
Airborne Fungi in Show Caves from Southern Spain
by
 , , ,
, , ,
Irene Dominguez-Moñino
 ,
,
Valme Jurado
 ,
,
Miguel Angel Rogerio-Candelera
,
Bernardo Hermosin
and
Cesareo Saiz-Jimenez
* Instituto de Recursos Naturales y Agrobiologia, IRNAS-CSIC, 41012 Sevilla, Spain
*
Author to whom correspondence should be addressed.
Appl. Sci. 2021, 11(11), 5027; https://0-doi-org.brum.beds.ac.uk/10.3390/app11115027
Submission received: 26 April 2021
/
Revised: 25 May 2021
/
Accepted: 26 May 2021
/
Published: 29 May 2021
(This article belongs to the Special Issue Fungi Associated with Indoor Environments and Materials)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The aerobiology of caves in Southern Spain possesses special characteristics, different from caves located in Northern Spain. Previous studies demonstrated the influence of outdoor air on caves in the north and the existence of two different patterns, depending on the season. In summer there is an abundance of Ascomycota, whereas in winter Basidiomycota predominates, which are related to the periods of stagnation and ventilation, respectively. In caves in Southern Spain the presence of airborne Basidiomycota is scarce and Ascomycota represents the main group of fungi widely distributed across the caves in all seasons. The most characteristic features were the abundant presence of entomopathogenic fungi (Beauveria bassiana, Parengyodontium album, Pochonia chlamydosporia, Leptobacillium symbioticum, Leptobacillium leptobactrum) and Cladosporium cladosporioides in Cueva del Tesoro, Cueva de Ardales and Gruta de las Maravillas. However, the presence of yeasts of the genera Cutaneotrichosporon, Trichosporon, Cryptococcus, Naganishia, Cystobasidium, Microstroma and Phragmotaenium was exclusive to Gruta de las Maravillas. Fungal hazard in the three show caves were determined using an ecological indicator based on the concentration of spores in cave air.
1. Introduction
The study of the behavior of airborne fungi in caves is of interest because many caves contain Paleolithic paintings that should be protected from fungal outbreaks. In the past, fungal outbreaks were noticed in a number of caves, of which Lascaux and Castañar de Ibor caves were noteworthy [1,2]. In Lascaux Cave, the outbreak was difficult to control using biocides such as different benzalkonium chloride derivatives [3], while in the Castañar de Ibor Cave it was easily controlled by the adoption of an appropriate management (mechanical removal of the polluted sediment and sterilization of the lying materials with hydrogen peroxide) [4].
Caneva et al. [5] recognized that preventive conservation of cultural heritage includes aerobiological analysis in order to assess biological risk and propose strategies aimed at controlling biodeterioration. In the last decade, aerobiology has proved to be a suitable approach for studying the dispersion of airborne fungi in caves [6,7,8,9]. Fungal spores represent a potential risk to the conservation of cultural heritage since, in favorable cave environmental conditions (high relative humidity, stable temperature, nutrient inputs, etc.), fungi colonize speleothems and other mineral substrata in addition to the cadavers of animals and arthropods populating the cavity [2,10].
Airborne fungi are transported from outside the cave to the inside due to the fact that a door opening favors communication with the external atmosphere and the passage of warm air moves towards the interior of the cave. This implies that the rooms and galleries near a cave entrance present a higher concentration of spores than in deeper areas. This was also confirmed by a study of fungal sequences from spores inside and outside a cave, which resulted in being identical [8].
The aerobiology of caves in Southern Spain possesses special characteristics, different from those located in Northern Spain. Previous studies demonstrated the influence of exterior air on the cave and the existence of two different patterns, depending on the season, in La Garma Cave. In summer there was an abundance of Ascomycota, whereas in winter Basidiomycota predominated [11], which are related to periods of stagnation and ventilation, respectively. In Southern Spain the presence of airborne Basidiomycota in caves is scarce and Ascomycota represented the main group of fungi, which was widely distributed across the caves [6,12].
In this study we present aerobiological data on three show caves located in Andalusia, Southern Spain, recorded in four seasons over a year and discuss the origin of airborne fungi.
2. Materials and Methods
2.1. Cave Locations and Research Background
Three show caves located in Andalusia (Southern Spain) were studied: Cueva del Tesoro, Cueva de Ardales and Gruta de las Maravillas. The caves have different geomorphologies, management, and number of visitors. In 2011, the year of this aerobiological study, Cueva de Ardales in Ardales, Malaga, received 4018 visitors/year, Cueva del Tesoro in Rincon de la Victoria, Malaga, received 28,257, and Gruta de las Maravillas in Aracena, Huelva, received 130,314. Cueva del Tesoro and Cueva de Ardales contain Paleolithic paintings, while Gruta de las Maravillas is characterized by a wide variety of beautiful speleothems.
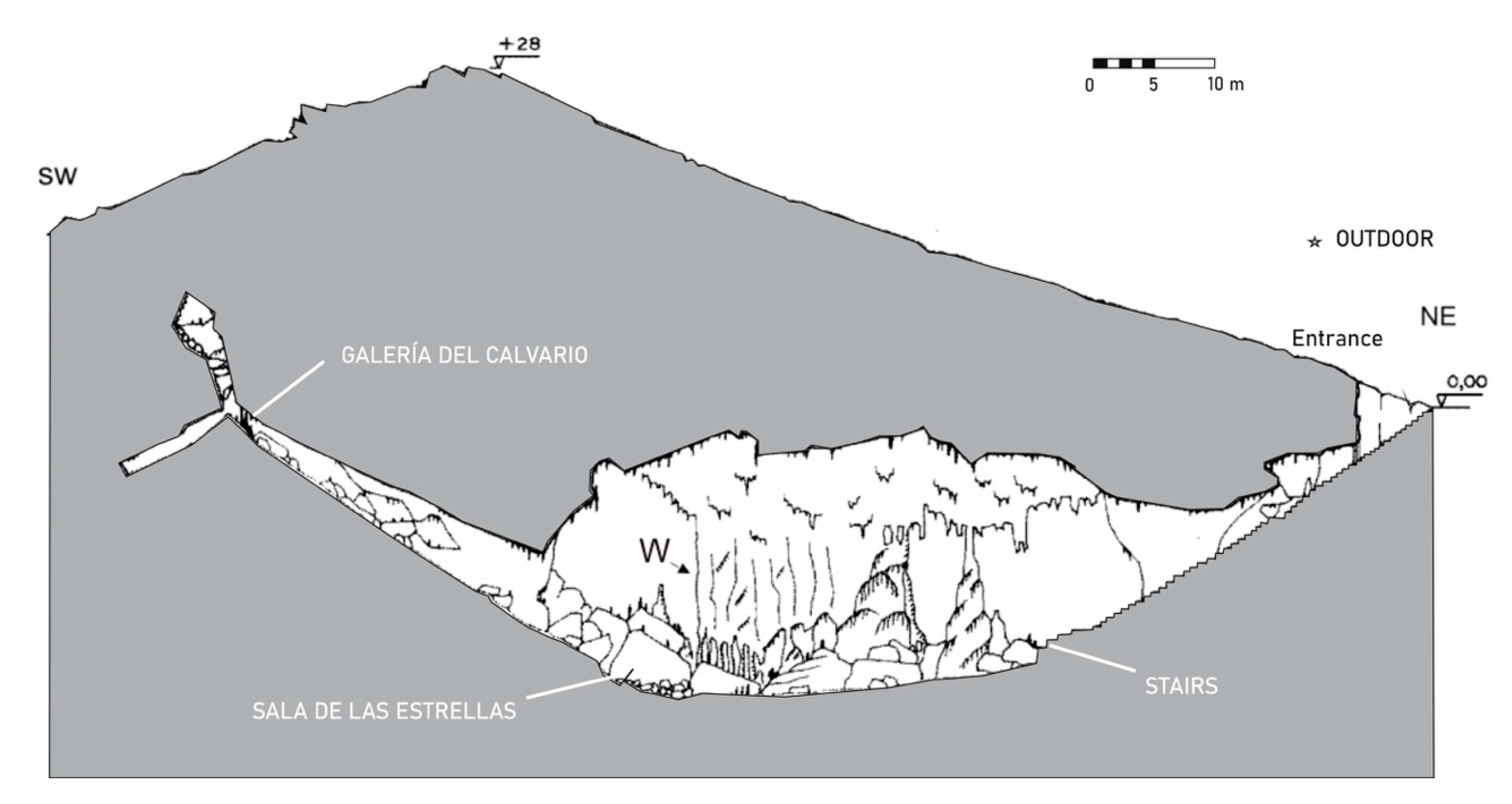
Cueva de Ardales (Figure 1) is located on the northern slopes of Cerro de la Calinoria and the entrance opens at 565 m above sea level. The cavity has a length of 1577 m. The maximum slope is 34.31 m. In a study carried out by Fernandez-Cortes et al. [6] different microclimatic parameters (temperature, concentration of carbon dioxide, and radon) were monitored. Inside the cave, air temperature was recorded in different areas. In Galeria del Calvario the temperature was around 17.45 °C, higher than those registered at the entrance (16.78 °C) and in Sala de las Estrellas (16.55 °C). This warming in Galeria del Calvario coincides with a radon concentration (222Rn) higher in this area (12,000–14,000 Bq/m3) than in the entrance area (6000–9000 Bq/m3).
Galeria del Calvario is the highest topographical area (Figure 1) and closest to the surface, but practically no aerodynamic processes take place between it and the outer atmosphere. Increases in air temperature and carbon dioxide concentrations were recorded in this area as a result of visits; the increase being proportional to the number of visitors. After the visit, there was a flow of carbon dioxide from Galeria del Calvario to Sala de las Estrellas, where the concentration was lower. In Sala de las Estrellas no changes in air temperature were observed due to visits [6].
When the cave temperature is higher than the outdoor temperature, the opening of the door causes a convective air circulation. The entry of exterior air is favored, which is colder and denser, and, simultaneously, the warmer and less dense air leaves the cavity. Therefore, this ventilation gives rise to a change in the levels of temperature, carbon dioxide and 222Rn in the cave atmosphere, mainly in areas with a direct connection to the entrance, such as the entrance stairs and Sala de las Estrellas [6].
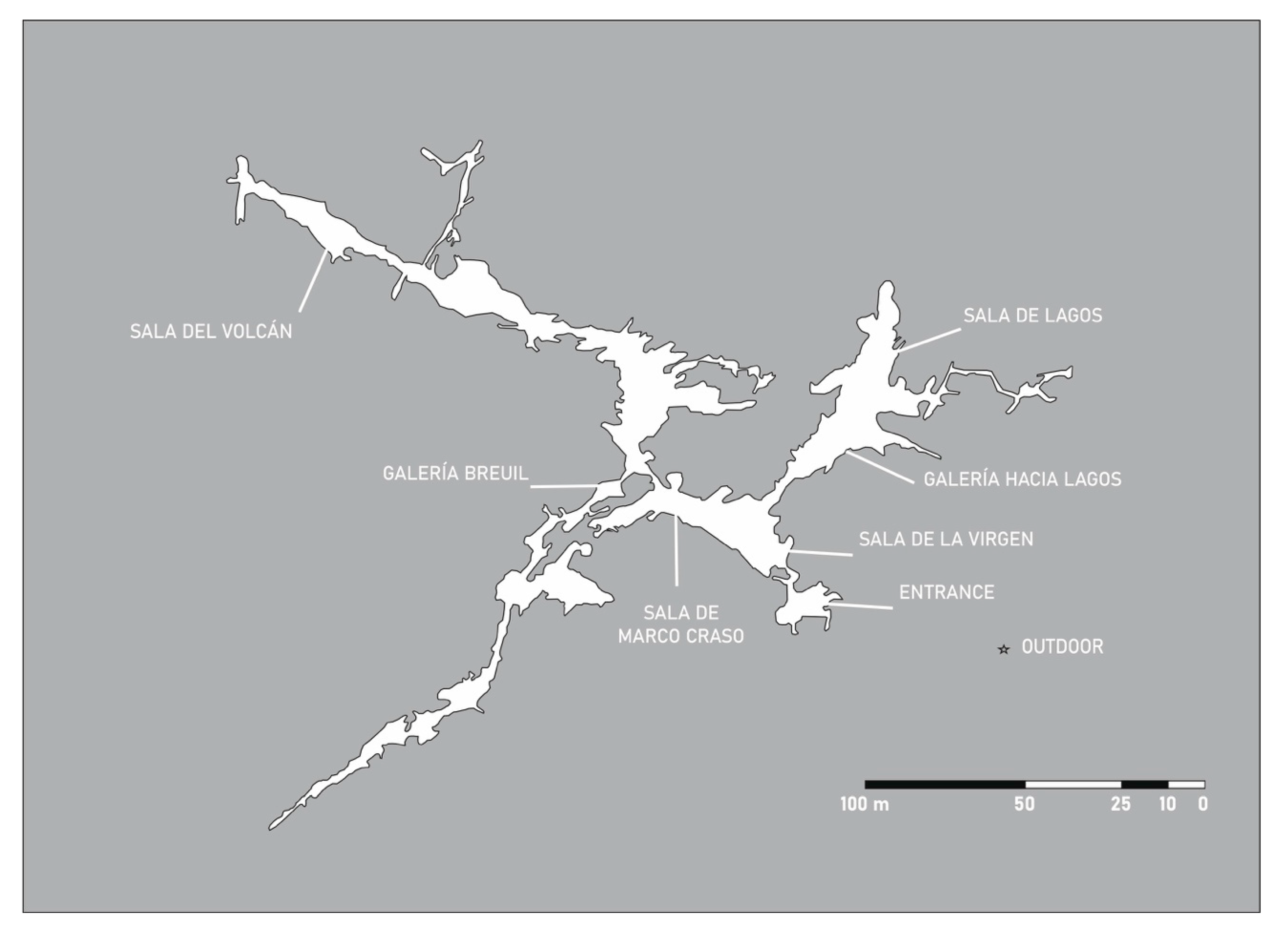
Cueva del Tesoro (Figure 2) is located in a calcareous massif corresponding to the Complejo Malaguide (Precambric–Lower Eocene) and is formed on a thick bed of about 70 m of white limestone (Dogger–Malm). The cave is about 80 m above sea level and the water supply is reduced to water from direct infiltration of rain that falls on the karst surface with a few dripping points and the response time to rain was estimated to be 2–3 h of delay from the start of the rain. This explains why the generation of speleothems is so scarce in the cave.
Measurements of CO2, temperature and relative humidity in the cave demonstrate particular micro-environmental features. In Sala de Lagos and Sala del Volcan (Figure 2) CO2 concentrations were of the order of 200 to 550 ppm, temperature was 14.8–16.8 °C, and the relative humidity was 80 to 90%; normal values for a karstic cavity. However, CO2 concentrations increased toward the end of the cave, reaching 20,000 ppm and temperature increased to 22.7 °C and relative humidity was 100% [13].
The cave is also characterized by anthropic alterations, either on the surface with the urbanization of the area and exploitation of quarries. Inside the cave, many adaptations for facilitating the visits were adopted, including introduction of building materials foreign to the karst, lighting, construction of an artificial lake, artificial buckets with water, installation of an elevator, etc. [14]. A zone of the cave, Galeria Breuil, with rock art, is out of the tourist trail.
The walls of the cave are generally affected by processes of surface corrosion, favored in some galleries by microenvironmental conditions. The most conspicuous effect of lighting was the development of phototrophic communities of cyanobacteria, green algae and mosses on walls and speleothems, coinciding with the lighting points (Figure 3). These areas were cleaned in 2014.
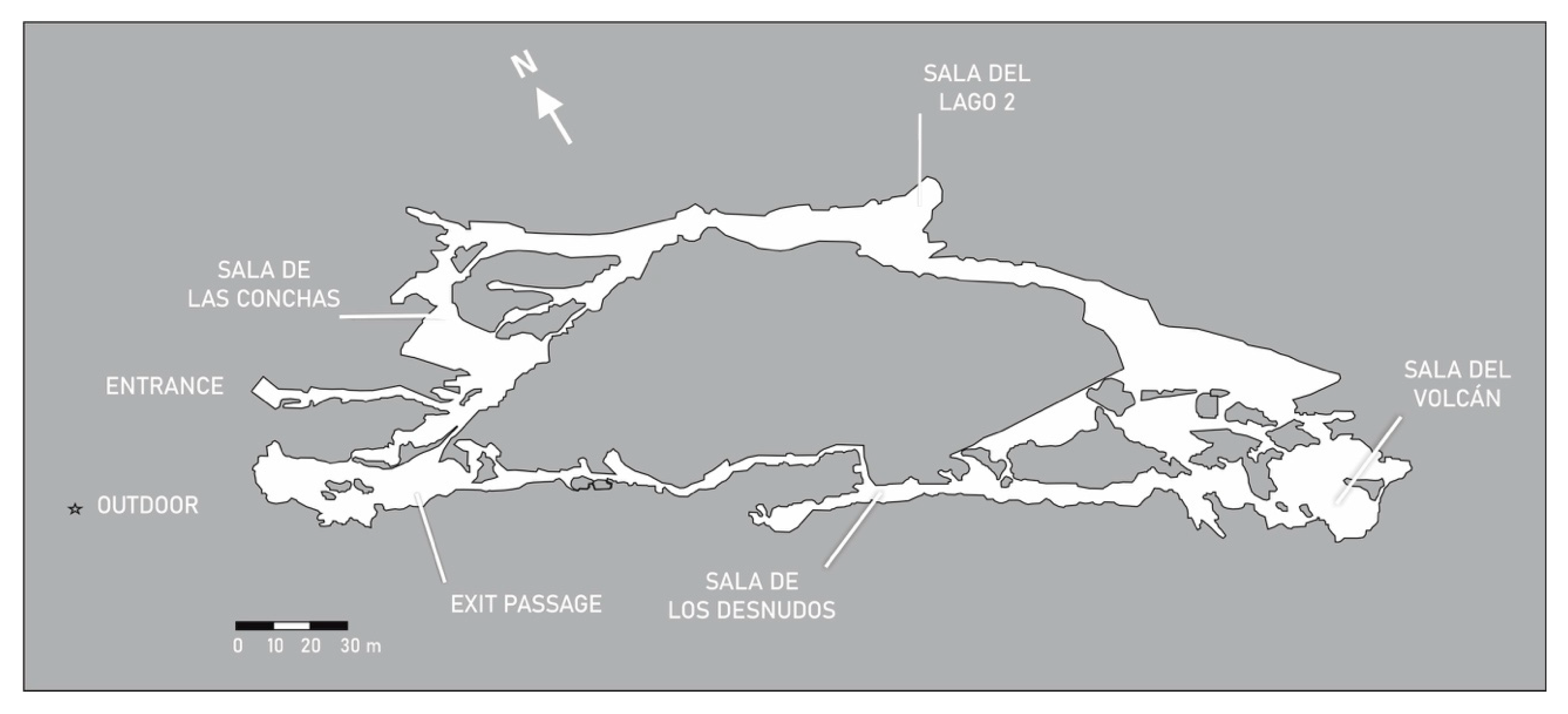
Gruta de las Maravillas (Figure 4) is located in Cerro del Castillo hill, a Precambrian–Cambrian sequence belonging to the so-called Aracena Metamorphic Belt, within the Iberian Massif and formed by marbles, quartzites, granodiorites and gneisses [15]. The cave, within the urban area of Aracena town, has three levels of superimposed galleries in a predominantly horizontal trail, with a length of 2130 m. The two lower levels are conditioned for the visit, about 1000 m in length, while the third, higher, is inaccessible to the public [16]. The cave comprises long narrow passages with dimensions under 5 m and large cavities over 40 m height.
The cavity’s tourist interest lies, on the one hand, in its great variety of calcite and aragonite speleothems, especially in the intermediate level, and on the other, in the flooded galleries of the lower level, where a large number of underwater lamps illuminate various endokarstic forms [17].
The cave showed a zoning of air temperature in different sectors with the highest values in the deeper galleries and the lowest temperatures in Sala del Lago (15.7–16.1 °C) similar to groundwater temperature (15.9–16.3 °C). In the areas furthest from the entrance the temperature can exceed 19.5 °C, corresponding to narrow areas and unfavorable ventilation conditions (Sala del Volcan). The temperature increased with the visits [18].
The concentration of CO2 in the air increased notably with visitors, even tripling the values prior to the visit. In the room closest to the entrance, the concentrations of CO2 ranged between 500 and 600 ppm, while, in the most remote and poorly ventilated areas, the lowest value was 1500 ppm. The influence of the visitors provoked an increase in concentration, reaching values of up to 5000 ppm [18].
2.2. Aerobiology
The aerobiological analysis allowed achieving a representative inventory of the spores of cultivable fungi present in the three caves: Cueva de Ardales, Cueva del Tesoro, and Gruta de las Maravillas. Four sampling campaigns corresponding to a season were carried out between May 2011 and March 2012. In each campaign, representative cave locations inside the cave and one outside were sampled. The sampling points corresponded to the different rooms and galleries and varied depending on the cave length (Figure 5). A total of 68 samples, plus duplicates, were taken over the four seasons: 16 samples from Cueva de Ardales, 28 from Cueva del Tesoro and 24 from Gruta de las Maravillas.
The sampling was performed with a Surface Air System, Duo SAS Super 360 (International pBI, Milan, Italy) installed one meter above the ground. Two replicates of air samples (100 L each one) were directly recovered onto 90 mm Petri dishes. The culture medium used was Dichloran Rose Bengal—Chloramphenicol—Agar (Merck, Darmstad, Germany). The volume of air sampled was selected according to the data of Porca et al. [7] and proved to be the most efficient for an accurate counting. The methodology used has been exhaustively described in previous works [8,9,11].
Briefly, the culture plates were incubated at 25 °C for five days [8]. The colonies were counted, corrected and expressed as colony-forming units per cubic meter, following the Duo SAS sampler manufacturer’s instructions. Morphologically different fungal colonies were isolated on malt extract agar medium (MEA) and stored on MEA slant at 4 °C [9].
Fungal mycelia were collected from the plates and transferred to a 1.5 mL Eppendorf tube containing 500 µL TNE buffer (10 mM Tris–HCl, 100 mM NaCl, 1 mM EDTA; pH 8) and glass beads. The mixture was shaken in a cell disrupter (Fast Prep-24, Solon, OH, USA) at full speed for 3 min.
The DNA was purified by phenol/chloroform extraction and ethanol precipitation [9]. The fungal internal transcribed spacer (ITS) regions, including ITS1, 5.8S rDNA and ITS2, were amplified using the primers ITS1 (5′-TCCGTAGGTGAACCTGCGG-3′) and ITS4 (5′-TCCTCCGCTTATTGATATGC-3′). PCR amplifications were performed in a FlexCycler thermal cycler (Analytik Jena, Jena, Germany) using the following thermal conditions: 94 °C for 2 min; followed by 35 cycles of 94 °C for 1 min, 50 °C for 1 min and 72 °C for 2 min; and a final extension cycle at 72 °C for 5 min [4]. Amplification products were evaluated by electrophoresis on 1% (w/v) agarose gels, stained with SYBR Green I (Roche Diagnostics, Mannheim, Germany) and visualized under UV light [9].
Positive PCR products were sent to Macrogen Inc. (Amsterdam, The Netherlands) for sequencing. For the phylogenetic identification of strains, the sequences were compared, using the BLASTn algorithm, to the non-redundant databases of sequences deposited at the National Center for Biotechnology Information (NCBI). The sequences were deposited into the GenBank database under accession numbers MZ159761–MZ159899.
3. Results
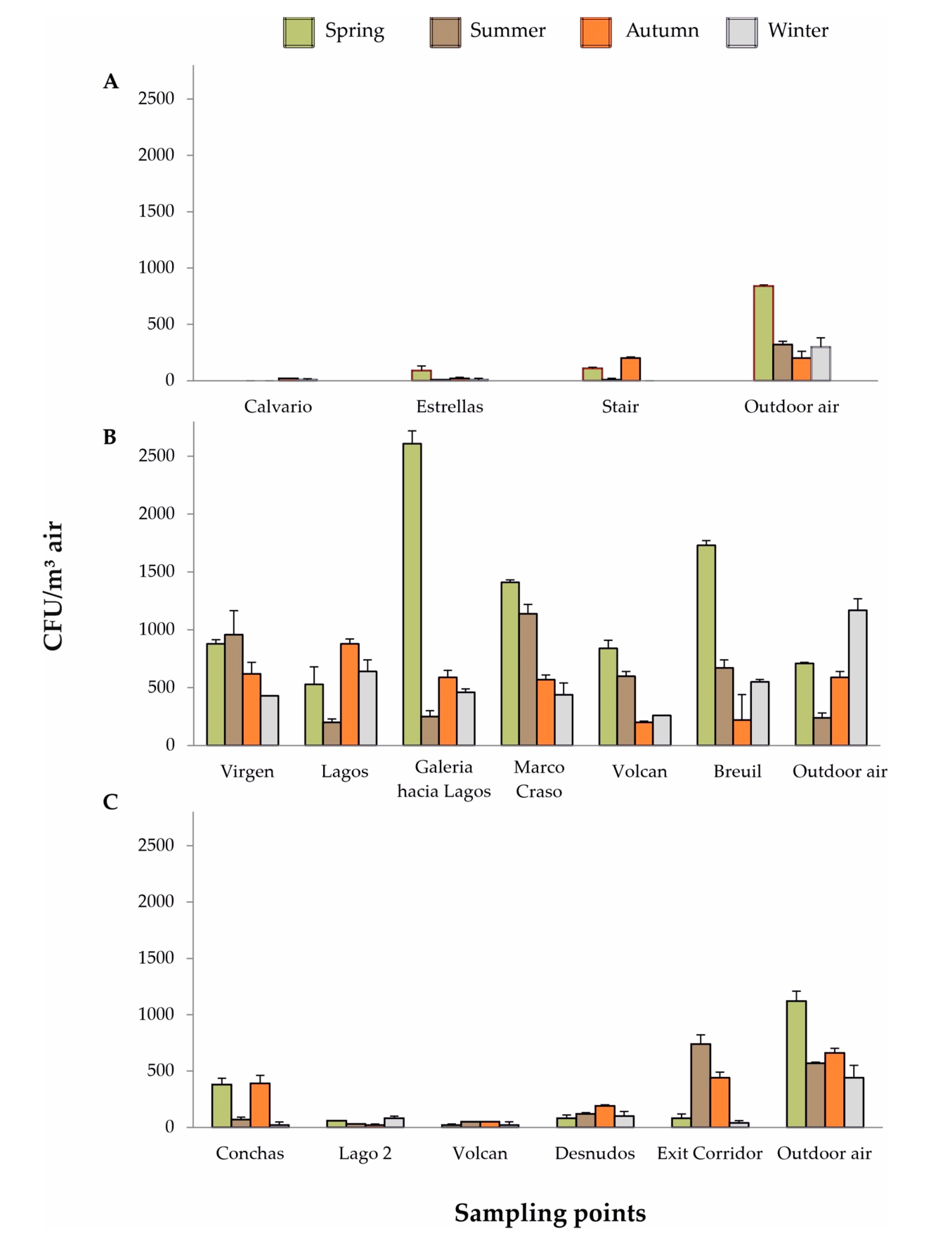
A comparison of the highest CFU/m3 in rooms and galleries over the four seasons in Cueva de Ardales, Cueva del Tesoro, and Gruta de las Maravillas showed that there were large differences among the caves (Figure 5, Supplementary Tables S1–S12). Cueva de Ardales was the cave that was less contaminated by airborne fungi, which is in agreement with the topography and number of visitors. Gruta de las Maravillas contamination was between the other two, due to the high volume of some of the rooms, in spite of receiving the higher number of visitors. To this may contribute the cave management (frequent cleaning of the rooms and galleries). The most contaminated cave is Cueva del Tesoro, that has a medium number of visitors, but two factors contributed to this pollution, the small volume of galleries and rooms and the abundance of phototrophic biofilms along the cave, induced by the artificial lighting (Figure 3).
3.1. Cueva de Ardales
This cave presented the lowest spore concentration from all the three caves investigated, in addition to the lowest diversity as only 10 different fungal species were collected. The cave morphology is a U, where the entrance is located at one of the ends and in the other one Galeria del Calvario; the huge Sala de las Estrellas is located at the bottom (Figure 1).
Supplementary Tables S1–S4 shows the fungal CFU/m3 (abundances > 10%) of the sampling points and seasons. It is noteworthy that in spring and summer samplings, no spores were collected in Galeria del Calvario, whereas in winter only 10 CFU/m3 and in autumn 20 CFU/m3 were retrieved. This was due to the morphology of the cave because this gallery is the highest topographical area where practically no aerodynamic processes take place between the outer atmosphere and the rest of the cave.
On the other hand, in spring and autumn the high concentrations of spores outdoor (840 and 200 CFU/m3, respectively) were reflected at the end of the stairs leading to the main cave room (110 and 200 CFU/m3, respectively), next to the entrance, but not in winter when no spores were collected at this point.
The most abundant fungi identified in Cueva de Ardales were Cladosporium cladosporioides in spring, Beauveria bassiana in summer, Parengyodontium album in autumn, and Leptobacillium leptobactrum and Cladosporium cladosporioides in winter. Outdoor, Cladosporium cladosporioides was abundant in all seasons.
3.2. Cueva del Tesoro
This cave presents an unusual morphology, with an entrance in the middle of the cave (Figure 2), accessing Sala de la Virgen through an elevator and stairs and an air entrance in Sala de Lagos. In addition, at the sampling time, the speleothems and cave walls were abundantly coated by phototrophic biofilms (Figure 3) that were further cleaned in 2014 [14].
Regarding the fungal concentrations recorded in the three cavities throughout the seasonal samplings, the highest values of CFU/m3 were found in Cueva del Tesoro. This can be explained by the relatively small dimensions of the rooms and galleries, which could contribute to the concentration of spores, in addition to the abundance of phototrophic biofilms all over the cave. These concentrations were up to 3.6 times higher inside in spring and 4.75 times higher in summer. Eighteen different species were recorded in this cave.
Supplementary Tables S5–S8 list the fungal CFU/m3 (abundances > 10%) in the different sampling points and seasons. In spring, the most abundant fungi were Parengyodontium album, Pochonia chlamydosporia and Aspergillus nidulans; in summer Pochonia chlamydosporia and Leptobacillium symbioticum; and in autumn and winter Cladosporium cladosporioides.
It is noticeable that the CFU/m3 inside the cave was considerable higher than those registered outdoor in all seasons, except in winter, a pattern opposite to the other two caves. Thus, for example, in spring the higher rates were 2610 CFU/m3 in Galeria hacia Lagos and 1730 CFU/m3 in Galeria Breuil, whereas outside was 710 CFU/m3.
3.3. Gruta de las Maravillas
Supplementary Tables S9–S12 present the fungal CFU/m3 (abundances > 10%) in the different sampling points and seasons. Twenty two different fungi were isolated in the air of this cave. In spring, the most abundant fungus was Pochonia chlamydosporia; in summer Pochonia chlamydosporia and Penicillium chrysogenum; in autumn Pochonia chlamydosporia; and in winter Leptobacillium symbioticum. The difference in terms of biodiversity of this cave with respect to the other two caves was the presence of the yeast genera Cutaneotrichosporon, Trichosporon, Cryptococcus, Cystobasidium, Microstroma, Phragmotaenium, and Naganishia which were among the most abundant fungi in all seasons except spring.
4. Discussion
4.1. Fungal Ecology
The influence of air currents from the outdoors was observed in the three caves, which transport spores, giving rise to an increase in fungal concentrations in the areas of the cavity closest to the entrance. Fernandez-Cortes et al. [6] reported that in Cueva de Ardales, and as a consequence of the opening of the access door for the entrance of visitors, there was an increase in the concentrations of fungi. This was particularly high in Galeria del Calvario, the zone where the paintings and engravings are located, which increased by 100 times in terms of CFU/m3 after a visit of 32 people. In our sampling, one year later and without the influence of previous visits, the CFU/m3 in Galeria del Calvario was zero or negligible (Supplementary Tables S1–S4). These data note that the visits should be controlled or, if needed, cancelled in caves with rock art.
In Gruta de las Maravillas, Sala de las Conchas, one of the areas closest to the entrance, the concentrations of fungi followed the same seasonal patterns observed in the outdoor air, highlighting the influence of the exterior atmosphere. The large volume of other rooms seemed to influence fungal concentrations, causing a dilution effect. Thus, the highest fungal concentrations were recorded in rooms and galleries of relatively small dimensions, which could contribute to the concentration of spores, as was the case in Cueva del Tesoro.
Regarding the ecology of the most abundant fungal genera in the three caves, it was found that Cladosporium was the predominant genus inside the three caves. Six species were identified, C. cladosporioides, C. halotolerans, C. tenuissimun, C. limoniforme, C. ramotenellum, and C. floccosum, in addition to two species outdoor: C. sphaerospermum and C. subinflatum.
The abundance of Cladosporium is a common pattern in Nerja and Altamira, among other caves [8,12], in addition to indoor environments [19,20]. The genus Cladosporium is one of the most frequent in outdoor environments, so its presence indoors is generally attributed to transport through air currents [6,8].
According to Fernandez-Cortes et al. [6] the higher concentrations of Cladosporium, in a sampling carried out after a visit of 40 people in Cueva de Ardales, showed the influence of the opening of the door in the creation of air currents between the interior and exterior of the cavity, with the consequent entry of spores. Cladosporium cladosporioides, one of the most abundant fungi in Cueva de Ardales and Cueva del Tesoro has been recorded in other caves, such as Lascaux [2], Altamira [8], and three Slovakian caves [21].
Pochonia chlamydosporia (=Metacordyceps chlamydosporia), a recognized entomopathogen [22,23], appeared distributed throughout the rooms of Cueva del Tesoro in summer and autumn, and in Gruta de las Maravillas in all season, except winter. Their presence in all seasons except in winter is remarkable. The occurrence is coincident with the bloom of insects in spring and death in autumn. This fungus was one of the most abundant species in Castañar de Ibor Cave [7].
Other abundant entomopathogenic species in the caves were Parengyodontium album, Leptobacillium symbioticum, Leptobacillium leptobactrum and Beauveria bassiana.
Parengyodontium album is related to spiders and mites. It is normally found in underground environments, such as catacombs [24] and caves [2]. This was the predominant species inside Cueva de Ardales in autumn. At the end of the entrance stair, the sampling provided 200 CFU/m3 and P. album presented an abundance of 91.9%. The high concentration of P. album in this sampling point may be due to the fact that being closer to the exterior, it is more frequented by arthropods than other deeper cavity zones.
The species of the genus Leptobacillium are saprotrophic, and includes entomopathogenic strains, such as L. symbioticum and L. leptobactrum (=Verticillium leptobactrum) [25,26]. Leptobacillium symbioticum was predominant in winter in all the rooms of Gruta de las Maravillas, except in Sala de las Conchas, and appeared in a few rooms in summer (Sala de las Conchas) and Sala del Lago 2 in autumn. This fungus was also present in all the show rooms in Cueva del Tesoro. Leptobacillium leptobactrum was identified in Cueva de Ardales (Sala de las Estrellas), in spring and winter. In Gruta de las Maravillas (Sala del Volcan), both L. symbioticum and L. leptobactrum amounted 100% of the fungi isolated in winter. Leptobacillium leptobactrum was previously recorded as Verticillium leptobactrum, in the air of Lascaux, a cave densely populated by arthropods [2,9,27].
The abundance of the genus Leptobacillium in winter may be due to the fact that their species are parasitic of mites [25] and lepidopteran larvae and pupae [26], coinciding with the fact that most troglophiles species enter a dormant state in the winter season.
The entomopathogenic Beauveria bassiana is found naturally in soils and on insects and mites, with the capacity to infect different species, which is why it is widely used for pest control [28]. This species was identified in the end of the stair in Cueva de Ardales in spring and summer samplings and in Cueva del Tesoro in winter (Sala de la Virgen). It has also been recorded in Lascaux Cave [2], and Domica Cave [21].
An additional and single record of entomopathogenic fungus was Lecanicillium tenuipes, majority in Cueva del Tesoro, Sala de Lagos, in summer. This species was isolated from different insects [29,30].
Schizophyllum commune was one of the most abundant fungus in Cueva del Tesoro, (Sala de la Virgen, Sala de Lagos) and outdoor in spring, and in Sala de Marco Craso in summer. This basidiomycete is a saprophytic fungus that grows on decaying plants or wood [31]. The presence of basidiomycetes is usually common in caves [11,21] and could be associated with the presence of structures and/or wood fragments, common in caves, such as Altamira, Nerja, etc. Therefore, the presence of S. commune in Sala de la Virgen, could be attributed to the wooden platform found in this room, although their transport from outdoor cannot be discarded due to the proximity of this room to the entrance.
The genera Aspergillus and Penicillium were the most abundant in Cueva de Nerja [12]. This trend was also found in Cueva del Tesoro, located some 40 km from Nerja. Four Aspergillus species, A. versicolor, A. thesauricus, A. nidulans and A. parasiticus, were retrieved in this cave. Aspergillus thesauricus was described from strains isolated for the first time in Cueva del Tesoro [32]. A few Penicillium, P. citreonigrum, P. lanosum, P. consobrinum, and P. chrysogenum were found in Cueva del Tesoro, whereas P. chrysogenum, P. minioluteum, P. flavigenum, and P. brevicompactum were isolated from Gruta de las Maravillas; where P. chrysogenum reached an abundance of 82.7% in Sala de los Desnudos.
Cueva de Nerja and Cueva del Tesoro, in addition to the short distance from one to another, have in common the presence of abundant phototrophic biofilms on the speleothems and walls due to the artificial lighting installed for facilitating the visits. The abundance of these two genera could be related with the cyanobacteria and algae biofilms populating the two caves. Grbić et al. [33] and Gallego-Cartagena et al. [34] found a close association between Aspergillus and Penicillium spp. and cyanobacterial biofilms.
It should be noted the presence of yeasts of the genera Cutaneotrichosporon, Trichosporon, Cryptococcus, Naganishia, Cystobasidium, Microstroma and Phragmotaenium in Gruta de las Maravillas and their absence in the other two caves could be related to the high number of visitors. The presence of yeasts in the caves was related with the frequentation of people and animals [35]. Cutaneotrichosporon, Trichosporon and Cryptococcus species have been isolated from bat guano in caves in Japan [36,37]. Cutaneotrichosporon moniliiforme was the most abundant species in Gruta de las Maravillas (Sala de las Conchas) in summer, which was also identified in the air of Castañar de Ibor Cave [38]. In Gruta de las Maravillas, Trichosporon asteroides was also isolated from two rooms in summer. Species of the genus Phragmotaenium are commonly cited as constituents of the phylloplane microflora, especially in leaves infected by powdery mildew, some of these species being of interest as biological control agents for other fungi [39]. Naganishia species are ubiquitous saprophytic yeasts, frequently isolated from different environments, and can behave as opportunistic pathogens [40]. Naganishia diffluens was found in Gruta de las Maravillas (Sala del Lago 2) in autumn and winter, and this and other yeasts were associated with insects [41].
A few more Basidiomycota species were isolated from the caves. Ustilago cynodontis in Cueva de Ardales, Emmia latemarginata, Peniophora pilatiana and Paraconiothyrium fuckelii in Gruta de las Maravillas, and Bjerkandera adusta and Sistotrema sp. in Cueva del Tesoro. The sampling sites where these fungi were retrieved, near the entrance and outdoors, point to transport into the caves. A similar origin can be suggested for some Ascomycota, endophytic or plant pathogens, such as Cadophora malorum, Cephalotrichum purpureofuscum and Cyphellophora olivacea in Gruta de las Maravillas, Alternaria alternata, Acrodontium pigmentosum and Nectria pseudotrichia in Cueva de Ardales, and Gnomoniopsis idaeicola and Cytospora acaciae in Cueva del Tesoro.
4.2. Fungal Hazard in Show Caves
Porca et al. [7] established a classification or index of fungal hazard in European show caves with rock art, based on the concentration of spores in the cave air. Five categories were established, from <50 CFU/m3 to >1000 CFU/m3. The application of this index to the Andalusian caves revealed that Cueva de Ardales was the least contaminated, which agrees with the fact that it is the cave with the lowest number of visits. This cavity corresponded to a category 2 (between 50 and 150 CFU/m3, which represents an alarm signal and some periodic controls and studies are required to disclose the problem). However, CFU/m3 can increase on specific occasions, for example after group visits, as Fernandez-Cortes et al. [6] pointed out, particularly in Galeria del Calvario, which reached 1010 CFU/m3 and is the most sensitive area to visits.
Gruta de las Maravillas, despite the high number of visitors that it receives, had relatively low fungal concentrations, being higher in Sala de las Conchas and in the exit passage, which are the areas closest to the entrance. This cavity must be classified as category 3, as a cave threatened by fungi, reaching between 150 and 500 CFU/m3 and for which a different cave management (visits, air currents, closure of doors, etc.) and controls are necessary. However, in Gruta de las Maravillas two factors intervene to cause the fungal concentrations to not be very high, considering the number of visitors/year; on the one hand, the large volume of the rooms and, on the other, periodic maintenance and cleaning tasks.
The minimum values of CFU/m3 recorded in Cueva del Tesoro were equivalent to the maximums in the other two caves. The factors that can influence the high concentrations are: two connections with the exterior, relatively small dimensions of its rooms and galleries, and the abundant presence of phototrophic communities on the speleothems and walls [14]. This cave would be in a category 5 (caves that suffer irreversible ecological damage and potentially harmful to the conservation of paintings, with concentrations of fungi higher than 1000 CFU/m3 of air). However, the phototrophic biofilms were removed and the speleothems and walls were cleaned in 2014 [14], after this study, which probably at the present time has reduced the number of fungal spores in the air.
Regarding the potential danger to human health, the most abundant fungi recorded in the caves have been considered opportunistic pathogens. They encompass Cladosporium cladosporioides [42], Beauveria bassiana [43], Parengyodontium album [44], Schyzophyllum commune [45], Aspergillus nidulans [46], Penicillium chrysogenum [47], in addition to basidiomycetous yeasts [48,49,50]. Jurado et al. [51] reviewed the role of pathogenic and opportunistic microorganisms in caves and concluded that “the most-frequent diseases produced by cave microorganisms are located in the respiratory system and, therefore, it is recommended that visitors should, at the least, wear protective masks”.
Another important aspect is the biodeterioration caused by fungi thriving in caves. The most recurrent species isolated from rocks and speleothems and reported as producing deterioration are Cladosporium cladosporioides, Parengyodontium album and Penicillium chrysogenum [52,53,54,55]. Fungal deterioration can be evidenced by observation of structural and chemical changes using microscopic and spectroscopic techniques [56,57].
5. Conclusions
The most characteristic features in the three Southern Spain show caves, Cueva del Tesoro, Cueva de Ardales and Gruta de las Maravillas, were the abundant presence of entomopathogenic fungi (Beauveria bassiana, Parengyodontium album, Pochonia chlamydosporia, Leptobacillium symbioticum, Leptobacillium leptobactrum) and Cladosporium cladosporioides. The presence of yeasts of the genera Cutaneotrichosporon, Trichosporon, Cryptococcus, Naganishia, Cystobasidium, Microstroma and Phragmotaenium was restricted to Gruta de las Maravillas. The caves showed different levels of fungal hazards, as determined using an ecological indicator based on the concentration of spores in cave air. The threats were mainly linked to the cave morphology, management, and number of visitors.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/app11115027/s1, Table S1: Sampling campaign in Cueva de Ardales, spring 2011. CFU/m3, accession numbers, identifications and abundances, Table S2: Sampling campaign in Cueva de Ardales, summer 2011. CFU/m3, accession numbers, identifications and abundances, Table S3: Sampling campaign in Cueva de Ardales, autumn 2011. CFU/m3, accession numbers, identifications and abundances, Table S4: Sampling campaign in Cueva de Ardales, winter 2012. CFU/m3, accession numbers, identifications and abundances, Table S5: Sampling campaign in Cueva del Tesoro, spring 2011. CFU/m3, accession numbers, identifications and abundances, Table S6: Sampling campaign in Cueva del Tesoro, summer 2011. CFU/m3, accession numbers, identifications and abundances, Table S7: Sampling campaign in Cueva del Tesoro, autumn 2011. CFU/m3, accession numbers, identifications and abundances, Table S8: Sampling campaign in Cueva del Tesoro, winter 2012. CFU/m3, accession numbers, identifications and abundances, Table S9: Sampling campaign in Gruta de las Maravillas, spring 2011. CFU/m3, accession numbers, identifications and abundances, Table S10: Sampling campaign in Gruta de las Maravillas, summer 2011. CFU/m3, accession numbers, identifications and abundances, Table S11: Sampling campaign in Gruta de las Maravillas, autumn 2011. CFU/m3, accession numbers, identifications and abundances, Table S12: Sampling campaign in Gruta de las Maravillas, winter 2012. CFU/m3, accession numbers, identifications and abundances. Tables S1 to S12.
Author Contributions
Conceptualization, C.S.-J.; investigation, I.D.-M., V.J., M.A.R.-C. and B.H.; writing—original draft preparation, C.S.-J.; writing—review and editing, C.S.-J. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Junta de Andalucia, grant number RNM-5137. I.D.-M. acknowledges the pre-doctoral grant associated to project RNM-5137.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data, under accession numbers MZ159761–MZ159899, were deposited into the GenBank database.
Acknowledgments
The authors wish to acknowledge the professional support of the CSIC Interdisciplinary Thematic Platform Open Heritage: Research and Society (PTI-PAIS) as well as the facilities of the cave managers for sampling.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dupont, J.; Jacquet, C.; Dennetiere, B.; Lacoste, S.; Bousta, F.; Orial, G.; Cruaud, C.; Couloux, A.; Roquebert, M.F. Invasion of the French Paleolithic painted cave of Lascaux by members of the Fusarium solani species complex. Mycologia 2007, 99, 526–533. [Google Scholar]
- Bastian, F.; Alabouvette, C.; Saiz-Jimenez, C. The impact of arthropods on fungal community structure in Lascaux Cave. J. Appl. Microbiol. 2009, 106, 1456–1462. [Google Scholar] [CrossRef]
- Martin-Sanchez, P.M.; Nováková, A.; Bastian, F.; Alabouvette, C.; Saiz-Jimenez, C. Use of biocides for the control of fungal outbreaks in subterranean environments: The case of the Lascaux Cave in France. Environ. Sci. Technol. 2012, 46, 3762–3770. [Google Scholar] [CrossRef] [PubMed]
- Jurado, V.; Porca, E.; Cuezva, S.; Fernandez-Cortes, A.; Sanchez-Moral, S.; Saiz-Jimenez, C. Fungal outbreak in a show cave. Sci. Total Environ. 2010, 408, 3632–3638. [Google Scholar] [CrossRef] [Green Version]
- Caneva, G.; De Nuntiis, P.; Fornaciari, M.; Ruga, L.; Valenti, P.; Pasquariello, G. Aerobiology applied to the preventive conservation of cultural heritage. Aerobiologia 2020, 36, 99–103. [Google Scholar] [CrossRef]
- Fernandez-Cortes, A.; Cuezva, S.; Sanchez-Moral, S.; Porca, E.; Jurado, V.; Saiz-Jimenez, C. Detection of human-induced environmental disturbances in a show cave. Environ. Sci. Pollut. Res. 2011, 18, 1037–1045. [Google Scholar] [CrossRef] [Green Version]
- Porca, E.; Jurado, V.; Martin-Sanchez, P.M.; Hermosin, B.; Bastian, F.; Alabouvette, C.; Saiz-Jimenez, C. Aerobiology: An ecological indicator for early detection and control of fungal outbreaks in caves. Ecol. Indic. 2011, 11, 1594–1598. [Google Scholar] [CrossRef]
- Garcia-Anton, E.; Cuezva, S.; Jurado, V.; Porca, E.; Miller, A.Z.; Fernandez-Cortes, A.; Saiz-Jimenez, C.; Sanchez-Moral, S. Combining stable isotope (δ13C) of trace gases and aerobiological data to monitor the entry and dispersion of microorganisms in caves. Environ. Sci. Pollut. Res. 2014, 21, 473–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Sanchez, P.M.; Jurado, V.; Porca, E.; Bastian, F.; Lacanette, D.; Alabouvette, C.; Saiz-Jimenez, C. Airborne microorganisms in Lascaux Cave (France). Int. J. Speleol. 2014, 43, 295–303. [Google Scholar] [CrossRef]
- Nováková, A.; Kubátová, A.; Sklenář, F.; Hubka, V. Microscopic fungi on cadavers and skeletons from cave and mine environments. Czech Mycol. 2018, 70, 101–121. [Google Scholar] [CrossRef]
- Sanchez-Moral, S.; Jurado, V.; Fernandez-Cortes, A.; Cuezva, S.; Martin-Pozas, T.; Gonzalez-Pimentel, J.L.; Ontañon, R.; Saiz-Jimenez, C. Environment-driven control of the dispersal of fungi in subterranean ecosystems: The case of La Garma Cave (northern Spain). Int. Microbiol. 2021. submitted. [Google Scholar]
- Docampo, S.; Trigo, M.M.; Recio, M.; Melgar, M.; García-Sánchez, J.; Cabezudo, B. Fungal spores content of the atmosphere of the Cave of Nerja (southern Spain): Diversity and origin. Sci. Total Environ. 2011, 409, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Moñino, I. Evaluación y Control de Comunidades Microbianas en Cuevas Turísticas. Ph.D. Thesis, Instituto de Recursos Naturales y Agrobiología, Sevilla, Spain, 2015. [Google Scholar]
- Jurado, V.; Hernández-Mariné, M.; Rogerio-Candelera, M.A.; Domínguez-Moñino, I.; Gutiérrez-Patricio, S.; Saiz-Jiménez, C. La Cueva del Tesoro: Un Paraíso Microbiano. In Cuevatur 2014/Iberoamérica Subterránea; Calaforra, J.M., Durán, J.J., Eds.; ACTE: Madrid, Spain, 2014; pp. 431–439. [Google Scholar]
- Martínez-Moreno, F.J.; Galindo-Zaldívar, J.; Pedrera, A.; Teixido, T.; Ruano, P.; Peña, J.A.; González-Castillo, L.; Ruiz-Constán, A.; López-Chicano, M.; Martín-Rosales, W. Integrated geophysical methods for studying the karst system of Gruta de las Maravillas (Aracena, Southwest Spain). J. Appl. Geophys. 2014, 107, 149–162. [Google Scholar] [CrossRef]
- Pulido-Bosch, A.; Martín-Rosales, W.; López-Chicano, M.; Vallejos, A. Human impacts in a touristic cave (Aracena, Spain). Environ. Geol. 1997, 31, 142–149. [Google Scholar] [CrossRef] [Green Version]
- Martín-Rosales, W.; López-Chicano, M.; Rodríguez, C.M.; Pulido-Bosch, A. Morfología, espeleotemas y climatología de la Gruta de las Maravillas, Aracena (Huelva). Espeleotemas 1995, 5, 1–12. [Google Scholar]
- Martín-Rosales, W.; Cerón, J.C.; López-Chicano, M.; Fernández, I. Aspectos ambientales e hidrogeológicos de la Gruta de las Maravillas (Huelva, España). Bol. Geol. Min. 2003, 114, 247–254. [Google Scholar]
- Bensch, K.; Groenewald, J.Z.; Meijer, M.; Dijksterhuis, J.; Jurjevic, Z.; Andersen, B.; Houbraken, J.; Crous, P.W. Cladosporium species in indoor environments. Stud. Mycol. 2018, 89, 177–301. [Google Scholar] [CrossRef]
- Segers, F.J.J.; van Laarhoven, K.A.; Huinink, H.P.; Adan, O.C.G.; Wösten, H.A.B.; Dijksterhuis, J. The indoor fungus Cladosporium halotolerans survives humidity dynamics markedly better than Aspergillus niger and Penicillium rubens despite less growth at lowered steady-state water activity. Appl. Environ. Microbiol. 2016, 82, 5089–5098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nováková, A. Microscopic fungi isolated from the Domica Cave system (Slovak Karst National Park, Slovakia). A review. Int. J. Speleol. 2009, 38, 71–82. [Google Scholar] [CrossRef] [Green Version]
- Kepler, R.M.; Sung, G.H.; Ban, S.; Nakagiri, A.; Chen, M.J.; Huang, B.; Li, Z.; Spatafora, J.W. New teleomorph combinations in the entomopathogenic genus Metacordyceps. Mycologia 2012, 104, 182–197. [Google Scholar] [CrossRef]
- Kim, H.S.; Adhikari, M.; Yadav, D.R.; Kim, S.W.; Um, Y.H.; Lee, H.B.; Lee, Y.S. First report of Metacordyceps chlamydosporia (Cordyceps chlamydosporia) isolated from soil in Korea. Korean J. Med. Mycol. 2016, 44, 48–50. [Google Scholar] [CrossRef] [Green Version]
- Jurado, V.; Sanchez-Moral, S.; Saiz-Jimenez, C. Entomogenous fungi and the conservation of the cultural heritage: A review. Int. Biodeter. Biodegr. 2008, 62, 325–330. [Google Scholar] [CrossRef] [Green Version]
- Zare, R.; Gams, W. More white verticillium-like anamorphs with erect conidiophores. Mycol. Progress 2016, 15, 993–1030. [Google Scholar] [CrossRef]
- Okane, I.; Nonaka, K.; Kurihara, Y.; Abe, J.P.; Yamaoka, Y. A new species of Leptobacillium, L. symbioticum, isolated from mites and sori of soybean rust. Mycoscience 2020, 61, 165–171. [Google Scholar] [CrossRef]
- Bastian, F.; Jurado, V.; Nováková, A.; Alabouvette, C.; Saiz-Jimenez, C. The microbiology of the Lascaux Cave. Microbiology 2010, 156, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Santamaría, A.; Costa-Comelles, J.; Alonso, A.; Rodríguez, J.M.; Ferrer, J. Ensayo del hongo entomopatógeno Beauveria bassiana (Balsamo) Vuillemin para el control de la mosca blanca de los cítricos Aleurothrixus floccosus (Maskell) (Homoptera: Aleyrodidae) y su acción sobre el parásito Cales noacki (Howard) (Hymenoptera: Aphelinidae). Bol. San. Veg. Plagas 1998, 24, 695–706. [Google Scholar]
- Zare, R.; Gams, W. A revision of Verticillium section Prostrata. IV. The genera Lecanicillium and Simplicillium gen. nov. Nova Hedwigia. 2001, 73, 1–50. [Google Scholar] [CrossRef]
- Castillo, S.Y.; Bernal Vega, J.A.; Lezcano, J.; Piepenbring, M.; Cáceres, O. Hongos entomopatógenos asociados a insectos recolectados en plantaciones de café en el oeste de Panamá. Tecnociencia 2013, 15, 29–39. [Google Scholar]
- Chang, S.; Lui, W. Analysis of the mating types of Schizophyllum commune in the natural population of Hong Kong. Bot. Bull. Acad. Sin. 1969, 10, 74–88. [Google Scholar]
- Nováková, A.; Hubka, V.; Saiz-Jimenez, C.; Kolarik, M. Aspergillus baeticus sp. nov. and Aspergillus thesauricus sp. nov.: Two new species in section Usti originating from Spanish caves. Int. J. Syst. Evol. Microbiol. 2012, 62, 2778–2785. [Google Scholar] [CrossRef] [PubMed]
- Grbić, M.L.; Vukojević, J.; Simić, G.S.; Krizmanić, J.; Stupar, M. Biofilm forming cyanobacteria, algae and fungi on two historic monuments in Belgrade, Serbia. Arch. Biol. Sci. 2010, 62, 625–631. [Google Scholar] [CrossRef]
- Gallego-Cartagena, E.; Morillas, H.; Maguregui, M.; Patiño-Camelo, K.; Marcaida, I.; Morgado-Gamero, W.; Silva, L.F.O.; Madariaga, J.M. A comprehensive study of biofilms growing on the built heritage of a Caribbean industrial city in correlation with construction materials. Int. Biodeter. Biodegr. 2020, 147, 104874. [Google Scholar] [CrossRef]
- Vaughan-Martini, A.; Angelini, P.; Zacchi, L. The influence of human and animal visitation on the yeast ecology of three Italian caverns. Ann. Microbiol. 2000, 50, 133–140. [Google Scholar]
- Sugita, T.; Kikuchi, K.; Makimura, K.; Urata, K.; Someya, T.; Kamei, K.; Niimi, M.; Uehara, Y. Trichosporon species isolated from guano samples obtained from bat-inhabited caves in Japan. Appl. Environ. Microbiol. 2005, 71, 7626–7629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takashima, M.; Kurakado, S.; Cho, O.; Kikuchi, K.; Sugiyama, J.; Sugita, T. Description of four Apiotrichum and two Cutaneotrichosporon species isolated from guano samples from bat-inhabited caves in Japan. Int. J. Syst. Evol. Microbiol. 2020, 70, 4458–4469. [Google Scholar] [CrossRef] [PubMed]
- Porca Belío, E. Aerobiología: Mecanismos de Dispersión de los Microorganismos en Cuevas Turísticas. Ph.D. Thesis, Instituto de Recursos Naturales y Agrobiología, Sevilla, Spain, 2011. [Google Scholar]
- Urquhart, E.J.; Menzies, J.G.; Punja, Z.K. Growth and biological control activity of Tilletiopsis species against powdery mildew (Sphaerotheca fuliginea) on greenhouse cucumber. Phytopathology 1994, 84, 341–351. [Google Scholar] [CrossRef]
- Fonseca, A.; Boekhout, T.; Fell, J.W. Cryptococcus Vuillemin (1901). In The Yeasts, a Taxonomic Study; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; pp. 1661–1737. [Google Scholar]
- Brentassi, M.E.; Medina, R.; de la Fuente, D.; Franco, M.E.E.; Toledo, A.V.; Saparrat, M.C.N.; Balatti, P.A. Endomycobiome associated with females of the planthopper Delphacodes kuscheli (Hemiptera: Delphacidae): A metabarcoding approach. Helyon 2020, 6, e04634. [Google Scholar] [CrossRef]
- Sandoval-Denis, M.; Sutton, D.A.; Martin-Vicente, A.; Cano-Lira, J.F.; Wiederhold, N.; Guarro, J.; Gené, J. Cladosporium species recovered from clinical samples in the United States. J. Clin. Microbiol. 2015, 53, 2990–3000. [Google Scholar] [CrossRef] [Green Version]
- Tucker, D.L.; Beresford, C.H.; Sigler, L.; Rogers, K. Disseminated Beauveria bassiana infection in a patient with acute lymphoblastic leukemia. J. Clin. Microbiol. 2004, 42, 5412–5414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsang, C.-C.; Chan, J.F.W.; Pong, W.-M.; Chen, J.H.K.; Ngan, A.H.Y.; Cheung, M.; Lai, C.K.C.; Tsang, D.N.C.; Lau, S.K.P.; Woo, P.C.Y. Cutaneous hyalohyphomycosis due to Parengyodontium album gen. et comb. nov. Med. Mycol. 2016, 54, 699–713. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Yi, Y.; Cho, S.-Y.; Lee, D.-G.; Chun, H.-S.; Park, C.; Kim, Y.-J.; Park, Y.-J. Pneumonia due to Schizophyllum commune in a patient with acute myeloid leukemia: Case report and literature review. Infect. Chemother. 2021, 53, e67. [Google Scholar]
- Bastos, R.W.; Valero, C.; Silva, L.P.; Schoen, T.; Drott, M.; Brauer, V.; Silva-Rocha, R.; Lind, A.; Steenwyk, J.L.; Rokas, A.; et al. Functional characterization of clinical isolates of the opportunistic fungal pathogen Aspergillus nidulans. mSphere 2020, 5, e00153-20. [Google Scholar] [CrossRef] [Green Version]
- Geltner, C.; Lass-Flörl, C.; Bonatti, H.; Müller, L.; Stelzmüller, I. Invasive pulmonary mycosis due to Penicillium chrysogenum: A new invasive pathogen. Transplantation 2013, 95, e21–e23. [Google Scholar] [CrossRef]
- Cooper, C.R. Yeasts Pathogenic to Humans. In The Yeasts, a Taxonomic Study; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; pp. 9–19. [Google Scholar]
- Mariné, M.; Brown, N.A.; Riaño-Pachón, D.M.; Goldman, G.H. On and under the skin: Emerging basidiomycetous yeast infections caused by Trichosporon species. PLoS Pathog. 2015, 11, e1004982. [Google Scholar] [CrossRef]
- Morales-López, S.E.; Garcia-Effron, G. Infections due to rare Cryptococcus species. A literature review. J. Fungi 2021, 7, 279. [Google Scholar] [CrossRef]
- Jurado, V.; Laiz, L.; Rodriguez-Nava, V.; Boiron, P.; Hermosin, B.; Sanchez-Moral, S.; Saiz-Jimenez, C. Pathogenic and opportunistic microorganisms in caves. Int. J. Speleol. 2010, 39, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Zucconi, L.; Gagliardi, M.; Isola, D.; Onofri, S.; Andaloro, M.C.; Pelosi, C.; Pogliani, P.; Selbmann, L. Biodeterioration agents dwelling in or on the wall paintings of the Holy Saviour’s cave (Vallerano, Italy). Int. Biodeter. Biodegr. 2012, 70, 40–46. [Google Scholar] [CrossRef]
- Ogórek, R.; Dyląg, M.; Višňovská, Z.; Tancinová, D.; Zalewski, D. Speleomycology of air and rock surfaces in Driny Cave (Lesser Carpathians, Slovakia). J. Cave Karst Stud. 2016, 78, 119–127. [Google Scholar] [CrossRef]
- Leplat, J.; François, A.; Bousta, F. Parengyodontium album, a frequently reported fungal species in the cultural heritage environment. Fungal Biol. Rev. 2020, 34, 126–135. [Google Scholar] [CrossRef]
- Mang, S.M.; Scrano, L.; Camele, I. Preliminary studies on fungal contamination of two rupestrian churches from Matera (Southern Italy). Sustainability 2020, 12, 6988. [Google Scholar] [CrossRef]
- Pyzik, A.; Ciuchcinski, K.; Dziurzynski, M.; Dziewit, L. The Bad and the good—Microorganisms in cultural heritage environments—An update on biodeterioration and biotreatment approaches. Materials 2021, 14, 177. [Google Scholar] [CrossRef] [PubMed]
- Saiz-Jimenez, C.; Miller, A.Z.; Martin-Sanchez, P.M.; Hernandez-Marine, M. Uncovering the origin of the black stains in Lascaux Cave in France. Environ. Microbiol. 2012, 14, 3220–3231. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Cross section of Cueva de Ardales and locations of the aerobiological sampling points.

Figure 2.
Map of Cueva del Tesoro and sampling locations.

Figure 3.
Speleothems and cave walls abundantly coated by phototrophic biofilms in Cueva del Tesoro. (A) Sala del Aguila; (B) Sala de Lagos; (C) Sala de Marco Craso; (D–G) Sala de Lagos.
Figure 3.
Speleothems and cave walls abundantly coated by phototrophic biofilms in Cueva del Tesoro. (A) Sala del Aguila; (B) Sala de Lagos; (C) Sala de Marco Craso; (D–G) Sala de Lagos.

Figure 4.
Map of Gruta de las Maravillas and sampling locations.

Figure 5.
CFU/m3 of fungi in the four samplings. (A) Cueva de Ardales; (B) Cueva del Tesoro; (C) Gruta de las Maravillas.
Figure 5.
CFU/m3 of fungi in the four samplings. (A) Cueva de Ardales; (B) Cueva del Tesoro; (C) Gruta de las Maravillas.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dominguez-Moñino, I.; Jurado, V.; Rogerio-Candelera, M.A.; Hermosin, B.; Saiz-Jimenez, C. Airborne Fungi in Show Caves from Southern Spain. Appl. Sci. 2021, 11, 5027. https://0-doi-org.brum.beds.ac.uk/10.3390/app11115027
AMA Style
Dominguez-Moñino I, Jurado V, Rogerio-Candelera MA, Hermosin B, Saiz-Jimenez C. Airborne Fungi in Show Caves from Southern Spain. Applied Sciences. 2021; 11(11):5027. https://0-doi-org.brum.beds.ac.uk/10.3390/app11115027
Chicago/Turabian StyleDominguez-Moñino, Irene, Valme Jurado, Miguel Angel Rogerio-Candelera, Bernardo Hermosin, and Cesareo Saiz-Jimenez. 2021. "Airborne Fungi in Show Caves from Southern Spain" Applied Sciences 11, no. 11: 5027. https://0-doi-org.brum.beds.ac.uk/10.3390/app11115027
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.