
Signaling Pathways Associated with Macrophage-Activating Polysaccharide Isolated from Korea Red Ginseng


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Extraction and Purification of KRG-P
2.3. Cell Culture
2.4. Cytotoxicity Assay
2.5. Cytokine and NO Production
2.6. Immunoblot Analysis
2.7. Real-Time PCR
2.8. Statistical Analysis
3. Results
3.1. Chemical Properties of Polysaccharide (KRG-P) Isolated from KRG
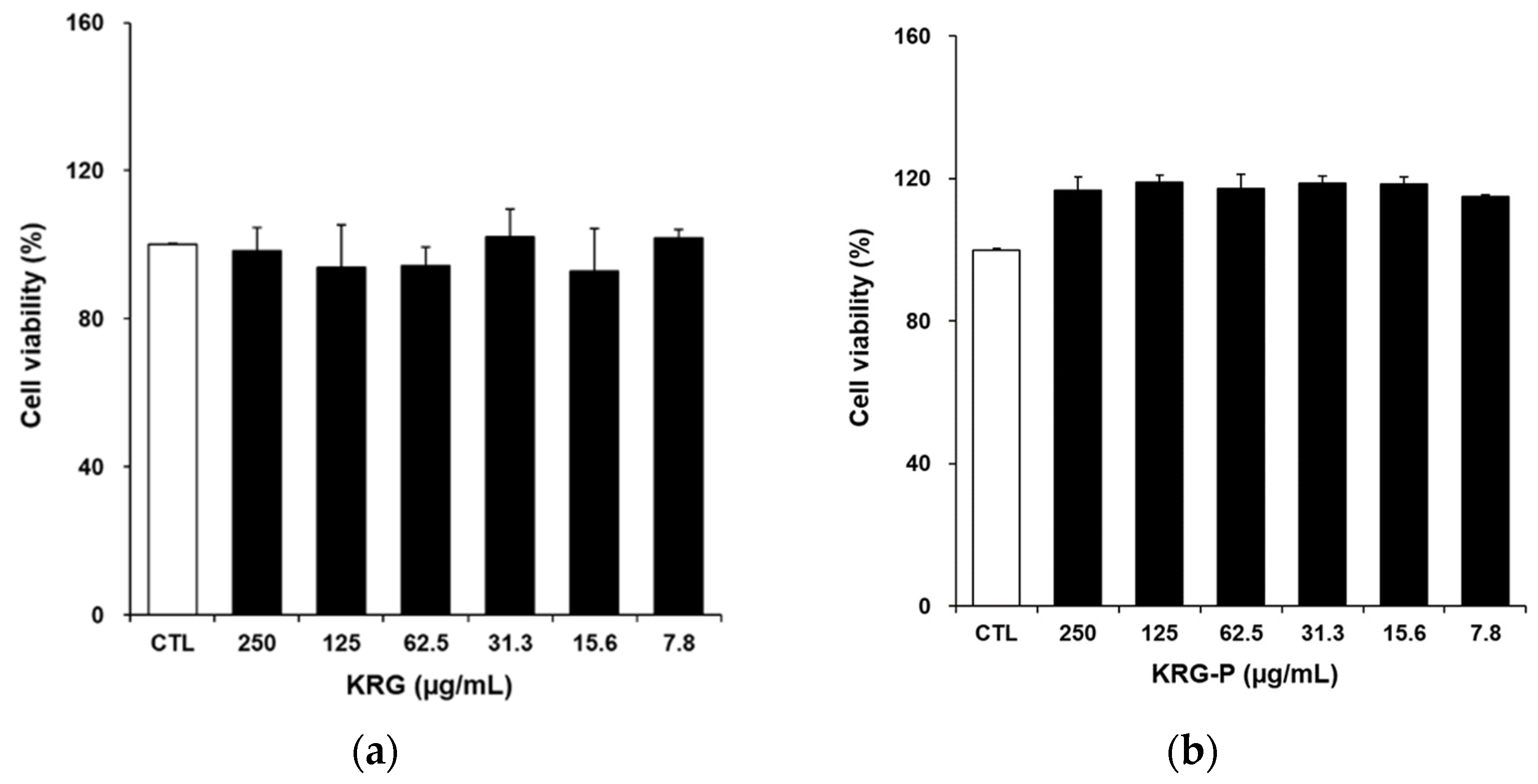
3.2. Cytotoxicity Assay of KRG and KRG-P
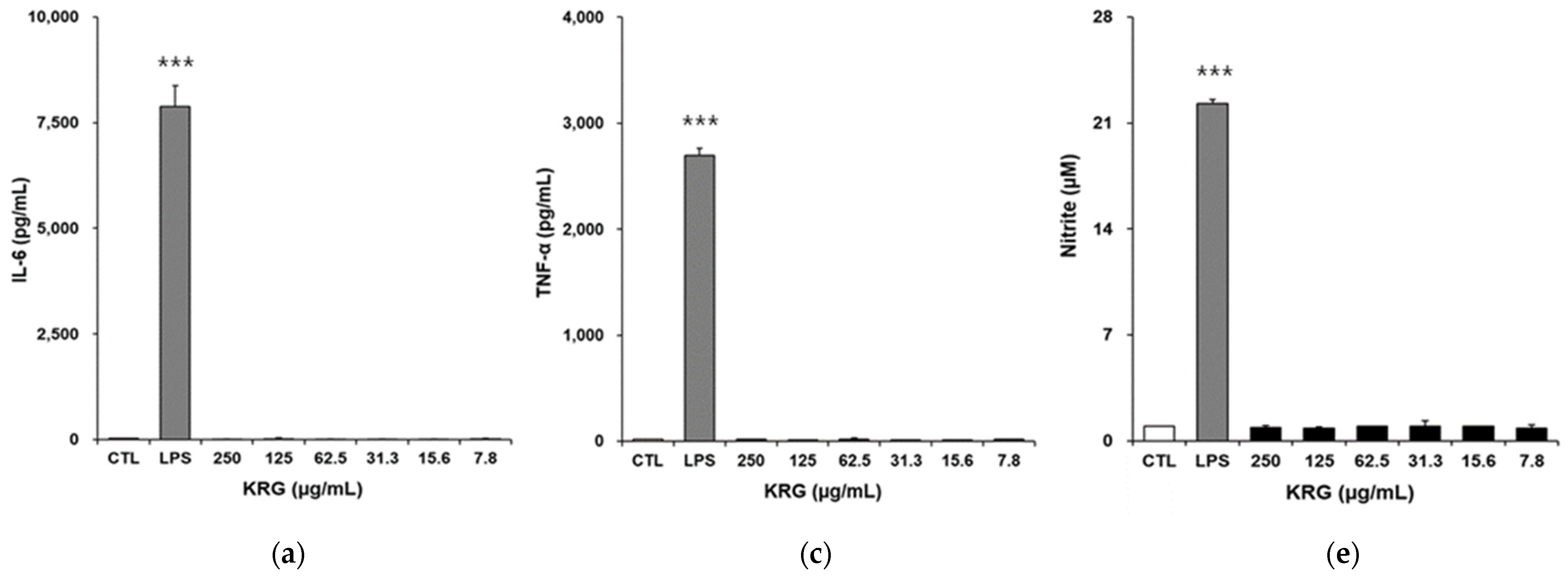
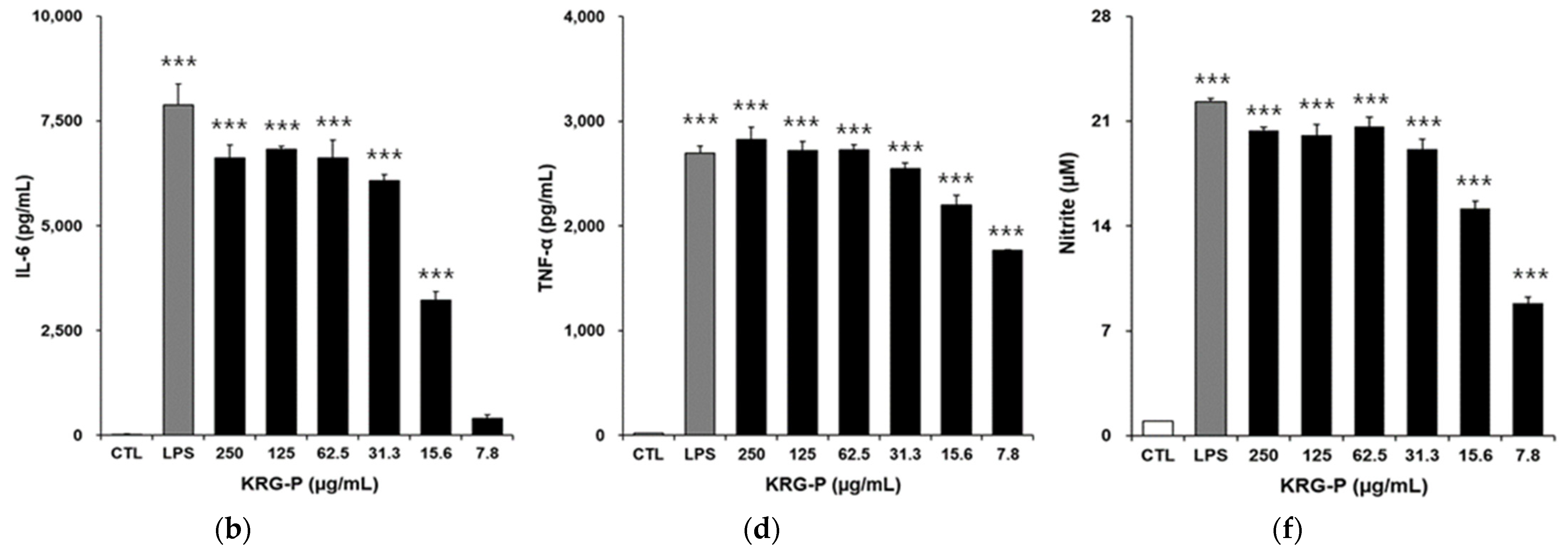
3.3. Effects of KRG and KRG-P on IL-6, TNF-α, and NO Production in RAW264.7 Cells
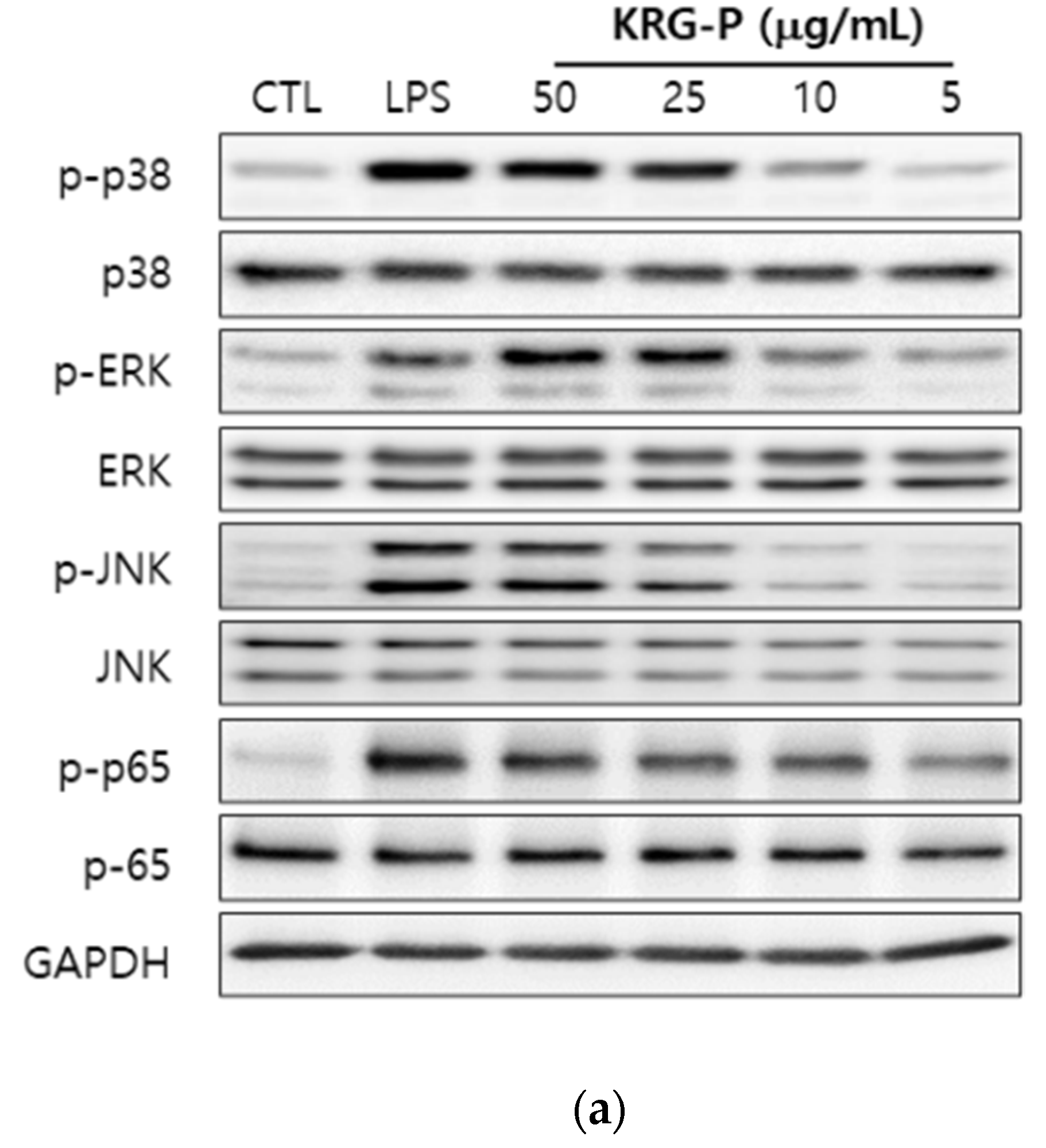
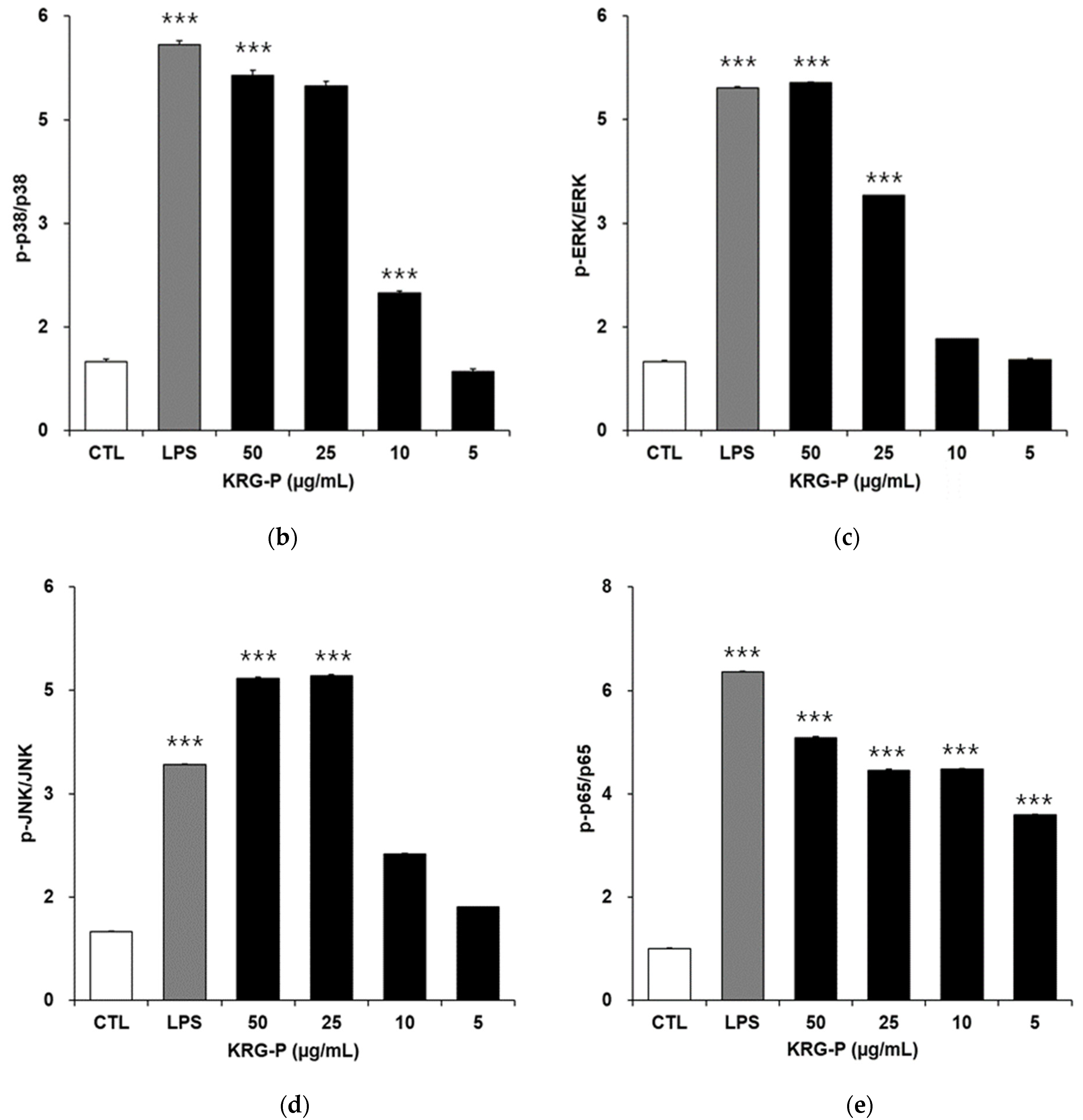
3.4. Effects of KRG-P on MAPKs and NF-κB Phosphorylation in RAW264.7 Cells
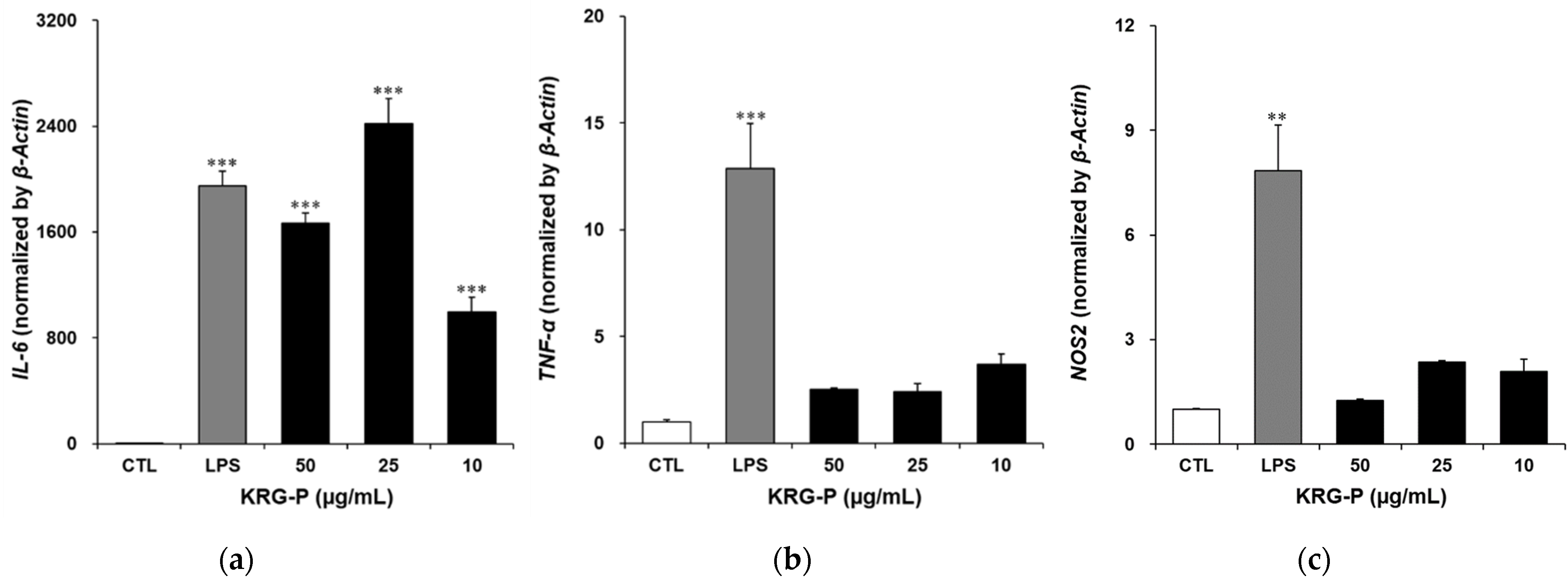
3.5. Effects of KRG-P on IL-6, TNF-α, and NOS2 Gene Expression in RAW264.7 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Baek, K.S.; Hong, Y.D.; Kim, Y.; Sung, N.Y.; Yang, S.; Lee, K.M.; Park, J.Y.; Park, J.S.; Rho, H.S.; Shin, S.S.; et al. Anti-inflammatory activity of AP-SF, a ginsenoside-enriched fraction, from Korean ginseng. J. Ginseng Res. 2015, 39, 155–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, B.H.; Park, M.H.; Woo, L.K.; Woo, W.S.; Han, Y.N. Studies on the antioxidant components of Korean ginseng. In Proceedings of the Ginseng Society Conference 1978, Seoul, Korea, 7–11 September 1978; pp. 13–17. [Google Scholar]
- Wong, A.S.; Che, C.M.; Leung, K.W. Recent advances in ginseng as cancer therapeutics: A functional and mechanistic overview. Nat. Prod. Rep. 2015, 32, 256–272. [Google Scholar] [CrossRef]
- Shibata, S. Chemistry and cancer preventing activities of ginseng saponins and some related triterpenoid compounds. J. Korean Med. Sci. 2001, 16, S28–S37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.H.; Jung, B.H.; Kim, S.Y.; Lee, E.H.; Chung, B.C. The antistress effect of ginseng total saponin and ginsenoside Rg3 and Rb1 evaluated by brain polyamine level under immobilization stress. Pharmacol. Res. 2006, 54, 46–49. [Google Scholar] [CrossRef]
- Kennedy, D.O.; Scholey, A.B. Ginseng: Potential for the enhancement of cognitive performance and mood. Pharmacol. Biochem. Behav. 2003, 75, 687–700. [Google Scholar] [CrossRef]
- Wang, J.; Li, S.; Fan, Y.; Chen, Y.; Liu, D.; Cheng, H.; Gao, X.; Zhou, Y. Anti-fatigue activity of the water-soluble polysaccharides isolated from panax ginseng CA Meyer. J. Ethnopharmacol. 2010, 130, 421–423. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Kwon, R.H.; Jang, M.W.; Ha, B.J. Antioxidant and anti-wrinkle effects of steamed three ginseng extracts. J. Soc. Cosmet. Sci. Korea 2012, 38, 155–162. [Google Scholar]
- Lee, J.H.; Cho, S.H.; Yun, M.Y.; An, S.; Jang, H.H.; Lee, S.N.; Song, G.Y. Anti-wrinkle effect of rare ginsenosides, produced from ginsenoside Rd. Asian J. Beauty Cosmetol. 2015, 13, 909–916. [Google Scholar]
- Attele, A.S.; Wu, J.A.; Yuan, C.S. Ginseng pharmacology: Multiple constituents and multiple actions. Biochem. Pharmacol. 1999, 58, 1685–1693. [Google Scholar] [CrossRef]
- Christensen, L.P. Ginsenosides: Chemistry, biosynthesis, analysis, and potential health effects. Adv. Food Nutr. Res. 2009, 55, 1–99. [Google Scholar]
- Kim, C.S.; Choi, K.J.; Kim, S.C.; Ko, S.Y.; Sung, H.S.; Lee, Y.G. Controls of the hydrolysis of ginseng saponins by neutralization of organic acids in red ginseng extract preparations. J. Ginseng Res. 1998, 22, 205–210. [Google Scholar]
- Xu, X.F.; Gao, Y.; Xu, S.Y.; Liu, H.; Xue, X.; Zhang, Y.; Zhang, H.; Liu, M.N.; Xiong, H.; Lin, R.C.; et al. Remarkable impact of steam temperature on ginsenosides transformation from fresh ginseng to red ginseng. J. Ginseng Res. 2018, 42, 277–287. [Google Scholar] [CrossRef]
- In, G.; Ahn, N.G.; Bae, B.S.; Lee, M.W.; Park, H.W.; Jang, K.H.; Cho, B.G.; Han, C.K.; Park, C.K.; Kwak, Y.S. In situ analysis of chemical components induced by steaming between fresh ginseng, steamed ginseng, and red ginseng. J. Ginseng Res. 2017, 41, 361–369. [Google Scholar] [CrossRef] [Green Version]
- Thetsrimuang, C.; Khammuang, S.; Chiablaem, K.; Srisomsap, C.; Sarnthima, R. Antioxidant properties and cytotoxicity of crude polysaccharides from Lentinus polychrous Lév. Food Chem. 2011, 128, 634–639. [Google Scholar] [CrossRef]
- Schepetkin, I.A.; Quinn, M.T. Botanical polysaccharides: Macrophage immunomodulation and therapeutic potential. Int. Immunopharmacol. 2006, 6, 317–333. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.H.; Park, H.R.; Shin, M.S.; Cho, S.Y.; Choi, H.J.; Shin, K.S. Antitumor metastasis activity of pectic polysaccharide purified from the peels of Korean Citrus Hallabong. Carbohydr. Polym. 2014, 111, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Shin, M.S.; Hwang, S.H.; Yoon, T.J.; Kim, S.H.; Shin, K.S. Polysaccharides from ginseng leaves inhibit tumor metastasis via macrophage and NK cell activation. Int. J. Biol. Macromol. 2017, 103, 1327–1333. [Google Scholar] [CrossRef] [PubMed]
- Ruijun, W.; Shi, W.; Yijun, X.; Mengwuliji, T.; Lijuan, Z.; Yumin, W. Antitumor effects and immune regulation activities of a purified polysaccharide extracted from Juglan regia. Int. J. Biol. Macromol. 2015, 72, 771–775. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Cho, S.M.; Kim, W.J.; Hong, K.B.; Suh, H.J.; Yu, K.W. Red ginseng polysaccharide alleviates cytotoxicity and promotes anti-inflammatory activity of ginsenosides. Food Sci. Technol. 2021, 41, 1–6. [Google Scholar]
- Uthaisangsook, S.; Day, N.K.; Bahna, S.L.; Good, R.A.; Haraguchi, S. Innate immunity and its role against infections. Ann. Allergy Asthma Immunol. 2002, 88, 253–264. [Google Scholar] [CrossRef]
- Park, D.H.; Han, B.; Shin, M.S.; Hwang, G.S. Enhanced intestinal immune response in mice after oral administration of Korea red ginseng-derived polysaccharide. Polymers 2020, 12, 2186. [Google Scholar] [CrossRef]
- Shin, K.S. Immunostimulating plant polysaccharides: Macrophage immunomodulation and its possible mechanism. Food Sci. Ind. 2012, 45, 12–22. [Google Scholar]
- West, A.P.; Brodsky, I.E.; Rahner, C.; Woo, D.K.; Erdjument-Bromage, H.; Tempst, P.; Walsh, M.C.; Choi, Y.; Shadel, G.S.; Ghosh, S. TLR signalling augments macrophage bactericidal activity through mitochondrial ROS. Nature 2011, 472, 476–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, D.G.; Kim, M.C.; Park, M.K.; Park, K.M.; Quan, F.S.; Song, J.M.; Wee, J.J.; Wang, B.Z.; Cho, Y.K.; Compans, R.W.; et al. Protective effect of ginseng polysaccharides on influenza viral infection. PLoS ONE 2012, 7, e33678. [Google Scholar] [CrossRef] [PubMed]
- MacMicking, J.; Xie, Q.W.; Nathan, C. Nitric oxide and macrophage function. Annu. Rev. Immunol. 1997, 15, 323–350. [Google Scholar] [CrossRef]
- Lee, S.J.; In, G.; Han, S.T.; Lee, M.H.; Lee, J.W.; Shin, K.S. Structural characteristics of a red ginseng acidic polysaccharide rhamnogalacturonan I with immunostimulating activity from red ginseng. J. Ginseng Res. 2020, 44, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Stimpel, M.; Proksch, A.; Wagner, H.; Lohmann-Matthes, M.L. Macrophage activation and induction of macrophage cytotoxicity by purified polysaccharide fractions from the plant Echinacea purpurea. Infect. Immun. 1984, 46, 845–849. [Google Scholar] [CrossRef] [Green Version]
- Byeon, S.E.; Lee, J.; Kim, J.H.; Yang, W.S.; Kwak, Y.S.; Kim, S.Y.; Choung, E.S.; Rhee, M.H.; Cho, J.Y. Molecular mechanism of macrophage activation by red ginseng acidic polysaccharide from Korean red ginseng. Mediat. Inflamm. 2012, 2012, 732860. [Google Scholar] [CrossRef]
- Rao, K.M.K. MAP kinase activation in macrophages. J. Leukoc. Biol. 2001, 69, 3–10. [Google Scholar] [PubMed]
- Kim, H.; Kim, H.W.; Yu, K.W.; Suh, H.J. Polysacchrides fractionated from enzyme digests of Korean red ginseng water extracts enhanced the immunostimulatory activity. Int. J. Biol. Macromol. 2019, 121, 913–920. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, J.; Lee, S.; Lee, J.-H.; Kang, K.-S.; Shin, M.-S. Signaling Pathways Associated with Macrophage-Activating Polysaccharide Isolated from Korea Red Ginseng. Appl. Sci. 2021, 11, 7111. https://0-doi-org.brum.beds.ac.uk/10.3390/app11157111
Gao J, Lee S, Lee J-H, Kang K-S, Shin M-S. Signaling Pathways Associated with Macrophage-Activating Polysaccharide Isolated from Korea Red Ginseng. Applied Sciences. 2021; 11(15):7111. https://0-doi-org.brum.beds.ac.uk/10.3390/app11157111
Chicago/Turabian StyleGao, Jie, Sullim Lee, Ji-Hwan Lee, Ki-Sung Kang, and Myoung-Sook Shin. 2021. "Signaling Pathways Associated with Macrophage-Activating Polysaccharide Isolated from Korea Red Ginseng" Applied Sciences 11, no. 15: 7111. https://0-doi-org.brum.beds.ac.uk/10.3390/app11157111