fish assemblages using historical data: The Seine River basin (France) during the last two centuries. Environ. Sci. Pollut. Res. 2018, 25, 23430–23450. [Google Scholar] [CrossRef] [PubMed]
- Kuczynski, L.; Chevalier, M.; Laffaille, P.; Legrand, M.; Grenouillet, G. Indirect effect of temperature on fish population abundances through phenological changes. PLoS ONE 2017, 12, e0175735. [Google Scholar] [CrossRef]
- Lamouroux, N.; Olivier, J.-M.; Capra, H.; Zylberblat, M.; Chandesris, A.; Roger, P. Fish community changes after minimum flow increase: Testing quantitative predictions in the Rhône River at Pierre-Bénite, France. Freshw. Biol. 2006, 51, 1730–1743. [Google Scholar] [CrossRef]
- Latli, A.; Descy, J.-P.; Mondy, C.P.; Floury, M.; Viroux, L.; Otjacques, W.; Marescaux, J.; Depiereux, E.; Ovidio, M.; Usseglio-Polatera, P.; et al. Long-term trends in trait structure of riverine communities facing predation risk increase and trophic resource decline. Ecol. Appl. 2017, 27, 2458–2474. [Google Scholar] [CrossRef]
- Daufresne, M.; Veslot, J.; Capra, H.; Carrel, G.; Poirel, A.; Olivier, J.-M.; Lamouroux, N. Fish community dynamics (1985–2010) in multiple reaches of a large river subjected to flow restoration and other environmental changes. Freshw. Biol. 2015, 60, 1176–1191. [Google Scholar] [CrossRef]
- Gido, K.B.; Dodds, W.K.; Eberle, M.E. Retrospective analysis of fish community change during a half-century of landuse and streamflow changes. J. N. Am. Benthol. Soc. 2010, 29, 970–987. [Google Scholar] [CrossRef]
- Fredrich, F. Long-term investigations of migratory behaviour of asp (Aspius aspius L.) in the middle part of the Elbe River, Germany. J. Appl. Ichthyol. 2003, 19, 294–302. [Google Scholar] [CrossRef]
- Lucas, M.C.; Batley, E. Seasonal movements and behaviour of adult barbel Barbus barbus, a riverine cyprinid fish: Implications for river management. J. Appl. Ecol. 1996, 33, 1345–1358. [Google Scholar] [CrossRef]
- Ovidio, M.; Baras, E.; Goffaux, D.; Birtles, C.; Philippart, J.C. Environmental unpredictability rules the autumn migration of brown trout (Salmo trutta L.) in the Belgian ardennes. Hydrobiologia 1998, 371–372, 263–274. [Google Scholar] [CrossRef] [Green Version]
- Ovidio, M.; Parkinson, D.; Philippart, J.-C.; Baras, E. Multiyear homing and fidelity to residence areas by individual barbel (Barbus barbus). Belg. J. Zool. 2007, 137, 183–190. [Google Scholar]
- Lennox, R.J.; Aarestrup, K.; Cooke, S.J.; Cowley, P.D.; Deng, Z.D.; Fisk, A.T.; Harcourt, R.G.; Heupel, M.; Hinch, S.G.; Holland, K.N.; et al. Envisioning the Future of Aquatic Animal Tracking: Technology, Science, and Application. BioScience 2017, 67, 884–896. [Google Scholar] [CrossRef]
- Bergé, J.; Capra, H.; Pella, H.; Steig, T.; Ovidio, M.; Bultel, E.; Lamouroux, N. Probability of detection and positioning error of a hydro acoustic telemetry system in a fast-flowing river: Intrinsic and environmental determinants. Fish. Res. 2012, 125–126, 1–13. [Google Scholar] [CrossRef]
- Brevé, N.W.P.; Buijse, A.D.; Kroes, M.J.; Wanningen, H.; Vriese, F.T. Supporting decision-making for improving longitudinal connectivity for diadromous and potamodromous fishes in complex catchments. Sci. Total Environ. 2014, 496, 206–218. [Google Scholar] [CrossRef]
- Capra, H.; Pella, H.; Ovidio, M. Individual movements, home ranges and habitat use by native rheophilic cyprinids and non-native catfish in a large regulated river. Fish. Manag. Ecol. 2018, 25, 136–149. [Google Scholar] [CrossRef] [Green Version]
- Poulet, N.; Reyjol, Y.; Collier, H.; Lek, S. Does fish scale morphology allow the identification of populations at a local scale? A case study for rostrum dace Leuciscus leuciscus burdigalensis in River Viaur (SW France). Aquat. Sci. 2005, 67, 122–127. [Google Scholar] [CrossRef]
- Radinger, J.; Wolter, C. Patterns and predictors of fish dispersal in rivers. Fish Fish. 2014, 15, 456–473. [Google Scholar] [CrossRef]
- Bao, J.; Li, W.; Zhang, C.; Mi, X.; Li, H.; Zhao, X.; Cao, N.; Twardek, W.M.; Cooke, S.J.; Duan, M. Quantitative assessment of fish passage efficiency at a vertical-slot fishway on the Daduhe River in Southwest China. Ecol. Eng. 2019, 141, 105597. [Google Scholar] [CrossRef]
- Benitez, J.-P.; Ovidio, M. The influence of environmental factors on the upstream movements of rheophilic cyprinids according to their position in a river basin. Ecol. Freshw. Fish 2018, 27, 660–671. [Google Scholar] [CrossRef]
- Rasmussen, J.E.; Belk, M.C. Individual Movement of Stream Fishes: Linking Ecological Drivers with Evolutionary Processes. Rev. Fish. Sci. Aquac. 2017, 25, 70–83. [Google Scholar] [CrossRef]
- Huet, M. Aperçu des relations entre la pente et les populations piscicoles des eaux courantes. Schweiz. Z. Hydrol. 1949, 11, 332–351. [Google Scholar] [CrossRef]
- Schiemer, F.; Spindler, T. Endangered fish species of the Danube river in Austria. River Res. Appl. 1989, 4, 397–407. [Google Scholar] [CrossRef]
- Philippart, J.C.; Vrancken, M. Atlas des poissons de Wallonie. Distribution, écologie, éthologie, pêche, conservation. Cah. D’éthologie Appliquée 1983, 3, 1–395. [Google Scholar]
- Mann, H.B. Nonparametric tests against trend. Econometrica 1945, 13, 245–259. [Google Scholar] [CrossRef]
- Kendall, M.G. Rank Correlation Methods; Griffin: London, UK, 1975. [Google Scholar]
- Zajicek, P.; Wolter, C. The gain of additional sampling methods for the fish-based assessment of large rivers. Fish. Res. 2018, 197, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Clark, S.J.; Jackson, J.R.; Lochmann, S.E. A comparison of shoreline seines with fyke nets for sampling littoral fish communities in floodplain lakes. N. Am. J. Fish. Manag. 2007, 27, 676–680. [Google Scholar] [CrossRef]
- Legrand, M.; Briand, C.; Buisson, L.; Artur, G.; Azam, D.; Baisez, A.; Barracou, D.; Bourré, N.; Carry, L.; Caudal, A.-L.; et al. Contrasting trends between species and catchments in diadromous fish counts over the last 30 years in France. Knowl. Manag. Aquat. Ecosyst. 2020, 421, 7. [Google Scholar] [CrossRef] [Green Version]
- Morris, E.K.; Caruso, T.; Buscot, F.; Fischer, M.; Hancock, C.; Maier, T.S.; Meiners, T.; Müller, C.; Obermaier, E.; Prati, D.; et al. Choosing and using diversity indices: Insights for ecological applications from the German Biodiversity Exploratories. Ecol. Evol. 2014, 4, 3514–3524. [Google Scholar] [CrossRef] [Green Version]
- Blondel, J. Guilds or functional groups: Does it matter? Oikos 2003, 100, 223–231. [Google Scholar] [CrossRef]
- Wolter, C.; Freyhof, J. Diel distribution patterns of fishes in a temperate large lowland river. J. Fish Biol. 2004, 64, 632–642. [Google Scholar] [CrossRef]
- Ovidio, M.; Sonny, D.; Dierckx, A.; Watthez, Q.; Bourguignon, S.; de le Court, B.; Detrait, O.; Benitez, J.P. The use of behavioural metrics to evaluate fishway efficiency. River Res. Appl. 2017, 33, 1484–1493. [Google Scholar] [CrossRef]
- Stuart, I.G.; Berghuis, A.P. Upstream passage of fish through a vertical-slot fishway in an Australian subtropical river. Fish. Manag. Ecol. 2002, 9, 111–122. [Google Scholar] [CrossRef]
- Nzau Matondo, B.; Ovidio, M. Dynamics of upstream movements of the European eel Anguilla anguilla in an inland area of the River Meuse over the last 20 years. Environ. Biol. Fishes 2016, 99, 223–235. [Google Scholar] [CrossRef] [Green Version]
- Descy, J.-P.; Leitao, M.; Everbecq, E.; Smitz, J.S.; Delige, J.-F. Phytoplankton of the river Loire, France: A biodiversity and modelling study. J. Plankton Res. 2012, 34, 120–135. [Google Scholar] [CrossRef] [Green Version]
- Pigneur, L.-M.; Falisse, E.; Roland, K.; Everbecq, E.; Deliège, J.-F.; Smitz, J.S.; Van Doninck, K.; Descy, J.-P. Impact of invasive Asian clams, Corbicula spp., on a large river ecosystem. Freshw. Biol. 2014, 59, 573–583. [Google Scholar] [CrossRef]
- Otjacques, W.; Latli, A.; Bernard, B.; Ovidio, M.; Depiereux, E.; Kestemont, P. Recent decline of roach Rutilus rutilus stock in a large river ecosystem in relation with its population dynamics. Fundam. Appl. Limnol. 2015, 187, 151–163. [Google Scholar] [CrossRef]
- Otjacques, W.; De Laender, F.; Kestemont, P. Discerning the causes of a decline in a common European fish, the roach (Rutilus rutilus L.): A modelling approach. Ecol. Model. 2016, 322, 92–100. [Google Scholar] [CrossRef]
- Guillerault, N.; Delmotte, S.; Boulêtreau, S.; Lauzeral, C.; Poulet, N.; Santoul, N. Does the non-native European catfish Silurus glanis threaten French river fish populations? Freshw. Biol. 2015, 60, 922–928. [Google Scholar] [CrossRef] [Green Version]
- Cucherousset, J.; Horký, P.; Slavík, O.; Ovidio, M.; Arlinghaus, R.; Boulêtreau, S.; Britton, R.; García-Berthou, E.; Santoul, F. Ecology, behaviour and management of the European catfish. Rev. Fish Biol. Fish. 2018, 28, 177–190. [Google Scholar] [CrossRef] [Green Version]
- Pfauserová, N.; Slavík, O.; Horký, P.; Kolářjová, J.; Randák, T. Migration of non-native predator Asp (Leuciscus aspius) from a reservoir poses a potential threat to native species in tributaries. Water 2019, 11, 1306. [Google Scholar] [CrossRef] [Green Version]
- Boulêtreau, S.; Gaillagot, A.; Carry, L.; Tétard, S.; De Oliveira, E.; Santoul, F. Adult Atlantic salmon have a new freshwater predator. PLoS ONE 2018, 13, e0196046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prignon, C.; Micha, J.C.; Rimbaud, G.; Philippart, J.C. Rehabilitation efforts for Atlantic salmon in the Meuse basin area: Synthesis 1983–1998. Hydrobiologia 1999, 410, 69–77. [Google Scholar] [CrossRef]
- Ovidio, M.; Dierckx, A.; Bunel, S.; Grandry, L.; Spronck, C.; Benitez, J.P. Poor Performance of a Retrofitted Downstream Bypass Revealed by the Analysis of Approaching Behaviour in Combination with a Trapping System. River Res. Appl. 2017, 33, 27–36. [Google Scholar] [CrossRef]
- Ovidio, M.; Renardy, S.; Dierckx, A.; Nzau Matondo, B.; Benitez, J.-P. Improving bypass performance and passage success of Atlantic salmon smolts at an old fish-hostile hydroelectric power station: A challenging task. Ecol. Eng. 2021, 160, 106148. [Google Scholar] [CrossRef]
- Teichert, N.; Benitez, J.; Dierckx, A.; Tétard, S.; De Oliveira, E.; Trancart, T.; Feunteun, E.; Ovidio, M. Development of an accurate model to predict the phenology of Atlantic salmon smolt spring migration. Aquat. Conserv. 2020, 30, 1552–1565. [Google Scholar] [CrossRef]
- Jurajda, P.; Černý, J.; Polačik, M.; Valová, Z.; Janáč, M.; Blažek, R.; Ondračková, M. The recent distribution and abundance of non-native Neogobius fishes in the Slovak section of the River Danube. J. Appl. Ichthyol. 2005, 21, 319–323. [Google Scholar] [CrossRef]
- Kerr, J.R.; Vowles, A.S.; Crabb, M.C.; Kemp, P.S. Selective fish passage: Restoring habitat connectivity without facilitating the spread of a non-native species. J. Environ. Manag. 2021, 279, 110908. [Google Scholar] [CrossRef]
- Baudoin, J.M.; Bergun, V.; Chanseau, M.; Larinier, M.; Ovidio, M.; Sremski, W.; Steinbach, P.; Voegtle, B. Assessing the Passage of Obstacles by Fish. Concepts, Design and Application; Onema: Paris, France, 2015. [Google Scholar]
- Reichard, M.; Jurajda, P.; Ondračková, M. The effect of light intensity on the drift of young-of-the-year cyprinid fishes. J. Fish Biol. 2002, 61, 1063–1066. [Google Scholar] [CrossRef]
- Nzau Matondo, B.; Ovidio, M. Decreased stock entering the Belgian Meuse is associated with the loss of colonisation behaviour in yellow-phase European eels. Aquat. Living Resour. 2018, 31, 7. [Google Scholar] [CrossRef]
- Brönmark, C.; Skov, C.; Brodersen, J.; Nilsson, P.A.; Hansson, L.A. Seasonal migration determined by a trade-off between predator avoidance and growth. PLoS ONE 2008, 3, e1957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, J.M.; Ferreira, M.T. Use of aquatic biota to detect ecological changes in freshwater: Current status and future directions. Water 2020, 12, 1611. [Google Scholar] [CrossRef]
- Santos, R.; Poulet, N.; Besnard, A. Life-history traits correlate with temporal trends in freshwater fish populations for common European species. Freshw. Biol. 2021, 66, 317–331. [Google Scholar] [CrossRef]
- Nzau Matondo, B.; Benitez, J.-P.; Dierckx, A.; Rollin, X.; Ovidio, M. An evaluation of restocking practice and demographic stock assessment methods for cryptic juvenile european eel in upland rivers. Sustainability 2020, 12, 1124. [Google Scholar] [CrossRef] [Green Version]
- Mouton, A.M.; Buysse, D.; Stevens, M.; van den Neucker, T.; Coeck, J. Evaluation of riparian habitat restoration in a lowland river. River Res. Appl. 2012, 28, 845–857. [Google Scholar] [CrossRef]
- van Leeuwen, C.H.A.; Museth, J.; Sandlund, O.T.; Qvenild, T.; Vøllestad, L.A. Mismatch between fishway operation and timing of fish movements: A risk for cascading effects in partial migration systems. Ecol. Evol. 2016, 6, 2414–2425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamanaka, H.; Minamoto, T. The use of environmental DNA of fishes as an efficient method of determining habitat connectivity. Ecol. Ind. 2016, 62, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Deiner, K.; Fronhofer, E.A.; Mächler, E.; Walser, J.-C.; Altermatt, F. Environmental DNA reveals that rivers are conveyer belts of biodiversity information. Nat. Commun. 2016, 7, 12544. [Google Scholar] [CrossRef] [Green Version]
- Reid, A.J.; Carlson, A.K.; Creed, I.F.; Eliason, E.J.; Gell, P.A.; Johnson, P.T.J.; Kidd, K.A.; MacCormack, T.J.; Olden, J.D.; Ormerod, S.J.; et al. Emerging threats and persistent conservation challenges for freshwater biodiversity. Biol. Rev. 2019, 94, 849–873. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Fishway F1 | Fishway F2 |
|---|---|---|
| Fishway type | Pool and weir | Alterne vertical slot |
| Construction year | 1980 | 1998 |
| Discharge of fishway (m3/s) | 0.3 | 1 |
| Attraction flow (m3/s) | 0.2 | 2.5 |
| Length (m) | 48 | 165 |
| Pool number | 56 | 26 |
| Pool size, length × width (m) | 1.5 to 2.5 × 1.2 | 4.7 to 9.7 × 2.5 |
| Pool water depth (m) | 1 | 1.5 |
| Height between pools (m) | 0.15 | 0.3 |
| Water depth of slot (m) | 0.3 | 1.3 |
| Slot width (m) | 0.35 | 0.4 |
| Species | Ecol. Group | N Total | N F1 | N F2 | |
|---|---|---|---|---|---|
| Common Name | Latin Name | (N Adult—N Juvenile) | (N Adult—N Juvenile) | (N Adult—N Juvenile) | |
| Anguillidae | |||||
| European eel | Anguilla anguilla | Eury. | 19,163 (19,163–0) | 19,163 (19,163–0) | 0 |
| Cyprinidae | |||||
| Asp † | Aspius aspius | Rheo. | 752 (283–469) | 470 (1–469) | 282 (282–0) |
| Barbel | Barbus barbus | Rheo. | 464 (351–113) | 127 (24–103) | 337 (327–10) |
| Chub | Leuciscus cephalus | Rheo. | 1309 (709–600) | 642 (45–597) | 667 (664–3) |
| Common bleak | Alburnus alburnus | Eury. | 78,342 (23,866–54,476) | 77,358 (22,882–54,476) | 984 (984–0) |
| Common bream | Abramis brama | Eury. | 21,944 (19,504–2440) | 6352 (3921–2431) | 15,592 (15,583–9) |
| Common Carp † | Cyprinus carpio | Eury. | 198 (197–1) | 3 (2–1) | 195 (195–0) |
| Crucian carp | Carassius carassius | Limno. | 11 (11–0) | 6 (6–0) | 5 (5–0) |
| Dace | Leuciscus leuciscus | Rheo. | 77 (45–32) | 76 (44–32) | 1 (1–0) |
| Grass carp † | Ctenopharyngodon idella | 2 (2–0) | 0 | 2 (2–0) | |
| Gudgeon | Gobio gobio | Rheo. | 69 (68–1) | 64 (63–1) | 5 (5–0) |
| Ide | Leuciscus idus | Rheo | 3209 (68–3141) | 3148 (12–3136) | 61 (56v5) |
| Koi † | Cyprinus carpio | Limno. | 6 (6–0) | 0 | 6 (6–0) |
| Minnow | Phoxinus phoxinus | Rheo. | 53 (31–22) | 53 (31–22) | 0 |
| Nase | Chondrostoma nasus | Rheo. | 676 (308–368) | 369 (3–366) | 307 (305v2) |
| Prussian carp † | Carassius gibelio | Limno. | 10 (10–0) | 9 (9–0) | 1 (1–0) |
| Roach | Rutilus rutilus | Eury. | 222,256 (16,136–206,120) | 217,767 (11,700–206,067) | 4489 (4436–53) |
| Rudd | Scardinius erythrophthalmus | Eury. | 73 (72–1) | 52 (51–1) | 21 (21–0) |
| Silver bream | Blicca bjoerkna | Eury. | 1482 (1477–5) | 461 (456–5) | 1021 (1021–0) |
| Spirlin | Alburnoides bipunctatus | Rheo. | 780 (278–502) | 777 (275–502) | 3 (3–0) |
| Tench | Tinca tinca | Limno. | 249 (248–1) | 57 (57–0) | 192 (191–1) |
| Percidae | |||||
| Perch | Perca fluviatilis | Limno. | 36,107 (237–35,870) | 35,985 (192–35,793) | 122 (45–77) |
| Ruffe | Gymnocephalus cernua | Limno. | 5 (4–1) | 5 (4–1) | 0 |
| Zander | Sander lucioperca | Limno. | 4 (3–1) | 2 (2–0) | 2 (1–1) |
| Salmonidae | |||||
| Brook trout † | Salvelinus fontinalis | Rheo. | 3 (3–0) | 2 (2-0) | 1 (1–0) |
| Rainbow trout † | Oncorhynchus mykiss | Rheo. | 14 (14–0) | 7 (7-0) | 7 (7–0) |
| Salmon ‡ | Salmo salar | Rheo. | 180 (180–0) | 1 (1-0) | 179 (179–0) |
| Trout | Salmo trutta | Rheo. | 353 (346–7) | 37 (31-6) | 316 (315–1) |
| Other species | |||||
| Pike | Esox lucius | Limno. | 8 (8–0) | 0 | 8 (8–0) |
| Pumpkinseed † | Lepomis gibbosus | Limno. | 1 (1–0) | 0 | 0 |
| Catfish † | Silurus glanis | Eury. | 173 (173–0) | 0 | 173 (173–0) |
| Tilapia † | Oreochromis niloticus | Limno. | 1 (1–0) | 0 | 0 |
| Round goby † | Neogobius melanostomus | Limno. | 655 (100–555) | 653 (98-555) | 2 (2–0) |
| Kessler’s goby † | Ponticola kessleri | Limno. | 1 (1–0) | 0 | 1 (1–0) |
| Stickleback | Gasterosteus aculeatus | Eury. | 1 (1–0) | 0 | 0 |
| Total | 388,631 (83,905–304,726) | 363,648 (59,084–304,564) | 24,983 (24,821–162) | ||
| Species | Peak Period | N | Capture Size (mm) | Capture Periodicity | Capture Temperature (°C) | Capture Flow (m3/s) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (Capture Date) | ||||||||||||||
| P10 | P50 | P90 | P10 | P50 | P90 | P10 | P50 | P90 | P10 | P50 | P90 | |||
| Yellow eel | Spr + Sum + Aut | 19,163 | 253 | 316 | 405 | 133 | 160 | 202 | 18.3 | 22.1 | 24.8 | 49.3 | 121.2 | 219.7 |
| Trout | Spr + Sum | 226 | 371 | 500 | 648 | 137 | 164 | 198 | 17.5 | 20.1 | 22.7 | 50.1 | 98.3 | 248.4 |
| Aut | 99 | 454 | 562 | 690 | 284 | 316 | 344 | 9.4 | 11.9 | 18.2 | 40.9 | 151.1 | 411.7 | |
| Salmon | Spr | 63 | 667 | 780 | 921 | 119 | 140 | 163 | 14.3 | 18.1 | 21.2 | 44.3 | 95.3 | 222.4 |
| Aut | 95 | 655 | 754 | 900 | 267 | 287 | 326 | 10.3 | 14.8 | 17.0 | 43.6 | 94.2 | 287.6 | |
| Barbel | Spr | 244 | 438 | 568 | 648 | 94 | 124 | 150 | 12.3 | 14.9 | 19.4 | 60.7 | 175.4 | 382.2 |
| Aut | 90 | 312 | 435 | 604 | 273 | 287 | 325 | 12.1 | 15.2 | 18.0 | 30.9 | 98.6 | 304.9 | |
| Nase | Spr | 303 | 335 | 382 | 440 | 78 | 91 | 108 | 9.4 | 11.3 | 13.8 | 130.4 | 236.3 | 829.7 |
| Chub | Spr + Sum | 673 | 327 | 395 | 462 | 90 | 120 | 167 | 13.3 | 16.0 | 2.9 | 80.7 | 165.0 | 366.4 |
| Aut | 17 | 172 | 362 | 483 | 261 | 279 | 325 | 10.2 | 16.9 | 20.1 | 16.0 | 117.3 | 486.6 | |
| Dace | Spr | 23 | 105 | 125 | 173 | 93 | 128 | 157 | 9.3 | 17.9 | 21.3 | 71.1 | 160.2 | 454.4 |
| Aut | 21 | 108 | 120 | 175 | 281 | 291 | 334 | 10.5 | 14.6 | 15.0 | 65.3 | 65.3 | 277.6 | |
| Ide | Spr | 32 | 199 | 414 | 501 | 82 | 97 | 135 | 9.2 | 13.4 | 17.1 | 167.5 | 208.3 | 409.8 |
| Aut | 32 | 256 | 342 | 415 | 287 | 305 | 330 | 11.1 | 14.7 | 18.2 | 23.9 | 40.9 | 165.4 | |
| Asp | Spr | 233 | 436 | 498 | 548 | 108 | 130 | 157 | 15.2 | 17.2 | 21.1 | 55.6 | 102.6 | 214.6 |
| Sum | 48 | 415 | 449 | 525 | 176 | 193 | 221 | 19.8 | 22.0 | 24.1 | 52.4 | 122.2 | 277.7 | |
| Roach | Spr | 13,035 | 114 | 167 | 242 | 87 | 110 | 157 | 12.0 | 15.4 | 20.8 | 120.1 | 187.4 | 306.9 |
| Sum | 984 | 103 | 117 | 192 | 187 | 216 | 255 | 19.2 | 20.7 | 22.9 | 36.6 | 47.6 | 135.4 | |
| Aut | 1191 | 115 | 172 | 290 | 265 | 291 | 316 | 11.6 | 15.0 | 20.3 | 15.7 | 57.7 | 331.1 | |
| Rudd | Spr | 40 | 188 | 275 | 337 | 111 | 125 | 155 | 15.1 | 17.8 | 20.6 | 115.4 | 193.3 | 328.8 |
| Sum | 29 | 110 | 124 | 288 | 229 | 232 | 248 | 19.1 | 22.6 | 23.1 | 32.8 | 35.73 | 86.76 | |
| Bream | Spr | 18,940 | 314 | 366 | 411 | 107 | 123 | 147 | 13.9 | 17.1 | 20.0 | 69.9 | 141.9 | 301 |
| Sum | 464 | 159 | 370 | 419 | 173 | 187 | 260 | 17.8 | 21.0 | 23.6 | 40.8 | 88.2 | 278.2 | |
| Aut | 87 | 154 | 180 | 381 | 263 | 267 | 304 | 14.2 | 19.0 | 20.3 | 49.7 | 68.6 | 174.9 | |
| Silver bream | Spr | 1434 | 132 | 222 | 271 | 115 | 124 | 145 | 14.4 | 17.1 | 18.9 | 130.8 | 174.5 | 346.8 |
| Aut | 30 | 112 | 123 | 159 | 263 | 267 | 291 | 14.9 | 20.1 | 20.4 | 65.3 | 174.9 | 174.9 | |
| Tench | Spr + Sum | 244 | 374 | 420 | 468 | 124 | 142 | 185 | 17.3 | 18.8 | 24.1 | 51.3 | 115.3 | 238.8 |
| Carp | Spr + Sum | 196 | 487 | 641 | 762 | 113 | 145 | 182 | 16.9 | 19.7 | 23.8 | 43.7 | 113.6 | 229.5 |
| Crucian carp | Spr + Sum | 11 | 172 | 278 | 374 | 136 | 202 | 221 | 15.1 | 22.2 | 24.1 | 74.4 | 141.6 | 654.1 |
| Prussian carp | Spr + Sum | 10 | 160 | 245 | 326 | 109 | 174 | 240 | 17.7 | 20.0 | 23.8 | 68.4 | 129.0 | 188.2 |
| Bleak | Spr | 21,327 | 95 | 110 | 136 | 101 | 131 | 156 | 14.1 | 19.0 | 21.4 | 91.5 | 154.5 | 239.3 |
| Sum | 2084 | 90 | 90 | 117 | 183 | 210 | 238 | 20.6 | 22.7 | 23.2 | 41.5 | 52.1 | 136.1 | |
| Aut | 411 | 83 | 90 | 122 | 273 | 274 | 291 | 15.0 | 17.1 | 20.3 | 57.8 | 84.6 | 165.6 | |
| Sprilin | Spr + Sum | 502 | 66 | 70 | 82 | 170 | 197 | 244 | 18.6 | 22.9 | 24.7 | 22.2 | 64.1 | 149.3 |
| Gudgeon | Spr | 57 | 92 | 104 | 132 | 102 | 129 | 151 | 15.1 | 18.9 | 20.7 | 129.2 | 163.2 | 310.9 |
| Perch | Spr | 179 | 174 | 229 | 332 | 118 | 129 | 151 | 15.3 | 18.1 | 20.8 | 95.8 | 171.3 | 241.0 |
| Aut | 22 | 158 | 185 | 317 | 267 | 273 | 318 | 11.0 | 19.6 | 20.2 | 19.5 | 57.7 | 323.9 | |
| Sum | 34 | 152 | 172 | 321 | 187 | 238 | 260 | 17.7 | 22.0 | 24.1 | 36.2 | 82.5 | 200.2 | |
| Catfish | Spr + Sum | 171 | 851 | 1033 | 1340 | 128 | 155 | 189 | 17.1 | 21.3 | 23.8 | 26.7 | 84.5 | 211.1 |
| Goby | Spr | 75 | 61 | 75 | 106 | 130 | 140 | 153 | 16.9 | 19.1 | 21.3 | 52.31 | 108.4 | 558.5 |
| Sum | 23 | 60 | 66 | 115 | 202 | 232 | 256 | 18.7 | 23.1 | 24.1 | 17.8 | 35.7 | 84.9 | |
| Species | Peak Period | N | Capture Size (mm) | Capture Periodicity | Capture Temperature (°C) | Capture Flow (m3/s) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (Day of the Year) | ||||||||||||||
| P10 | P50 | P90 | P10 | P50 | P90 | P10 | P50 | P90 | P10 | P50 | P90 | |||
| Barbel | Spr | 18 | 56 | 74 | 154 | 102 | 145 | 168 | 14.4 | 19.7 | 22.8 | 36.1 | 61.9 | 169.1 |
| Sum | 86 | 48 | 97 | 157 | 181 | 207 | 251 | 18.7 | 22.1 | 24.2 | 34.1 | 78.8 | 257.7 | |
| Nase | Sum | 259 | 30 | 36 | 69 | 168 | 186 | 239 | 19.8 | 22.6 | 25.4 | 22.5 | 76.2 | 217.2 |
| Aut | 89 | 40 | 47 | 61 | 281 | 289 | 298 | 17.3 | 17.5 | 17.9 | 91.5 | 92.1 | 263.1 | |
| Chub | Sum | 557 | 40 | 50 | 69 | 181 | 190 | 232 | 22.3 | 23.6 | 25.1 | 51.5 | 86.4 | 97.3 |
| Dace | Sum | 20 | 47 | 66 | 80 | 176 | 197 | 227 | 21.1 | 23.9 | 24.6 | 23.2 | 50.3 | 96.4 |
| Ide | Sum | 3059 | 60 | 66 | 75 | 181 | 190 | 220 | 21.8 | 22.9 | 24.3 | 35.5 | 56.5 | 78.3 |
| Asp | Sum | 467 | 74 | 78 | 88 | 181 | 185 | 211 | 22.0 | 22.9 | 23.9 | 21.5 | 51.1 | 61.3 |
| Roach | Spr | 6011 | 49 | 49 | 84 | 140 | 161 | 162 | 20.2 | 22.2 | 22.2 | 25.4 | 25.4 | 147.0 |
| Sum | 198,460 | 41 | 49 | 60 | 181 | 190 | 228 | 21.8 | 23.6 | 25.1 | 40.0 | 70.0 | 97.3 | |
| Aut | 1649 | 51 | 57 | 88 | 268 | 289 | 302 | 13.3 | 17.7 | 19.5 | 44.2 | 109.9 | 193.6 | |
| Bream | Spr | 118 | 69 | 105 | 124 | 133 | 149 | 165 | 14.2 | 20.7 | 22.1 | 69.5 | 161.6 | 498.5 |
| Sum | 1723 | 50 | 60 | 70 | 224 | 237 | 244 | 20.7 | 21.8 | 23.0 | 20.2 | 38.5 | 71.2 | |
| Aut | 599 | 65 | 65 | 97 | 265 | 265 | 291 | 15.3 | 19.4 | 19.4 | 18.2 | 28.9 | 65.3 | |
| Bleak | Spr | 1664 | 50 | 70 | 78 | 135 | 160 | 171 | 19.2 | 22.1 | 24.5 | 53.0 | 99.5 | 156.5 |
| Sum | 50,299 | 50 | 51 | 60 | 207 | 221 | 240 | 21.9 | 24.2 | 25.9 | 51.5 | 82.0 | 84.9 | |
| Aut | 2513 | 58 | 67 | 72 | 266 | 269 | 287 | 16.0 | 18.2 | 21.3 | 33.0 | 64.9 | 165.6 | |
| Spirlin | Spr + Sum | 502 | 47 | 55 | 63 | 149 | 168 | 196 | 19.2 | 21.1 | 23.5 | 45.7 | 84.3 | 137.2 |
| Minnow | Spr | 22 | 38 | 42 | 44 | 129 | 129 | 155 | 19.6 | 19.6 | 21.7 | 27.3 | 47.0 | 145.9 |
| Perch | Spr | 92 | 43 | 102 | 127 | 142 | 164 | 171 | 16.4 | 20.6 | 22.7 | 30.3 | 121.0 | 163.0 |
| Sum | 35,665 | 68 | 73 | 76 | 197 | 203 | 223 | 19.3 | 23.0 | 23.7 | 50.8 | 64.9 | 96.4 | |
| Aut | 92 | 92 | 101 | 121 | 275 | 291 | 291 | 15.3 | 15.3 | 17.8 | 61.4 | 65.3 | 102.6 | |
| Goby | Spr | 301 | 38 | 47 | 56 | 139 | 142 | 156 | 17.6 | 19.2 | 22.5 | 52.3 | 58.9 | 97.3 |
| Sum | 254 | 27 | 36 | 52 | 186 | 229 | 245 | 21.3 | 23.9 | 25.4 | 17.0 | 48.0 | 74.6 | |
| Species | N Years | Indicators | Temporal Trend | Headcount | Mean Spring Temperature | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Tau | p-Value | t-Value | p-Value | R² | t-Value | p-Value | R² | |||
| Eel | 20 | Migration start | 0.34 | 0.04 * | −1.84 | 0.08 | 0.16 | −3.82 | 0.001 ** | 0.45 |
| Migration peak | 0.03 | 0.87 | −1.45 | 0.16 | 0.10 | −2.54 | 0.02 * | 0.26 | ||
| Migration end | −0.12 | 0.47 | −0.40 | 0.70 | 0.001 | −0.70 | 0.49 | 0.03 | ||
| Fish size | 0.86 | <0.001 *** | −4.69 | <0.001 *** | 0.55 | −0.53 | 0.61 | 0.02 | ||
| Barbel | 16 | Migration start | 0.19 | 0.30 | −0.37 | 0.72 | 0.01 | −3.98 | 0.001 ** | 0.51 |
| Migration peak | 0.07 | 0.71 | −1.06 | 0.31 | 0.07 | −2.95 | 0.001 ** | 0.37 | ||
| Migration end | 0.05 | 0.80 | −1.54 | 0.15 | 0.14 | −4.42 | <0.001 *** | 0.56 | ||
| Fish size | 0.48 | 0.01 * | 1.68 | 0.11 | 0.16 | −0.06 | 0.95 | 0.00 | ||
| Nase | 15 | Migration start | 0.40 | 0.04 * | 0.35 | 0.73 | 0.01 | −0.25 | 0.80 | 0.00 |
| Migration peak | 0.01 | 1.00 | −1.64 | 0.12 | 0.17 | −0.28 | 0.78 | 0.01 | ||
| Migration end | −0.32 | 0.10 | −0.32 | 0.75 | 0.01 | −0.19 | 0.85 | 0.00 | ||
| Fish size | −0.28 | 0.17 | −0.32 | 0.75 | 0.01 | 1.50 | 0.16 | 0.15 | ||
| Chub | 20 | Migration start | 0.04 | 0.39 | −0.98 | 0.34 | 0.05 | −2.66 | 0.02 * | 0.28 |
| Migration peak | −0.09 | 0.60 | −1.19 | 0.25 | 0.07 | −2.46 | 0.02 * | 0.25 | ||
| Migration end | 0.06 | 0.69 | −0.37 | 0.72 | 0.01 | −1.16 | 0.26 | 0.07 | ||
| Fish size | −0.14 | 0.39 | 0.40 | 0.69 | 0.01 | 0.40 | 0.69 | 0.01 | ||
| Asp | 12 | Migration start | −0.18 | 0.45 | −2.67 | 0.02 * | 0.41 | −2.47 | 0.03 * | 0.38 |
| Migration peak | 0.00 | 1 | −1.54 | 0.15 | 0.19 | −3.89 | 0.003 ** | 0.60 | ||
| Migration end | −0.12 | 0.62 | −0.37 | 0.72 | 0.01 | −1.93 | 0.08 | 0.27 | ||
| Fish size | 0.10 | 0.73 | 0.88 | 0.4 | 0.07 | 1.99 | 0.07 | 0.28 | ||
| Roach | 19 | Migration start | 0.21 | 0.23 | −1.19 | 0.28 | 0.07 | 0.60 | 0.56 | 0.02 |
| Migration peak | −0.11 | 0.53 | 0.72 | 0.48 | 0.03 | −0.01 | 0.97 | 0.00 | ||
| Migration end | −0.12 | 0.48 | 0.55 | 0.59 | 0.02 | 0.01 | 0.99 | 0.00 | ||
| Fish size | 0.23 | 0.18 | −0.97 | 0.35 | 0.05 | −0.90 | 0.38 | 0.05 | ||
| Bream | 20 | Migration start | −0.02 | 0.92 | −1.67 | 0.11 | 0.13 | −7.41 | <0.001 *** | 0.75 |
| Migration peak | −0.09 | 0.58 | −1.43 | 0.17 | 0.10 | −5.07 | <0.001 *** | 0.59 | ||
| Migration end | −0.06 | 0.74 | −1.52 | 0.14 | 0.11 | −5.33 | <0.001 *** | 0.61 | ||
| Fish size | 0.44 | 0.01 * | −4.50 | <0.001 *** | 0.53 | 0.25 | 0.81 | 0.00 | ||
| Carp | 14 | Migration start | 0.08 | 0.74 | −1.27 | 0.23 | 0.12 | −3.09 | 0.01 * | 0.44 |
| Migration peak | 0.28 | 0.19 | −1.29 | 0.22 | 0.12 | −2.68 | 0.02 * | 0.37 | ||
| Migration end | −0.26 | 0.21 | 0.88 | 0.39 | 0.06 | −1.01 | 0.33 | 0.08 | ||
| Fish size | 0.49 | 0.02 * | 0.04 | 0.97 | 0.00 | −1.79 | 0.10 | 0.21 | ||
| Bleak | 20 | Migration start | 0.52 | 0.002 ** | −1.22 | 0.24 | 0.08 | −1.02 | 0.32 | 0.05 |
| Migration peak | 0.35 | 0.03 * | −0.87 | 0.40 | 0.04 | −2.25 | 0.04 * | 0.22 | ||
| Migration end | −0.07 | 0.67 | 0.75 | 0.46 | 0.03 | −1.59 | 0.13 | 0.12 | ||
| Fish size | 0.11 | 0.51 | −0.70 | 0.49 | 0.03 | 2.60 | 0.02 * | 0.27 | ||
| Catfish | 12 | Migration start | 0 | 1 | 0.02 | 0.98 | 0.00 | −1.71 | 0.12 | 0.23 |
| Migration peak | 0.03 | 0.94 | −0.39 | 0.7 | 0.02 | −1.33 | 0.21 | 0.15 | ||
| Migration end | 0.41 | 0.07 | 0.57 | 0.58 | 0.03 | −0.24 | 0.81 | 0.01 | ||
| Fish size | −0.36 | 0.12 | −0.67 | 0.52 | 0.04 | −0.07 | 0.94 | 0.00 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benitez, J.-P.; Dierckx, A.; Rimbaud, G.; Nzau Matondo, B.; Renardy, S.; Rollin, X.; Gillet, A.; Dumonceau, F.; Poncin, P.; Philippart, J.-C.; et al. Assessment of Fish Abundance, Biodiversity and Movement Periodicity Changes in a Large River over a 20-Year Period. Environments 2022, 9, 22. https://0-doi-org.brum.beds.ac.uk/10.3390/environments9020022
Benitez J-P, Dierckx A, Rimbaud G, Nzau Matondo B, Renardy S, Rollin X, Gillet A, Dumonceau F, Poncin P, Philippart J-C, et al. Assessment of Fish Abundance, Biodiversity and Movement Periodicity Changes in a Large River over a 20-Year Period. Environments. 2022; 9(2):22. https://0-doi-org.brum.beds.ac.uk/10.3390/environments9020022
Chicago/Turabian StyleBenitez, Jean-Philippe, Arnaud Dierckx, Gilles Rimbaud, Billy Nzau Matondo, Séverine Renardy, Xavier Rollin, Alain Gillet, Frédéric Dumonceau, Pascal Poncin, Jean-Claude Philippart, and et al. 2022. "Assessment of Fish Abundance, Biodiversity and Movement Periodicity Changes in a Large River over a 20-Year Period" Environments 9, no. 2: 22. https://0-doi-org.brum.beds.ac.uk/10.3390/environments9020022