
Keratinophilic and Keratinolytic Fungi in Cave Ecosystems: A Culture-Based Study of Brestovská Cave and Demänovská Ľadová and Slobody Caves (Slovakia)

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
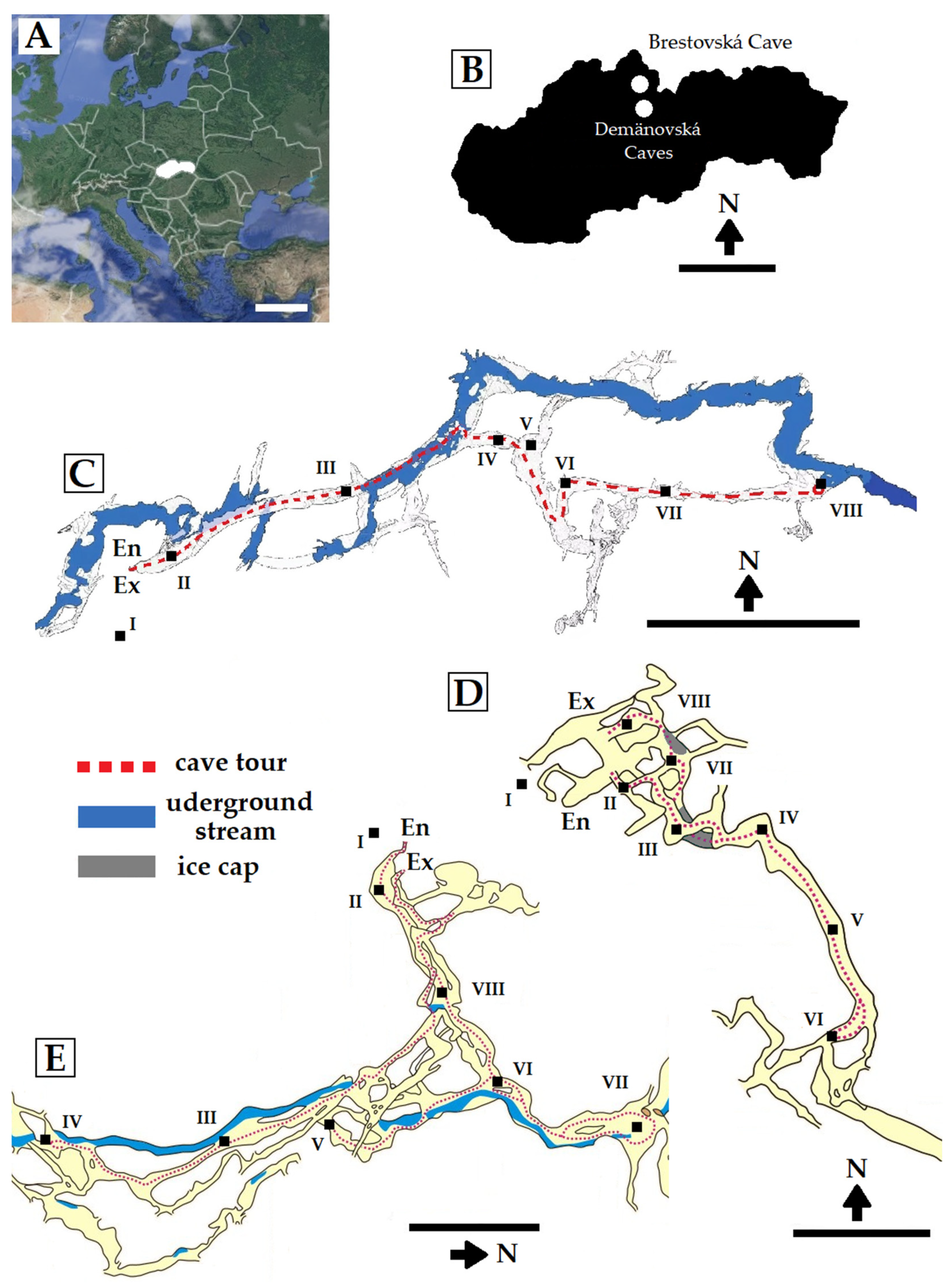
2.1. Study Area
2.2. Temperature Measurement
2.3. Sample Collection
2.4. Isolation of Fungi from Samples
2.5. Fungal Identification
2.6. Data Analyses
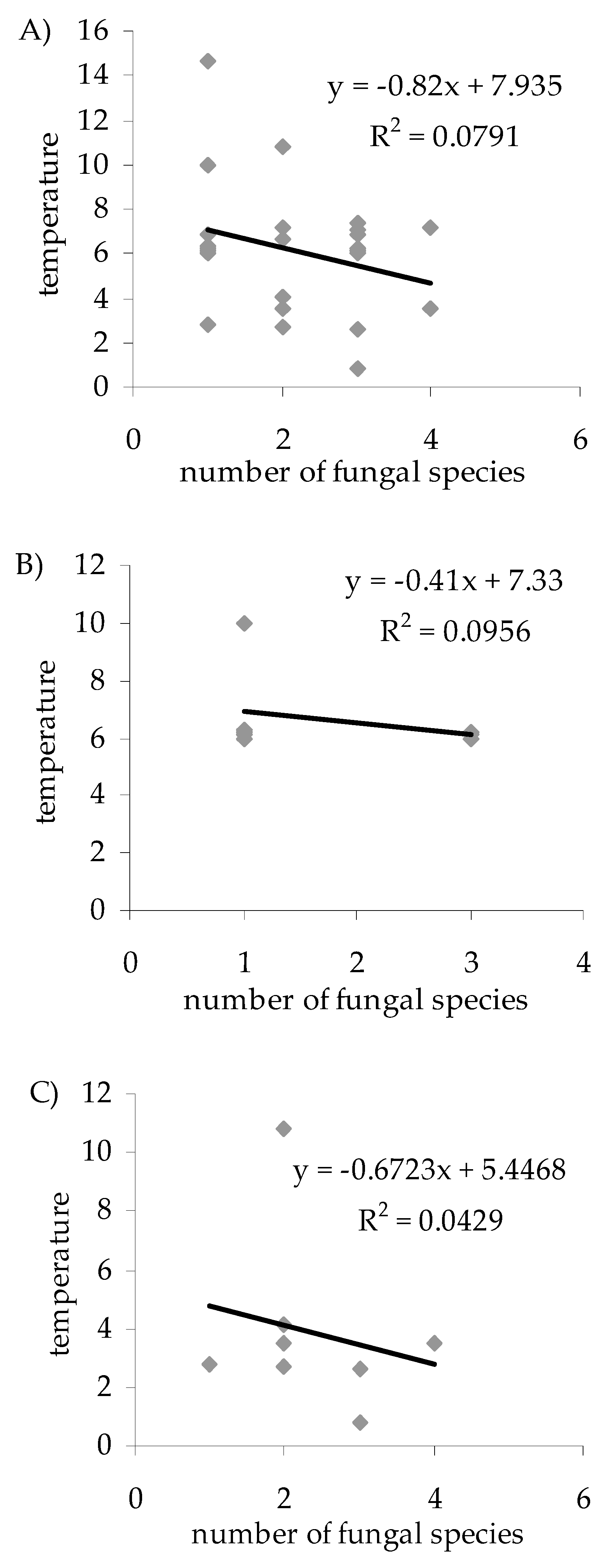
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Poulson, T.L.; White, W.B. The cave environment. Science 1969, 165, 971–981. [Google Scholar] [CrossRef] [PubMed]
- Barton, H.A.; Northup, D.E. Geomicrobiology in cave environments: Past current and future perspectives. J. Caves Karst Stud. 2007, 69, 163–178. [Google Scholar]
- Ogórek, R.; Višňovská, Z.; Tančinová, D. Mycobiota of underground habitats: Case study of Harmanecká Cave in Slovakia. Microb. Ecol. 2016, 71, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Ogórek, R.; Dyląg, M.; Višňovská, Z.; Tančinová, D.; Zalewski, D. Speleomycology of air and rock surfaces in Driny Cave (Lesser R. Carpathians, Slovakia). J. Cave Karst. Stud. 2016, 78, 119–127. [Google Scholar] [CrossRef]
- Ogórek, R.; Lejman, A.; Matkowski, K. Fungi isolated from Niedźwiedzia Cave in Kletno (Lower Silesia, Poland). Int. J. Speleol. 2013, 42, 161–166. [Google Scholar] [CrossRef] [Green Version]
- Kokurewicz, T.; Ogórek, R.; Pusz, W.; Matkowski, K. Bats increase the number of cultivable airborne fungi in the “Nietoperek” bat reserve in Western Poland. Microb. Ecol. 2016, 72, 36–48. [Google Scholar] [CrossRef] [Green Version]
- Cigna, A.A. Modern trend(s) in cave monitoring. Acta Carsologica 2002, 31, 35–54. [Google Scholar] [CrossRef]
- Snider, J.R.; Goin, C.; Miller, R.V.; Boston, P.J.; Northup, D.E. Ultraviolet radiation sensitivity in cave bacteria: Evidence of adaptation to the subsurface? Int. J. Speleol. 2009, 38, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Pflitsch, A.; Piasecki, J. Detection of an airflow system in Niedzwiedzia (Bear) Cave, Kletno, Poland. J. Cave Karst Stud. 2003, 65, 160–173. [Google Scholar]
- Pusz, W.; Ogórek, R.; Uklańska-Pusz, C.M.; Zagożdżon, P. Speleomycological research in underground Osówka complex in Sowie Mountains (Lower Silesia, Poland). Int. J. Speleol. 2014, 43, 27–34. [Google Scholar] [CrossRef] [Green Version]
- Ogórek, R.; Pusz, W.; Zagożdżon, P.P.; Kozak, B.; Bujak, H. Abundance and diversity of psychrotolerant cultivable mycobiota in winter of a former aluminous shale mine. Geomicrobiol. J. 2017, 34, 823–833. [Google Scholar] [CrossRef]
- Vanderwolf, K.J.; Malloch, D.; McAlpine, D.F.; Forbes, G.J. A world review of fungi, yeasts, and slime molds in caves. Int. J. Speleol. 2013, 42, 77–96. [Google Scholar] [CrossRef]
- Ogórek, R.; Pusz, W.; Lejman, A.; Uklańska-Pusz, C.M. Microclimate effects on number and distribution of fungi in the Włodarz underground complex in the Owl Mountains (Góry Sowie), Poland. J. Cave Karst Stud. 2014, 76, 146–153. [Google Scholar] [CrossRef]
- Nováková, A. Microscopic fungi isolated from the Domica Cave system (Slovak Karst National Park, Slovakia). A review. Int. J. Speleol. 2009, 38, 71–82. [Google Scholar] [CrossRef] [Green Version]
- Saiz-Jimenez, C. Microbiological and environmental issues in show caves. World J. Microbiol. Biotechnol. 2012, 28, 2453–2464. [Google Scholar] [CrossRef] [PubMed]
- Ogórek, R.; Speruda, M.; Borzęcka, J.; Piecuch, A.; Cal, M. First speleomycological study on the occurrence of psychrophilic and psychrotolerant aeromycota in the brestovská cave (Western Tatras Mts., Slovakia) and first reports for some species at underground sites. Biology 2021, 10, 497. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.C.; Wynn, P.M.; Barker, P.A. Natural and anthropogenic factors which influence aerosol distribution in Ingleborough Show Cave, UK. Int. J. Speleol. 2013, 42, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Domínguez-Villar, D.; Lojen, S.; Krklec, K. Is global warming affecting cave temperatures? Experimental and model data from aparadigmatic case study. Clim. Dyn. 2015, 45, 569–581. [Google Scholar] [CrossRef]
- Domínguez-Villar, D.; Fairchild, I.J.; Baker, A.; Carrasco, R.M.; Pedraza, J. Reconstruction of cave air temperature based on surface atmosphere temperature and vegetation changes: Implications for speleothem palaeoclimate records. Earth Planet. Sci. Lett. 2013, 369–370, 158–168. [Google Scholar] [CrossRef]
- Wigley, T.M.L.; Brown, M.C. Geophysical applications of heat and mass transfer in turbulent pipe flow. Bound.-Layer Meteorol. 1971, 1, 300–320. [Google Scholar] [CrossRef]
- Killing-Heinze, M.; Pflitsch, A.; Furian, W.; Allison, S. The of air temperature as a key parameter to identify climatic processes inside Carlsbad Cavern, New Mexico, USA. J. Cave Karst Stud. 2017, 79, 153–167. [Google Scholar] [CrossRef]
- Nadkarni, N.M.; Solano, R. Potential effects of climate change on canopy communities in a tropical cloud forest: An experimental approach. Oecologia 2002, 131, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Solache, M.A.; Casadevall, A. Global warming will bring new fungal diseases for mammals. MBio 2010, 1, e00061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurado, V.; Laiz, L.; Rodriguez-Nava, V.; Boiron, P.; Hermosin, H.; Sanchez-Moral, S.; Saiz-Jimenez, C. Pathogenic and opportunistic microorganisms in caves. Int. J. Speleol. 2010, 39, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Mulec, J. Human impact on underground cultural and natural heritage sites, biological parameters of monitoring and remediation actions for insensitive surfaces: Case of Slovenian show caves. J. Nat. Conserv. 2014, 22, 132–141. [Google Scholar] [CrossRef]
- Ogórek, R.; Dyląg, M.; Kozak, B. Dark stains on rock surfaces in Driny Cave (Little Carpathian Mountains, Slovakia). Extremophiles 2016, 20, 641–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogórek, R.; Dyląg, M.; Kozak, B.; Višňovská, Z.; Tančinová, D.; Lejman, A. Fungi isolated and quantified from bat guano and air in Harmanecká and Driny Caves (Slovakia). J. Cave Karst Stud. 2016, 78, 41–49. [Google Scholar] [CrossRef]
- Ogórek, R. Speleomycology of air in demänovská cave of liberty (Slovakia) and new airborne species for fungal sites. J. Cave Karst Stud. 2018, 80, 153–160. [Google Scholar] [CrossRef]
- Ogórek, R. Fungal Communities on rock surfaces in Demänovská Ice Cave and Demänovská Cave of Liberty (Slovakia). Geomicrobiol. J. 2018, 35, 266–276. [Google Scholar] [CrossRef]
- Ogórek, R.; Kozak, B.; Višňovská, Z.; Tančinová, D. Phenotypic and genotypic diversity of airborne fungal spores in Demänovská Ice Cave (Low Tatras, Slovakia). Aerobiologia 2018, 34, 13–28. [Google Scholar] [CrossRef] [Green Version]
- Ogórek, R.; Piecuch, A.; Višňovská, Z.; Cal, M.; Niedźwiecka, K. First report on the occurence of dermatophytes of Microsporum cookei clade and cose affinities to Paraphyton cookei in the Harmanecká Cave (Veľká Fatra Mts., Slovakia). Diversity 2019, 11, 191. [Google Scholar] [CrossRef] [Green Version]
- Simpanya, M.F. Dermatophytes: Their taxonomy, ecology and pathogenicity. Rev. Iberoam Micol. 2000, 669, 1–12. [Google Scholar]
- Gherbawy, Y.A.M.H. Keratinolytic and keratinophilic fungi of mangrove’s soil and air in the city of Qenaand their response to garlic extract and onion oil treatments. Acta Mycol. 1996, 31, 87–89. [Google Scholar] [CrossRef] [Green Version]
- Nudziková, Ľ. Vývoj návštevnosti sprístupnených jaskýň na Slovensku od roku 2009 (Course of show caves attendance in Slovakia since 2009). Aragonit 2014, 19, 35–38. (In Slovak) [Google Scholar]
- Droppa, A. Karst on Sivývrch. Československý Kras 1972, 23, 77–98. (In Slovak) [Google Scholar]
- Brestovská Cave. Slovak Caves Administration. Available online: https://www.ssj.sk (accessed on 2 December 2021).
- Marušin, M. Geological conditions—Factor of origin of two different cave systems in two adjacent valleys (the Demänovská Valley and the Jánska Valley, the Low Tatras, Slovakia). Acta Carsol. 2003, 32, 121–130. [Google Scholar]
- Piasecki, J.; Sawiński, T.; Strug, K.; Zelnika, J. Selected characteristics of the microclimate of the Demänovská Ice Cave (Slovakia). In Proceedings of the 2nd International Workshop on Ice Caves; Slovak Caves Administration: Demänovská Dolina, Slovakia, 2006; pp. 50–61. [Google Scholar]
- Demänovská Ice Cave. Slovak Caves Administration. Available online: http://www.ssj.sk/en/jaskyna/4-demanovska-cave-of-liberty (accessed on 2 December 2021).
- Demänovská Cave of Liberty. Slovak Caves Administration. Available online: http://www.ssj.sk (accessed on 2 December 2021).
- Vanbreuseghem, R. Technique biologique pour I’ isolement des dermat ophytes dusol. Ann. Soc. Belge. Med. Trop. 1952, 32, 173–178. [Google Scholar]
- Kuehn, H.H.; Orr, G.F. Arachniotus ruber (Van Tieghem) Schroeter. Trans. Brit. Mycol. Soc. 1964, 47, 553–558. [Google Scholar] [CrossRef]
- Farley, J.F.; Jersild, R.A.; Niederpruem, D.J. Ultrastructural aspects of ascosporulation in Arthroderma quadrifidum (=Trichophyton terrestre). Sabouraudia 1976, 14, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Chamuris, G.P. Nomenclatural adjustments in Stereum and Cylindrobasidium according to the Sydney Code. Mycotaxon 1984, 20, 587–588. [Google Scholar]
- Roux, C.; Van Warmelo, K.T. Conidiomata in Bartalinia robillardoides. Mycol. Res. 1990, 94, 109–116. [Google Scholar] [CrossRef]
- Weitzman, I.; Summerbell, R. The Dermatophytes. Clin. Microbiol. Rev. 1995, 8, 240–259. [Google Scholar] [CrossRef] [PubMed]
- Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of Penicillium subgenus. Penicillium a guide to identification of food and air-borne terverticillate Penicillia and their mycotoxins. Stud. Mycol. 2004, 49, 1–174. [Google Scholar]
- Heuchert, B.; Braun, U.; Schubert, K. Morphotaxonomic revision of fungicolous Cladosporium species (hyphomycetes). Schlechtendalia 2005, 13, 1–78. [Google Scholar]
- Lakshmipathy, D.T.; Kannabiran, K. Review on dermatomycosis: Pathogenesis and treatment. Nat. Sci. 2010, 2, 726–731. [Google Scholar] [CrossRef] [Green Version]
- Krzyściak, P.; Skóra, M.; Macura, A.B. Atlas Grzybów Chorobotwórczych Człowieka; MedPharm Polska: Wrocław, Poland, 2011; pp. 12–380. (In Polish) [Google Scholar]
- Jurjevic, Z.; Peterson, S.W.; Horn, B.W. Aspergillus section Versicolores: Nine new species and multilocus DNA sequence based phylogeny. IMA Fungus 2012, 3, 59–79. [Google Scholar] [CrossRef]
- Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Hong, S.-B.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Varga, J.; Yaguchi, T.; Samson, R.A. Identification and nomenclature of the genus Penicillium. Stud. Mycol. 2014, 78, 343–371. [Google Scholar] [CrossRef] [Green Version]
- Tsuji, M.; Tsujimoto, M.; Imura, S. Cystobasidium tubakii and Cystobasidium ongulense, new basidiomycetous yeast species isolated from East Ongul Island, East Antarctica. Mycoscience 2016, 58, 103–110. [Google Scholar] [CrossRef]
- Nguyen, T.T.T.; Lee, S.H.; Jeon, S.J.; Lee, H.B. First Records of Rare Ascomycete Fungi, Acrostalagmus luteoalbus, Bartalinia robillardoides, and Collariella carteri from Freshwater Samples in Korea. Mycobiology 2019, 47, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Dyląg, M.; Sawicki, A.; Ogórek, R. Diversity of species and susceptibility phenotypes toward commercially available fungicides of cultivable fungi colonizing bones of Ursus spelaeus on display in Niedźwiedzia Cave (Kletno, Poland). Diversity 2019, 11, 224. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Spellerberg, I.F.; Fedor, P. A tribute to Claude Shannon (1916–2001) and a plea for more rigorous use of species richness, species diversity and the ‘Shannon–Wiener’ Index. Glob. Ecol. Biogeogr. 2003, 12, 177–179. [Google Scholar] [CrossRef] [Green Version]
- Shannon, C.E.; Wiener, W. The Mathematical Theory of Communication; University Illinois Press: Urbana, IL, USA, 1963; p. 360. [Google Scholar]
- Lücking, R.; Aime, M.C.; Robbertse, B.; Miller, A.N.; Ariyawansa, H.A.; Aoki, T.; Cardinali, G.; Crous, P.W.; Druzhinina, I.S.; Geiser, D.M.; et al. Unambiguous identification of fungi: Where do we stand and how accurate and precise is fungal DNA barcoding? IMA Fungus 2020, 11, 14. [Google Scholar] [CrossRef] [PubMed]
- De Hoog, G.S.; Dukik, K.; Monod, M.; Packeu, A.; Stubbe, D.; Hendrickx, M.; Kupsch, C.; Stielow, J.B.; Freeke, J.; Göker, M.; et al. Toward a novel multilocus phylogenetic taxonomy for the dermatophytes. Mycopathologia 2017, 182, 5–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchisio, F.V.; Curetti, D.; Cassinelli, C.; Bordese, C. Keratinolytic and keratinophilic fungi in the soils of Papua New Guinea. Mycopathologia 1991, 115, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Ulfig, K. Studies of keratinolytic and keratinophilic fungi in sewage sludge by means of a multi-temperature hair baiting method. Pol. J. Environ. Stud. 2003, 12, 461–466. [Google Scholar]
- Korniłłowicz-Kowalska, T.; Bohacz, J. Biodegradation of keratin waste: Theory and practical aspects. Waste Manag. 2011, 31, 1689–1701. [Google Scholar] [CrossRef]
- Abdel-Fattah, H.M.; Moubasher, A.H.; Maghazy, S.M. Keratinolytic fungi in Egyptian soils. Microbiol. Immunol. 1982, 26, 177–180. [Google Scholar] [CrossRef] [Green Version]
- Korniłłowicz-Kowalska, T.; Bohacz, J. Some correlations between the occurrence frequency of keratinophilic fungi and selected soil properties. Acta Mycol. 2002, 37, 101–116. [Google Scholar] [CrossRef] [Green Version]
- Bohacz, J.; Korniłłowicz-Kowalska, T. Species diversity of keratinophilic fungi in various soil types. Cent. Eur. J. Biol. 2012, 7, 259–266. [Google Scholar] [CrossRef]
- Bohacz, J. Biodegradation of feather waste keratin by a keratinolytic soil fungus of the genus Chrysosporium and statistical optimization of feather mass loss. World J. Microb. Biot. 2017, 33, 13. [Google Scholar] [CrossRef] [Green Version]
- Ulfig, K.; Terakowski, M.; Płaza, G.; Kosarewicz, O. Keratinolytic fungi in sewage sludge. Mycopathologia 1996, 136, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Ulfig, K.; Guarro, J.; Cano, J.; Gené, J.; Vidal, P.; Figueras, M.J.; Łukasik, W. The occurrence of keratinolytic fungi in sediments of the river Tordera (Spain). FEMS Microbiol. Ecol. 1997, 22, 111–117. [Google Scholar] [CrossRef]
- Cascarosa, E.; Gea, G.; Arauzo, J. Thermochemical processing of meat and bone meal: A review. Renew. Sustain. Energy Rev. 2012, 16, 942–957. [Google Scholar] [CrossRef]
- Moorthy, K.; Prasanna, I.; Vimalan, S.; Lavanya, V.; Thamarai Selvi, A.; Mekala, T.; Thajuddin, N. Study on keratinophilic and keratinolytic fungi isolated from birds’ feathers and animal hairs. Biosci. Biotechnol. Res. Asia 2011, 8, 633–640. [Google Scholar] [CrossRef]
- Hubálek, Z. Keratinophilic fungi associated with free-living mammals and birds. In Biology of Dermatophytes and Other Keratinophilic Fungi, 1st ed.; Kushwaha, R.K.S., Guarro, J., Eds.; Revista Iberoamericana de Micología: Bilbao, Spain, 2000; pp. 86–92. [Google Scholar]
- Korniłłowicz-Kowalska, T.; Kitowski, I. Nests of Marsh harrier (Circus aeruginosus L.) as refuges of potentially phytopathogenic and zoopathogenic fungi. Saudi J. Biol. Sci. 2018, 25, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Ciesielska, A.; Korniłłowicz-Kowalska, T.; Kitowski, I.; Bohacz, J. The dispersal of rodent-borne strains of Aphanoascus keratinophilus and Chrysosporium tropicum by pellets of predatory birds. Avian Biol. Res. 2017, 10, 218–230. [Google Scholar] [CrossRef]
- Błyskal, B. Fungi utilizing keratinous substrates. Int. Biodeterior. Biodegrad. 2009, 63, 631–653. [Google Scholar] [CrossRef]
- Kunert, J. Physiology of keratinophilic fungi. In Biology of Dermatophytes and Other Keratinophilic Fungi, 1st ed.; Kushwaha, R.K.S., Guarro, J., Eds.; Revista Iberoamericana de Micología: Bilbao, Spain, 2000; pp. 77–85. [Google Scholar]
- Mitola, G.; Escalona, F.; Salas, R.; Garcia, E.; Ledesma, A. Morphological characterization of in-vitro human hair keratinolysis, produced by identified wild strains of Chrysosporium species. Mycopathologia 2002, 156, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Călin, M.; Constantinescu-Aruxandei, D.; Alexandrescu, E.; Răut, I.; Badea Doni, M.; Arsene, M.L.; Oancea, F.; Jecu, L.; Lazărb, V. Degradation of keratin substrates by keratinolytic fungi. Electron. J. Biotechnol. 2017, 28, 101–112. [Google Scholar] [CrossRef]
- Sutoyo, S.; Subandi; Ardyati, T.; Suharjono, I. Screening of keratinolytic fungi for biodegradation agent of keratin from chicken feather waste. Annu. Conf. Environ. Sci. Soc. Appl. 2019, 391, 012027. [Google Scholar] [CrossRef]
- Raudabaugh, D.B.; Miller, A.N. Nutritional capability of and substrate suitability for Pseudogymnoascus destructans, the causal agent of bat white-nose syndrome. PLoS ONE 2013, 8, e78300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumaresan, D.; Wischer, D.; Stephenson, J.; Hillebrand-Voiculescu, A.; Murrell, J.C. Microbiology of Movile Cave—Chemolithoautotrophic ecosystem. Geomicrobiol. J. 2014, 31, 186–193. [Google Scholar] [CrossRef]
- Vanderwolf, K.J.; Malloch, D.; Ivanova, V.N.; McAlpine, F.D. Lack of cave-associated mammals influences the fungal assemblages of in- sular solution caves in eastern Canada. J. Cave Karst Stud. 2016, 78, 198–207. [Google Scholar] [CrossRef]
- Held, B.W.; Salomon, C.E.; Blanchette, R.A. Diverse subterranean fungi of an underground iron ore mine. PLoS ONE 2020, 15, e023420. [Google Scholar] [CrossRef]
- Evolceanu, R.; Alteraş, I. Eine keratinophile Chrysosporium-Art mit ausgesprochen dermatophytischen, immunbiologischen Eigenschaften aus Guano von einer Grotte in Rumänien (unvollkommenes Stadium von Arthroderma multifidum-Dawson 1963?) (Erste) [A keratinophilic strain of Chrysosporium with outspoken dermatophytic, immunobiologic properties from guano in a cave in Rumania (imperfect stage of Arthroderma multifidum-Dawson 1963?) I]. Mycoses 1967, 10, 489–492. [Google Scholar]
- Lurie, H.I.; Borok, R. Trichophyton mentagrophytes isolated from the soil of caves. Mycologia 1955, 47, 506–510. [Google Scholar] [CrossRef]
- Lurie, H.I.; Way, M. The isolation of dermatophytes from the atmosphere of caves. Mycologia 1957, 49, 178–180. [Google Scholar] [CrossRef]
- Kajihiro, E.S. Occurrence of dermatophytes in fresh bat guano. Appl. Microbiol. 1965, 13, 720–724. [Google Scholar] [CrossRef]
- Balabanoff, V.A. Comparative studies of dermatophytes isolated from caves and stables in Bulgaria. Mycopathol. Mycol. Appl. 1967, 32, 237–248. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Liu, F.; Zhou, X.; Liu, X.Z.; Liu, S.J.; Cai, L. Culturable mycobiota from Karst caves in China, with descriptions of 20 new species. Persoonia 2017, 39, 1–31. [Google Scholar] [CrossRef]
- Brasch, J.; Gräser, Y. Trichophyton eboreum sp. nov. isolated from human skin. J. Clin. Microbiol. 2005, 43, 5230–5237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, C.K.; Borman, A.M.; Linton, C.J.; Bridge, P.D.; Johnson, E.M. Arthroderma olidum, sp. nov. A new addition to the Trichophyton terrestre complex. Med. Mycol. 2006, 44, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Brasch, J.; Gräser, Y. Trichophyton eboreum—Ein kürzlich beschriebener Dermatophyt. J. Dtsch. Dermatol. 2006, 4, 646–649. [Google Scholar] [CrossRef] [PubMed]
- Orlando, G.; Adorisio, S.; Delfino, D.; Chiavaroli, A.; Brunetti, L.; Recinella, L.; Leone, S.; D’Antonio, M.; Zengin, G.; Acquaviva, A.; et al. Comparative investigation of composition, antifungal, and anti-Inflammatory effects of the essential oil from three industrial hemp varieties from Italian cultivation. Antibiotics 2021, 10, 334. [Google Scholar] [CrossRef] [PubMed]
- Fowora, M.; Onyeaghasiri, F.; Olanlege, A.; Edu-Muyideen, I.; Adebesin, O. In Vitro susceptibility of dermatophytes to anti-fungal drugs and aqueous Acacia nilotica leaf extract in Lagos, Nigeria. J. Biomed. Sci. Eng. 2021, 14, 74–82. [Google Scholar] [CrossRef]
- Labuda, R.; Bernreiter, A.; Hochenauer, D.; Kubátová, A.; Kandemir, H.; Schüller, C. Molecular systematics of Keratinophyton: The inclusion of species formerly referred to Chrysosporium and description of four new species. IMA Fungus 2021, 12, e17. [Google Scholar] [CrossRef]
- Labuda, R.; Naďová, L.; Vén, T. First record of Chrysosporium europae, Ch. fluviale and Ch. minutisporosum in Slovakia. Biologia 2008, 63, 38–39. [Google Scholar] [CrossRef]
- Kochkina, G.; Ivanushkina, N.; Ozerskaya, S.; Chigineva, N.; Vasilenko, O.; Firsov, S.; Spirina, E.; Gilichinsky, D. Ancient fungi in Antarctic permafrost environments. FEMS Microbiol. Ecol. 2012, 82, 501–509. [Google Scholar] [CrossRef] [Green Version]
- Ulfig, K. The occurrence of keratinolytic fungi in waste and waste-contaminated habitats. Rev. Iberoam. Micol. 2000, 17, 44–49. [Google Scholar]
- Batra, A.; Saxena, R.K. Potential tannase producers from the genera Aspergillus and Penicillium. Process Biochem. 2005, 40, 1553–1557. [Google Scholar] [CrossRef]
- Abbas, C.A.; Groves, S.; Gander, J.E. Isolation, purification, and properties of Penicillium charlesii alkaline protease. J. Bacteriol. 1989, 171, 5630–5637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ondeyka, J.G.; Goegelman, R.T.; Schaeffer, J.M.; Kelemen, L.; Zitano, L. Novel antinematodal and antiparasitic agents from Penicillium charlesii. I. Fermentation, isolation and biological activity. J. Antibiot. 1990, 43, 1375–1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendell, M.J.; Macher, J.M.; Kumagai, K. Measured moisture in buildings and adverse health effects, A review. Indoor Air 2018, 28, 488–499. [Google Scholar] [CrossRef] [PubMed]
- Bastian, F.; Alabouvette, C.; Saiz-Jimenez, C. The impact of arthropods on fungal community structure in Lascaux Cave. J. Appl. Microbiol. 2009, 106, 1456–1462. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Ma, X.; Ma, Y.; Mao, L.; Wu, F.; Ma, X.; An, L.; Feng, H. Seasonal dynamics of airborne fungi in different caves of the Mogao Grottoes, Dunhuang, China. Int. Biodeterior. Biodegradation 2010, 64, 461–466. [Google Scholar] [CrossRef]
- Borzęcka, J.; Piecuch, A.; Kokurewicz, T.; Lavoie, K.H.; Ogórek, R. Greater Mouse-Eared Bats (Myotis myotis) hibernating in the Nietoperek bat reserve (Poland) as a vector of airborne culturable fungi. Biology 2021, 10, 593. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Sites 1 | Temperature of Soil/Sediment (°C) | |||||
|---|---|---|---|---|---|---|
| Brestovská Cave | Demänovská Ľadová Cave | Demänovská Slobody Cave | ||||
| I | 10.0 | a 2 | 10.8 | a | 14.7 | a |
| II | 6.3 | b | 3.5 | c | 7.2 | c |
| III | 6.2 | bc | 2.6 | d | 6.9 | d |
| IV | 6.1 | bc | 3.5 | c | 6.9 | d |
| V | 6.1 | bc | 2.7 | d | 7.1 | c |
| VI | 6.0 | c | 2.8 | d | 7.4 | b |
| VII | 6.0 | c | 0.8 | e | 7.2 | c |
| VIII | 6.2 | bc | 4.1 | b | 6.7 | e |
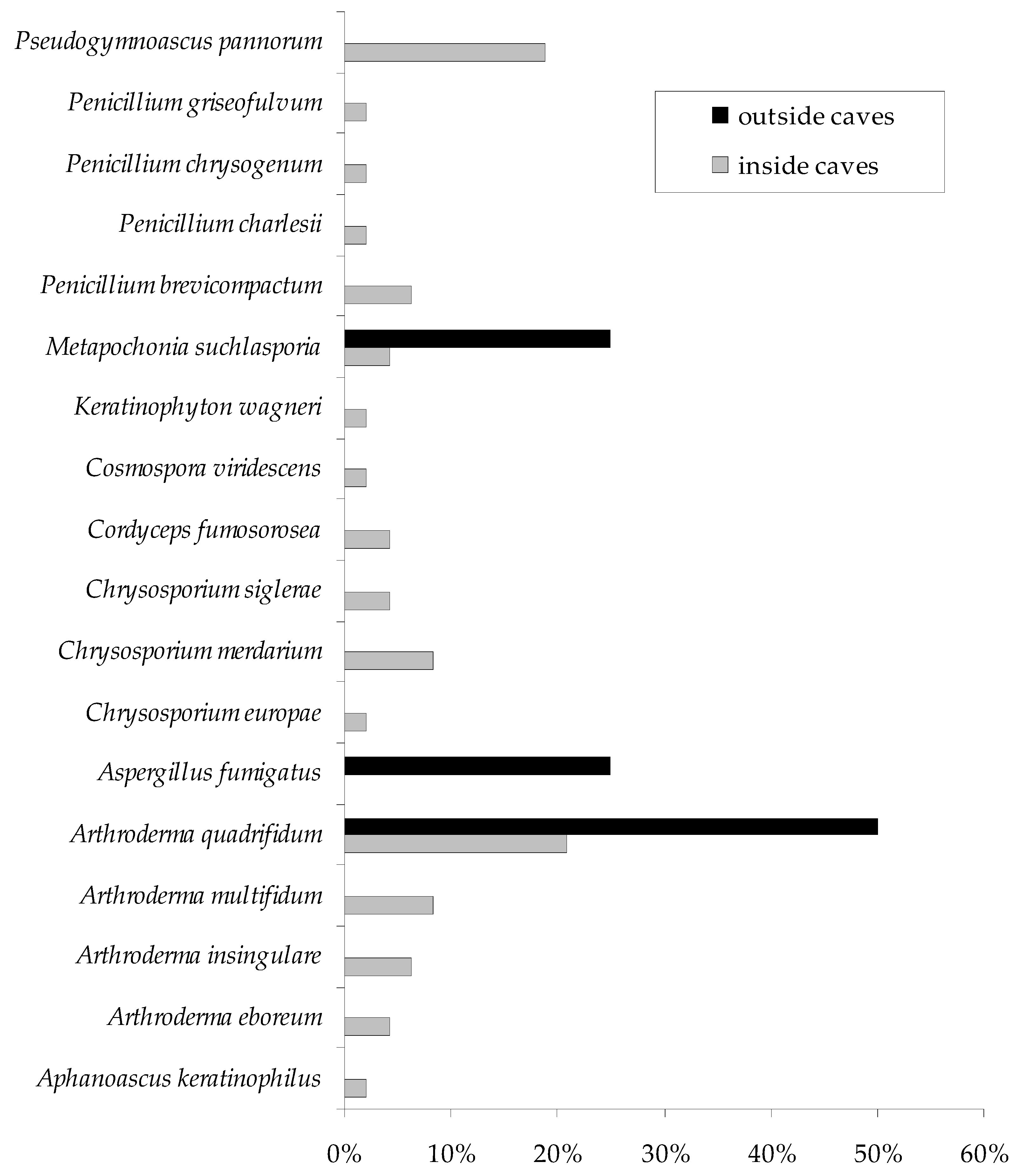
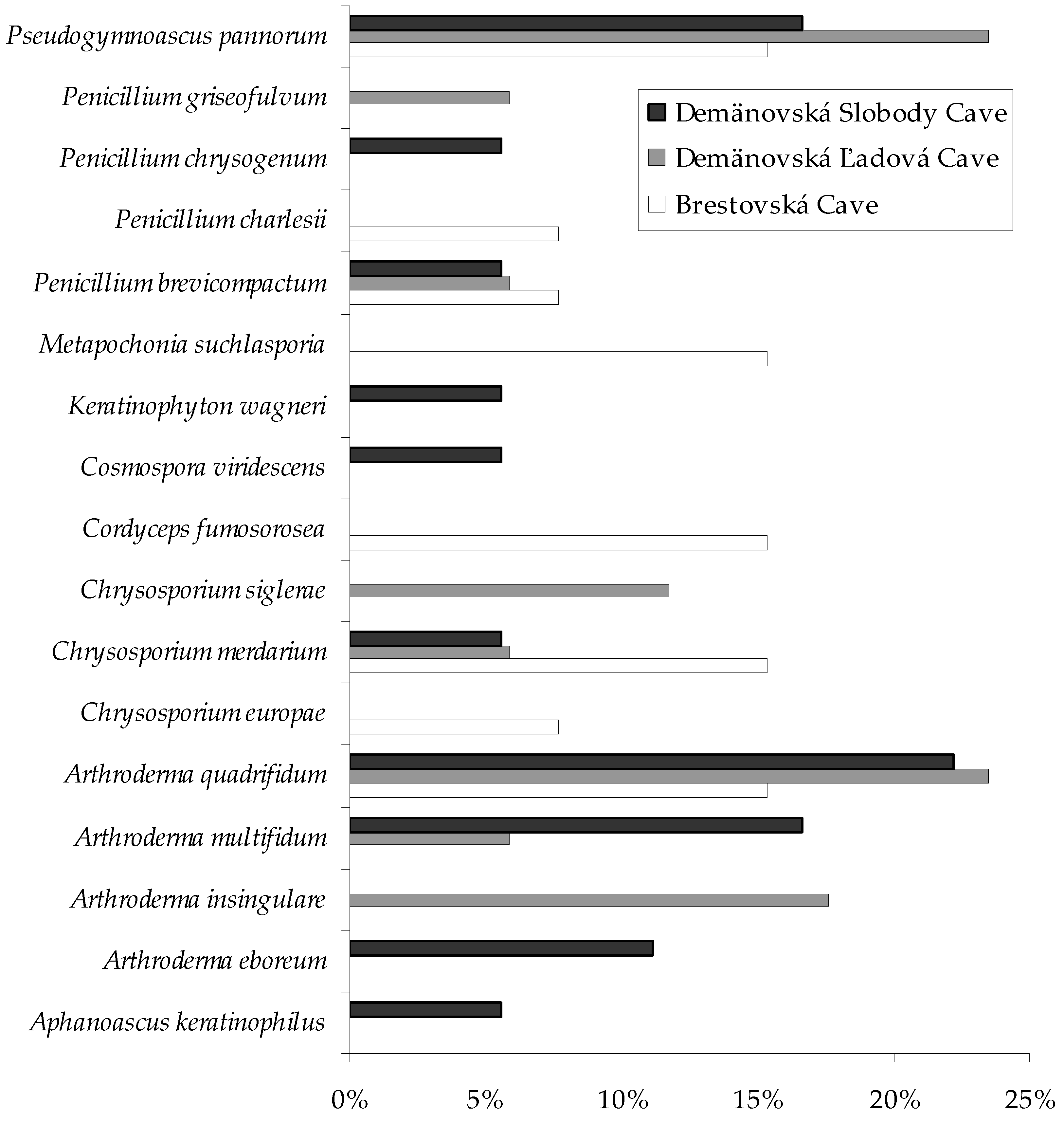
| Fungi | Study Sites 1 | |
|---|---|---|
| Family | Species | |
| Arthrodermataceae | Arthroderma eboreum | V 2 S 3, VIII S |
| Arthroderma insingulare | IV L, VII L, VIII L | |
| Arthroderma multifidum | VII L, II S, III S, IV S | |
| Arthroderma quadrifidum | I B, III B, VII B, III L, IV L, V L, VII L, I S, II S, III S, VII S, VIII S | |
| Aspergillaceae | Aspergillus fumigatus | I L |
| Penicillium brevicompactum | VII B, IV L, VI S | |
| Penicillium charlesii | V B | |
| Penicillium chrysogenum | VII S | |
| Penicillium griseofulvum | II L | |
| Clavicipitaceae | Metapochonia suchlasporia | II B, III B, I L |
| Cordycipitaceae | Cordyceps fumosorosea | III B, VIII B |
| Nectriaceae | Cosmospora viridescens | VII S |
| Onygenaceae | Aphanoascus keratinophilus | VI S |
| Chrysosporium europae | V B | |
| Chrysosporium merdarium | IV B, VI B, III L, V S | |
| Chrysosporium siglerae | II L, VII L | |
| Keratinophyton wagneri | VII S | |
| Pseudeurotiaceae | Pseudogymnoascus pannorum | V B, VII B, III L, IV L, V L, VIII L, III S, V S, VI S |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogórek, R.; Suchodolski, J.; Piecuch, A.; Przywara, K.; Višňovská, Z. Keratinophilic and Keratinolytic Fungi in Cave Ecosystems: A Culture-Based Study of Brestovská Cave and Demänovská Ľadová and Slobody Caves (Slovakia). Appl. Sci. 2022, 12, 1455. https://0-doi-org.brum.beds.ac.uk/10.3390/app12031455
Ogórek R, Suchodolski J, Piecuch A, Przywara K, Višňovská Z. Keratinophilic and Keratinolytic Fungi in Cave Ecosystems: A Culture-Based Study of Brestovská Cave and Demänovská Ľadová and Slobody Caves (Slovakia). Applied Sciences. 2022; 12(3):1455. https://0-doi-org.brum.beds.ac.uk/10.3390/app12031455
Chicago/Turabian StyleOgórek, Rafał, Jakub Suchodolski, Agata Piecuch, Katarzyna Przywara, and Zuzana Višňovská. 2022. "Keratinophilic and Keratinolytic Fungi in Cave Ecosystems: A Culture-Based Study of Brestovská Cave and Demänovská Ľadová and Slobody Caves (Slovakia)" Applied Sciences 12, no. 3: 1455. https://0-doi-org.brum.beds.ac.uk/10.3390/app12031455