
Lipoxygenase Enzymes, Oligosaccharides (Raffinose and Stachyose) and 11sA4 and A5 Globulins of Glycinin Present in Soybean Meal Are Not Drivers of Enteritis in Juvenile Atlantic Salmon (Salmo salar)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Formulations and Feed Manufacture
2.2. Experimental System and Feeding Trial
2.3. Analytical Methods
2.4. Production Parameters
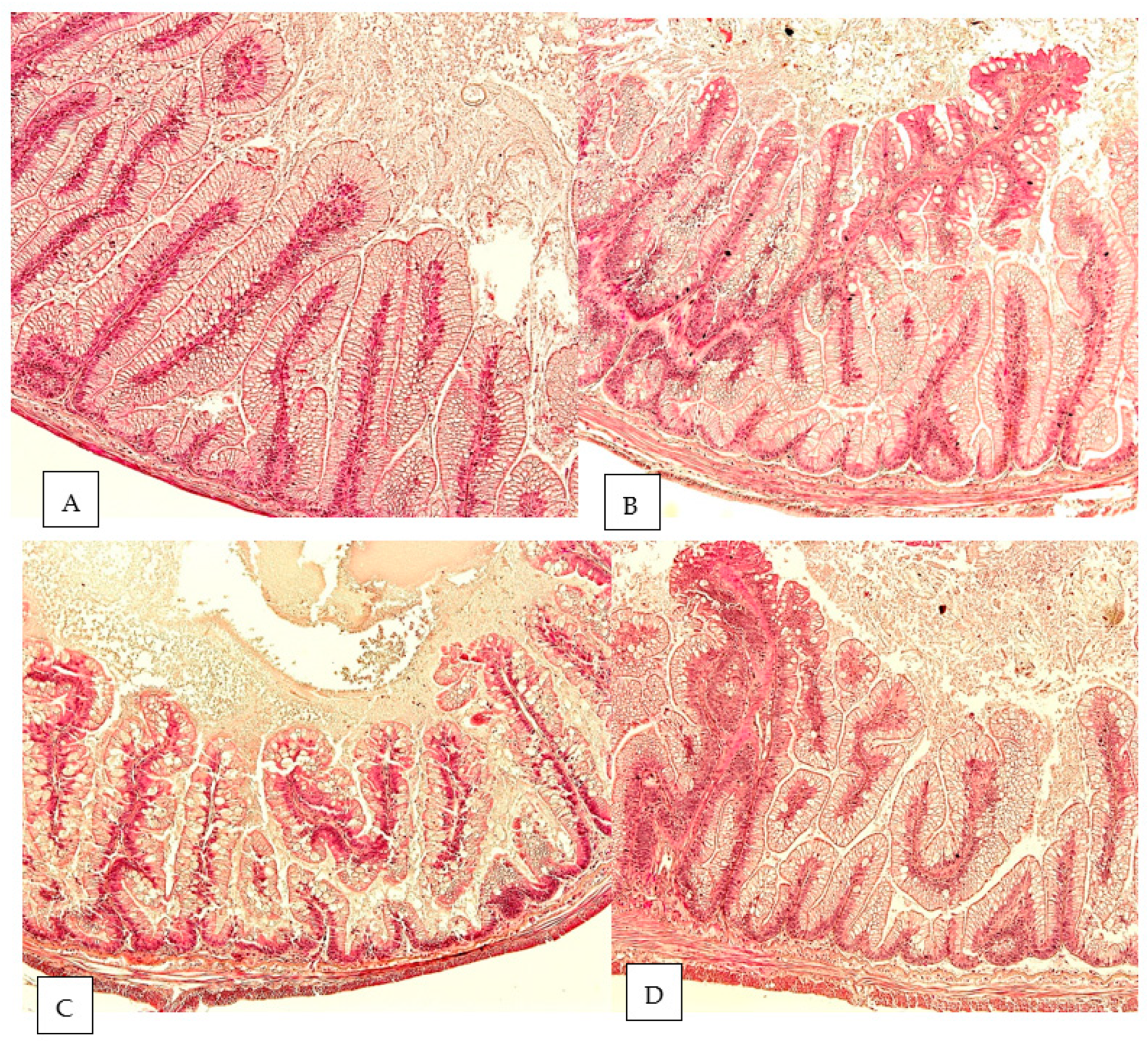
2.5. Histology
2.6. Statistical Analysis
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gatlin, D.M.; Barrows, F.T.; Brown, P.; Dabrowski, K.; Gaylord, T.G.; Hardy, R.W.; Herman, E.; Hu, G.; Krogdahl, A.; Nelson, R.; et al. Expanding the utilization of sustainable plant products in aquafeeds: A review. Aquac. Res. 2007, 38, 551–579. [Google Scholar] [CrossRef]
- NRC (National Research Council). Nutrient Requirements of Fish and Shrimp; Natlional Academic Press: Cambridge, MA, USA, 2011. [Google Scholar] [CrossRef]
- Bakke-McKellep, A.M.; Press, C.M.; Baeverfjord, G.; Krogdahl, A.; Landsverk, T. Changes in immune and enzyme histochemical phenotypes of cells in the intestinal mucosa of Atlantic salmon, Salmo salar L., with soybean meal-induced enteritis. J. Fish Dis. 2000, 23, 115–127. [Google Scholar] [CrossRef]
- Krogdahl, A.; Bakke-McKellep, A.M.; Baeverfjord, G. Effects of graded levels of standard soybean meal on intestinal structure, mucosal enzyme activities, and pancreatic responses in Atlantic salmon (Salmo salar L.). Aquac. Nutr. 2003, 9, 361–371. [Google Scholar] [CrossRef]
- Fuentes-Quesada, J.P.; Viana, M.T.; Rombenso, A.N.; Guerrero-Renteria, Y.; Nomura-Solis, M.; Gomez-Calle, V.; Lazo, J.P.; Mata-Sotres, J.A. Enteritis induction by soybean meal in Totoaba macdonaldi diets: Effects on growth performance, digestive capacity, immune response and distal intestine integrity. Aquaculture 2018, 495, 78–89. [Google Scholar] [CrossRef]
- Fuentes-Quesada, J.P.; Cornejo-Granados, F.; Mata-Sotres, J.A.; Ochoa-Romo, J.P.; Rombenso, A.N.; Gurerrero-Renteria, Y.; Lazo, J.P.; Pohlenz, C.; Ochoa-Leyva, A.; Viana, M.T. Prebiotic agavin in juvenile tototaba, Totoaba macdonaldi diets, to relieve soybean meal-induced enteritis: Growth performance, gut histology and microbiota. Aquac. Nutr. 2020. [Google Scholar] [CrossRef]
- Viana, M.T.; Rombenso, A.N.; Del Rio-Zaragoza, O.B.; Nomura, M.; Diaz-Arguello, R.; Mata-Sotres, J.A. Intestinal impairment of the California yellowtail, Seriola dorsalis, using soybean meal in the diet. Aquaculture 2019, 513, 734443. [Google Scholar] [CrossRef]
- Rossi, W., Jr.; Allen, K.M.; Habte-Tsion, H.-M.; Meesala, K.-M. Supplementation of glycine, prebiotic, and nucleotides in soybean meal-based diets for largemouth bass (Micropterus salmoides): Effects on production performance, whole-body nutrient composition and retention, and intestinal histopathology. Aquaculture 2021, 532, 736031. [Google Scholar] [CrossRef]
- Krogdahl, A.; Kortner, T.M.; Jaramillo-Torres, A.; Gamil, A.A.A.; Chikwati, E.; Li, Y.; Schmidt, M.; Herman, E.; Hymowitz, T.; Teimouri, S.; et al. Removal of three proteinaceous antinutrients from soybean does not mitigate soybean-induced enteritis in Atlantic salmon (Salmo salar, L.). Aquaculture 2020, 514, 734495. [Google Scholar] [CrossRef]
- Uran, P.A.; Schrama, J.W.; Rombout, J.H.W.M.; Obach, A.; Jensen, L.; Koppe, W.; Verreth, J.A.J. Soybean meal-induced enteritis in Atlantic salmon (Salmo salar L.) at different temperatures. Aquac. Nutr. 2008, 14, 324–330. [Google Scholar] [CrossRef]
- Mata-Sotres, J.A.; Viana, M.T.; Tinajero, A.; Del Rio-Zaragoza, O.; Skrzynska, A. Estrés nutricional en peces y los nuevos desafíos de la nutrición. In Revista Bio Ciencias–Coloquio de Nutrigenomica y Biotecnología Acuicola 2020; Universidad Autónoma de Nayarit: Tepic, Mexico, 2021; p. e1132. [Google Scholar] [CrossRef]
- Stone, D.A.J.; Bellgrove, E.J.; Forder, R.E.A.; Howarth, G.S.; Bansemer, M.A. Inducing subacute enteritis in yellowtail kingfish Seriola lalandi: The effect of dietary inclusion of soybean meal and grape seed extract on hindgut morphology and inflammation. N. Am. J. Aquac. 2018, 80, 59–68. [Google Scholar] [CrossRef]
- Legrand, T.P.R.A.; Wynne, J.W.; Weyrich, L.S.; Oxley, A.P.A. Investigation both mucosal immunity and microbiota in response to gut enteritis in yellowtail kingfish. Microorganisms 2020, 8, 1267. [Google Scholar] [CrossRef]
- Uran, P.A.; Goncalvez, A.A.; Taverne-Thiele, J.J.; Schrama, J.W.; Verreth, J.A.J.; Rombout, J.H.W.M. Soybean meal induces intestinal inflammation in common carp (Cyprinus carpio L.). Fish Shellfish Immunol. 2008, 25, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Van Der Marel, M.; Propsting, M.J.; Battermann, F.; Jung-Schroers, V.; Hubner, A.; Rombout, J.H.W.M.; Steinhagen, D. Differences between intestinal segments and soybean meal induced changes in intestinal mucus composition of common carp Cyprinus carpio L. Aquac. Nutr. 2014, 20, 12–24. [Google Scholar] [CrossRef]
- Van Den Ingh, T.S.G.A.M.; Krogdahl, A.; Olli, J.J.; Hendriks, H.G.C.J.M.; Koninkx, J.G.J.F. Effects of soybean-containing diets on the proximal and distal intestine in Atlantic salmon (Salmo salar) L. a morphological study. Aquaculture 1991, 94, 297–305. [Google Scholar] [CrossRef]
- Baeverfjord, G.; Krogdahl, A. Development and regression of soybean meal induced enteritis in Atlantic salmon, Salmo salar L.; distal intestine: A comparison with the intestines of fasted fish. J. Fish Dis. 1996, 19, 375–387. [Google Scholar] [CrossRef]
- Booman, M.; Forster, I.; Vederas, J.C.; Groman, D.B.; Jones, S.R.M. Soybean meal-induced enteritis in Atlantic salmon (Salmo salar) and Chinook salmon (Oncorhynchus tshawystscha) but not in pink salmon (O. gorbuscha). Aquaculture 2018, 483, 238–243. [Google Scholar] [CrossRef]
- Buttle, L.G.; Burrells, A.C.; Good, J.E.; Williams, P.D.; Southgate, P.J.; Burrells, C. The binding of soybean agglutinin (SBA) to the intestinal epithelium of Atlantic salmon, Salmo salar and Rainbow trout, Oncorhynchus mykiss, fed high levels of soybean meal. Vet. Immunol. Immunopathol. 2001, 80, 237–244. [Google Scholar] [CrossRef]
- Krogdahl, A.; Gajardo, K.; Kortner, T.M.; Penn, M.; Gu, M.; Berge, G.M.; Bakke, A.M. Soya saponins induce enteritis in Atlantic salmon (Salmo salar L.). J. Agric. Food Chem. 2015, 63, 3887–3902. [Google Scholar] [CrossRef] [PubMed]
- Barrows, F.T.; Stone, D.A.J.; Hardy, R.W. The effects of extrusion conditions on the nutritional value of soybean meal for rainbow trout (Oncorhynchus mykiss). Aquaculture 2007, 265, 244–252. [Google Scholar] [CrossRef]
- Yang, A.; Smyth, H.; Chalija, M.; James, A. Sensory quality of soymilk and tofu from soybeans lacking lipoxygenases. Food Sci. Nutr. 2016, 4, 207–215. [Google Scholar] [CrossRef]
- James, A.T.; Yang, A. Interactions of protein content and globulin subunit composition of soybean proteins in relation to tofu gel properties. Food Chem. 2016, 194, 284–289. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis; Association of Analytical Chemists: Arlington, VA, USA, 2005. [Google Scholar]
- Folch, J.; Less, M.; Sloane-Stanley, G.H. A simple method for the isolation and purification of total lipid from animal tissues. J. Biol. Chem. 1957, 276, 497–507. [Google Scholar] [CrossRef]
- Feng, L.; Ni, P.-J.; Jiang, W.-D.; Wu, P.; Liu, Y.; Jiang, J.; Kuang, S.-Y.; Tang, L.; Tang, W.-N.; Zhang, Y.-A.; et al. Decreased enteritis resistance ability by dietary low or excess levels of lipids through impairing the intestinal physical and immune barriers function of young grass carp (Ctenopharyngodon Idella). Fish Shellfish Immunol. 2017, 67, 493–512. [Google Scholar] [CrossRef] [PubMed]
- Olli, J.J.; Krogdahl, A.; Vabano, A. Dehulled solvent-extracted soybean meal as a protein source in diets for Atlantic salmon, Salmo salar L. Aquac. Res. 1995, 26, 167–174. [Google Scholar] [CrossRef]
- Knudsen, D.; Uran, P.; Arnous, A.; Koppe, W.; Frokler, H. Saponing-containing subfractions of soybean molasses induce enteritis in the distal intestine of Atlantic salmon. J. Agric. Food Chem. 2007, 55, 2261–2267. [Google Scholar] [CrossRef] [PubMed]
- Uran, P.A.; Schrama, J.W.; Jaafari, S.; Baardsen, G.; Rombout, J.H.W.M.; Koppe, W.; Verreth, J.A.J. Variation in commercial sources of soybean meal influences the severity of enteritis in Atlantic salmon (Salmo salar L.). Aquac. Nutr. 2009, 15, 492–499. [Google Scholar] [CrossRef]
- Chikwati, E.M.; Sahlmann, C.; Holm, H.; Penn, M.H.; Krogdahl, A.; Bakke, A.M. Alterations in digestive enzyme activities during the development of diet-induced enteritis in Atlantic salmon, Salmo salar L. Aquaculture 2013, 402–403, 28–37. [Google Scholar] [CrossRef]
- Sahlmann, C.; Sutherland, B.J.G.; Kortner, T.M.; Koop, B.F.; Krogdahl, A.; Bakke, A.M. Early response of gene expression in the distal intestine of Atlantic salmon (Salmo salar L.) during the development of soybean meal induced enteritis. Fish Shellfish Immunol. 2013, 34, 599–609. [Google Scholar] [CrossRef]
- Silva, P.F.; McGurk, C.; Knudsen, D.L.; Adams, A.; Thompson, K.D.; Bron, J.E. Histological evaluation of soya bean-induced enteritis in Atlantic salmon (Salmo salar L.): Quantitative image analysis vs. semi-quantitative visual scoring. Aquaculture 2015, 445, 42–56. [Google Scholar] [CrossRef]
- Jacobsen, H.J.; Samuelsen, T.A.; Girons, A.; Kousoulaki, K. Different enzyme incorporation strategies in Atlantic salmon diet containing soybean meal: Effects on feed quality, fish performance, nutrient digestibility and distal intestinal morphology. Aquaculture 2018, 491, 302–309. [Google Scholar] [CrossRef]
- Refstie, S.; Korsoen, O.J.; Storebakken, T.; Baeverfjord, G.; Lein, I.; Roem, A.J. Differing nutritional responses to dietary soybean meal in rainbow trout (Oncorhynchus mykiss) and Atlantic salmon (Salmo salar). Aquaculture 2000, 190, 49–63. [Google Scholar] [CrossRef]


{kind=link}
| Ingredients g kg−1 | FM Control | STD SBM | TLP SBM | 11sA4 Null SBM |
|---|---|---|---|---|
| Fishmeal a | 450.0 | 290.0 | 290.0 | 290.0 |
| Wheat flour | 223.0 | 110.0 | 93.0 | 106.0 |
| Wheat gluten | 120.0 | 89.0 | 106.0 | 93.0 |
| Blood meal a | 60.0 | 60.0 | 60.0 | 60.0 |
| Fish oil a | 70.0 | 70.0 | 70.0 | 70.0 |
| Poultry oil a | 70.0 | 70.0 | 70.0 | 70.0 |
| Stay-C 35% b | 1.0 | 1.0 | 1.0 | 1.0 |
| Vitamin mineral premix c | 6.0 | 6.0 | 6.0 | 6.0 |
| Standard soybean meal (STD SBM) d | 0.0 | 300.0 | 0.0 | 0.0 |
| Soybean meal triple lipoxygenase plus (TLP SBM) d | 0.0 | 0.0 | 300.0 | 0.0 |
| Soybean meal 11sA4 null (11sA4 null SBM) d | 0.0 | 0.0 | 0.0 | 300.0 |
| Methionine e | 0.0 | 3.0 | 3.0 | 3.0 |
| Taurine f | 0.0 | 1.0 | 1.0 | 1.0 |
| Proximate composition (g kg−1) | ||||
| Dry matter | 959 | 960 | 954 | 954 |
| Protein | 506 | 492 | 494 | 505 |
| Lipid | 166 | 171 | 179 | 170 |
| Ash | 68 | 63 | 62 | 62 |
| Gross energy (kJ g−1) | 23.7 | 24.2 | 24.3 | 24.3 |
| Amino acid (g kg−1) | ||||
| ASP | 39 | 43 | 45 | 43 |
| SER | 20 | 22 | 23 | 21 |
| GLU | 88 | 87 | 93 | 89 |
| GLY | 26 | 24 | 26 | 23 |
| HIS | 16 | 15 | 15 | 15 |
| ARG | 22 | 25 | 25 | 26 |
| THR | 19 | 19 | 20 | 19 |
| ALA | 25 | 24 | 24 | 24 |
| PRO | 29 | 28 | 29 | 28 |
| CYS | 5 | 5 | 5 | 5 |
| TYR | 14 | 19 | 20 | 19 |
| VAL | 23 | 23 | 24 | 23 |
| MET | 11 | 11 | 11 | 10 |
| LYS | 26 | 28 | 28 | 28 |
| ILE | 17 | 18 | 19 | 18 |
| LEU | 38 | 38 | 39 | 38 |
| PHE | 22 | 23 | 25 | 23 |
| TAU | 3 | 3 | 3 | 3 |
| Parameter | FM Control | STD SBM | TLP SBM | 11sA4 Null SBM |
|---|---|---|---|---|
| RPM | 220 | 220 | 220 | 220 |
| Feed (g min−1) | 72 | 62 | 59 | 53 |
| H2O (g min−1) | 16 | 22 | 20 | 20 |
| Torque (%) | 8 | 6 | 6 | 6 |
| SME (kJ kg−1) | 99 | 69 | 73 | 80 |
| Zone 1 (Cone) | 120 | 120 | 120 | 120 |
| Zone 2 | 95 | 95 | 95 | 95 |
| Zone 3 | 85 | 85 | 85 | 85 |
| Zone 4 (Inlet) | 70 | 70 | 70 | 70 |
| Die Diameter (mm) | 1.5 | 1.5 | 1.5 | 1.5 |
| Fishmeal | STD SBM | TLP SBM | 11sA4 null SBM | |
|---|---|---|---|---|
| Proximate composition (g kg−1) | ||||
| Dry matter | 911 | 949 | 918 | 926 |
| Protein | 719 | 495 | 521 | 462 |
| Lipid | 117 | 119 | 116 | 118 |
| Ash | 126 | 54 | 52 | 58 |
| Amino acid (g kg−1) | ||||
| ASP | 67.7 | 46.0 | 48.5 | 48.7 |
| SER | 29.8 | 19.6 | 20.8 | 21.6 |
| GLU | 97.4 | 69.5 | 73.4 | 69.1 |
| GLY | 45.2 | 16.0 | 16.3 | 16.7 |
| HIS | 16.2 | 9.1 | 10.2 | 9.3 |
| ARG | 39.7 | 24.6 | 26.0 | 25.0 |
| THR | 30.8 | 14.3 | 15.2 | 16.3 |
| ALA | 40.7 | 16.3 | 17.5 | 17.6 |
| PRO | 31.6 | 19.3 | 20.5 | 19.7 |
| CYS | 6.5 | 5.5 | 5.2 | 5.3 |
| TYR | 24.4 | 11.7 | 13.9 | 13.2 |
| VAL | 34.4 | 16.7 | 17.5 | 16.7 |
| MET | 22.8 | 2.8 | 3.0 | 3.1 |
| LYS | 58.9 | 21.1 | 23.7 | 23.4 |
| ILE | 29.0 | 15.8 | 16.7 | 17.5 |
| LEU | 52.6 | 27.6 | 30.4 | 29.8 |
| PHE | 30.1 | 18.2 | 18.8 | 20.4 |
| TAU | 7.8 | ND | ND | ND |
| Scoring | FM Control | STD SBM | TLP SBM | 11sA4 Null SBM | p-Value |
|---|---|---|---|---|---|
| Goblet cells | 1.9 ± 0.3 b | 3.2 ± 0.4 a | 3.4 ± 0.5 a | 3.7 ± 0.5 a | <0.001 |
| Lamina propria | 1.8 ± 0.4 b | 3.2 ± 0.4 a | 3.5 ± 0.5 a | 3.6 ± 0.5 a | <0.001 |
| Mucosal fold | 1.9 ± 0.5 b | 2.7 ± 0.7 ab | 2.9 ± 0.6 a | 3.3 ± 0.7 a | 0.001 |
| Blood Chemistry Parameters | FM Control | STD SBM | TLP SBM | 11sA4 Null SBM | p-Value |
|---|---|---|---|---|---|
| Albumin (g L−1) | 20.3 ± 0.6 ab | 20.0 ± 1.0 b | 22.3 ± 1.1 a | 20.7 ± 0.6 ab | 0.044 |
| Alkaline phosphatase (U L−1) | 21.3 ± 6.1 | 18.7 ± 4.7 | 24.3 ± 2.1 | 15.7 ± 5.7 | 0.242 |
| Anion gap (mmol L−1) | 59.3 ± 3.5 | 56.0 1.0 | 53.0 ± 2.6 | 58.0 ± 8.7 | 0.463 |
| AST (U L−1) | 301.3 ± 53.6 | 301.7 ± 28.7 | 359.5 ± 2.1 | 315.7 ± 31.5 | 0.361 |
| Bicarbonate (mmol L−1) | 6.3 ± 0.6 | 5.3 ± 1.1 | 5.7 ± 0.6 | 6.7 ± 2.1 | 0.577 |
| Chloride (mmol L−1) | 123.3 ± 8.7 | 120.3 ± 1.5 | 125.3 ± 5.1 | 120.0 ± 1.0 | 0.561 |
| Cholesterol (mmol L−1) | 13.2 ± 2.9 | 9.8 ± 1.1 | 12.4 ± 0.9 | 11.9 ± 1.4 | 0.236 |
| Creatine kinase (U L−1) | 7286 ±1425 | 7992 ± 1198 | 7446 ± 1719 | 8089 ± 1674 | 0.890 |
| Globulin (g L−1) | 18.7 ± 2.1 | 18.7 ± 1.1 | 21.0 ± 1.0 | 20.3 ± 0.6 | 0.142 |
| Glucose (mmol L−1) | 5.2 ± 0.8 | 4.2 ± 0.3 | 4.2 ± 0.2 | 4.0 ± 0.4 | 0.055 |
| Phosphate (mmol L−1) | 3.1 ± 0.3 | 2.7 ± 0.3 | 2.6 ± 0.3 | 2.5 ± 0.3 | 0.103 |
| Potassium (µm mmol−1) | 4.2 ± 0.6 | 4.1 ±0.6 | 4.1 ± 0.5 | 4.0 ± 0.4 | 0.963 |
| Protein (g L−1) | 39.0 ± 2.6 ab | 38.7 ± 1.5 b | 43.3 ± 1.5 a | 41.0 ± 1.0 ab | 0.042 |
| Ratio | 1.1 ± 0.1 | 1.1 ± 0.1 | 1.0 ± 0.1 | 1.0 ± 0.0 | 0.344 |
| Sodium (mmol L−1) | 184.7 ± 11.6 | 177.7 ± 0.6 | 179.7 ± 3.0 | 180.3 ± 5.1 | 0.683 |
| Triglyceride (mmol L−1) | 2.9 ± 0.4 | 3.5 ± 0.5 | 3.1 ± 0.8 | 3.5 ± 0.3 | 0.377 |
| Production Performance | FM Control | STD SBM | TLP SBM | 11sA4 Null SBM |
|---|---|---|---|---|
| 0 day | ||||
| Initial weight (g) | 37.3 ± 0.0 | 37.4 ± 0.0 | 37.3 ± 0.1 | 37.3 ± 0.0 |
| Initial CV (%) | 12.1 ± 0.5 | 15.7 ± 3.4 | 11.1 ± 0.8 | 11.5 ± 0.5 |
| 28 days | ||||
| Survival (%) | 99 ± 0.0 | 97 ± 0.0 | 99 ± 0.0 | 100 |
| CV (%) | 12.3 ± 1.3 | 13.0 ± 0.5 | 12.1 ± 0.7 | 13.1 ± 0.9 |
| Mid weight (g) | 49.6 ± 0.6 | 52.1 ± 0.7 | 49.8 ± 0.7 | 51.4 ± 0.6 |
| FCR | 1.0 ± 0.0 | 1.0 ± 0.0 | 1.1 ± 0.1 | 1.0 ± 0.0 |
| K | 1.3 ± 0.07 | 1.4 ± 0.09 | 1.5 ± 0.09 | 1.4 ± 0.01 |
| HSI | 1.8 ± 0.16 | 1.7 ± 0.17 | 1.6 ± 0.19 | 1.9 ± 0.40 |
| 56 days | ||||
| Survival (%) | 98.5 ± 1.5 | 97.0 ± 1.5 | 98.5 ± 1.5 | 100 |
| CV (%) | 15.2 ± 1.5 | 15.5 ± 0.8 | 14.6 ± 1.0 | 15.2 ± 0.7 |
| Final weight (g) | 59.4 ± 1.5 | 66.1 ± 1.2 | 65.1 ± 2.1 | 64.4 ± 0.7 |
| FCR | 1.0 ± 0.0 | 0.9 ± 0.0 | 1.0 ± 0.0 | 0.9 ± 0.0 |
| K | 1.5 ± 0.17 | 1.4 ± 0.02 | 1.4 ± 0.04 | 1.3 ± 0.03 |
| HSI | 1.6 ± 0.19 | 1.4 ± 0.11 | 1.4 ± 0.07 | 1.6 ± 0.25 |
| Experimental Design | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| IBW (g) | SGR (% BW) | FCR | Tank Volume (L) | Stocking Density (Fish/Tank) | Duration (Days) | Salinity | Temp. (°C) | Feeding | System | Refeed to Control | Ref. |
| 927 | 2000 | 25 | 21 | FW | 12–13 | Semi-RAS | [3] | ||||
| 280 | 0.94–1.05 | 0.81–0.94 | 450 | 54 | 60 | SW | 8.4 | [4] | |||
| 41 | 0.7 | 250 | 55 | 56 | FW | 14 | RAS | [9] | |||
| 300 | 400 | 50 | 20 | SW | 8 and 12 | 120% | RAS | [10] | |||
| 550 | 27,000 | 120 | 3.2 | 5 | 120% | Net pens | [16] | ||||
| 550 | 27,000 | 80 | 42 | SW | 10.8 and 8.2 | 120% | Net pens | Yes | [17] | ||
| 535|140|166 * | 1900|650|850 | 50 | 21 | SW | 9 | 1% BW | Flow-through | [18] | |||
| 54 | 1000 | 20 | 7 | FW | 15 | [19] | |||||
| 442 | 1000 | 22 | SW | 12–9 | RAS | [20] | |||||
| 900 | 125 000 | 300 | 300 | SW | 8.2 | Net pens | [27] | ||||
| 213|202 | 400|100 | 30|20 | 62|44 | SW | 8.3|9 | 120% | RAS | [28] | |||
| 396 | 400 | 25 | 28 | SW | 12 | 110% | RAS | [29] | |||
| 500–600 | 1000 | 25 | 21 | SW | 9 | RAS | [30] | ||||
| 500–600 | 1000 | 25–30 | 21 | SW | 9 | RAS | [31] | ||||
| 80 | 400 | 70 | 53 | SW | 8.6 | 120% | Flow-through | [32] | |||
| 60 | 1.36–1.47 | 0.71–0.74 | 500 | 75 | 93 | SW | 10.9 | 120% | Flow-through | [33] | |
| 207 | 0.7–0.9 | 0.92–1.17 | 600 | 50 | 84 | FW | 7 | 115% | Flow-through | [34] | |
| SBM Information | Dietary Treatments | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Location | Type | CP (%) | Pellet Type | CP (%) | TL (%) | GE (MJ /kg) | SBM Type/Inclusion (%) | SBM Type/Inclusion (%) | SBO Inclusion (%) | Ref. |
| Norway | With hulls, toasted and extracted | Extruded | 35 | 28 | SBM 30 | [3] | ||||
| Norway | Solvent-extracted, toasted | Extruded | 40–44 | 22–24 | 22–23 | SBM—8, 12, 15, 19, 27 | [4] | |||
| USA | Triple null and standard | Extruded | 40 | 26 | 24 | SBM 25 | Triple null SBM 27 | [9] | ||
| Netherlands | Extracted | Extruded | 45 | 30 | SBM 20 | [10] | ||||
| 43 | 20 | 15 | FFSBM 30 | SPC 28 | 0–10 | [16] | ||||
| SBM 33 | 0–8.5 | [17] | ||||||||
| Pelleted | 37 | 23 | SBM 20 | [18] | ||||||
| Soybean agglutinin | Soybean agglutinin 3.5 | [19] | ||||||||
| Soya saponins | 42–44 | 29–30 | 24 | Soy saponins—0, 2, 4, 6, 10 | [20] | |||||
| Dehulled solvent extracted SBM | Extruded | 40 | 22 | SBM 17 and 34 | [27] | |||||
| Norway | Defatted | Extruded | DSBM 20|molasses | [28] | ||||||
| NA, EU, SA | 44–49 | Extruded | 42 | 25 | 23 | 20 | [29] | |||
| Extracted | 43 | 28 | 24 | SBM 20 | [30] | |||||
| Extracted | 43 | 28 | 24 | SBM 20 | [31] | |||||
| Norway | Defatted | Extruded | 47 | 26 | 23 | SBM 25 + soy saponins 0.17 | Lupins + soy saponin concentrate 0.17 + soy saponins 0.11 | [32] | ||
| Switzerland | Extruded | 46 | 25 | 24 | SBM 20 | Pre-processed SBM 20 | [33] | |||
| Dehulled defatted SBM | Pelleted | 40 | 31 | 24 | SBM 30 | [34] | ||||
| Growth Impairment | Feed Intake | Level of DI Inflammation | Enzymes | Time Sampling (Days) | Tissues Analyzed | Parameters | Ref. |
|---|---|---|---|---|---|---|---|
| SBM yes SPC no | SBM +++ | PI and DI | MSA/LSC, ESA/LSC, LPSA/LSC, ESA/LPSA, GC/E, LM | [16] | |||
| Yes | +++ | 2, 7, 14 and 21 | DI | MF | [17] | ||
| ++ | 5′ N, Mg-ATPase, ALP, ACP, NSE, LAP, AAP | 21 | MI and DI | [3] | |||
| Low SBM no High SBM yes | No changes | +, ++ and +++ | ALP, LAP, maltase, isomaltase, lactase and sucrase | MI and DI | MF, SNV, LP, leycocyte | [4] | |
| +, ++ and +++ | DI | MF, SNV, LP, CT | [28] | ||||
| ++ and +++ | DI | MF, GC, LP SNV, EG, SM | [29] | ||||
| ++ and +++ | DI | MF, GC, LP SNV, EG, SM | [10] | ||||
| +, ++, and +++ | Pancreatic (trypsin, chymotrypsin, elastase, and lipase), chyme (LAP), brush border membrane (LAP and maltase) | 0, 1, 2, 3, 5, 7, 10, 14, 17, and 21 | PI, MI and DI | [30] | |||
| +, ++, and +++ | 1, 2, 3, 5 and 7 | DI | [31] | ||||
| Low saponins no Mid-high saponins yes | Low saponins no Mid-high saponins yes | +, ++, and +++ | Trypsin activity, bile acids, brush border membrane enzyme activity (LAP) | PI and DI | MF, LP, enterocyte vacuolization, GC, nucleos position within the enterocytes | [20] | |
| +++ | DI | MF, SNV, LP, CT | [32] | ||||
| +, ++, and +++ | 7, 14 and 21 | DI | Inflammation score, SM and microbiome | [18] | |||
| No | No changes | DI | MF, GC, LP, SNV, EG, SM | [33] | |||
| No | No changes | + and ++ | Brush border LAP, trypsin activity | DI | MF, SNV, SM, LP, microbiota, gene expression | [9] | |
| + and ++ | PI, MI and DI | MF, SNV, LP, CT, | [19] | ||||
| SBM 17 no SBM 34 yes | Body composition and blood biochemistry | [27] | |||||
| Yes | No changes | ++ and +++ | DI | MF, SNV, LP, CT, | [34] | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rombenso, A.N.; Blyth, D.; James, A.T.; Nikolaou, T.; Simon, C.J. Lipoxygenase Enzymes, Oligosaccharides (Raffinose and Stachyose) and 11sA4 and A5 Globulins of Glycinin Present in Soybean Meal Are Not Drivers of Enteritis in Juvenile Atlantic Salmon (Salmo salar). Appl. Sci. 2021, 11, 9327. https://0-doi-org.brum.beds.ac.uk/10.3390/app11199327
Rombenso AN, Blyth D, James AT, Nikolaou T, Simon CJ. Lipoxygenase Enzymes, Oligosaccharides (Raffinose and Stachyose) and 11sA4 and A5 Globulins of Glycinin Present in Soybean Meal Are Not Drivers of Enteritis in Juvenile Atlantic Salmon (Salmo salar). Applied Sciences. 2021; 11(19):9327. https://0-doi-org.brum.beds.ac.uk/10.3390/app11199327
Chicago/Turabian StyleRombenso, Artur N., David Blyth, Andrew T. James, Teisha Nikolaou, and Cedric J. Simon. 2021. "Lipoxygenase Enzymes, Oligosaccharides (Raffinose and Stachyose) and 11sA4 and A5 Globulins of Glycinin Present in Soybean Meal Are Not Drivers of Enteritis in Juvenile Atlantic Salmon (Salmo salar)" Applied Sciences 11, no. 19: 9327. https://0-doi-org.brum.beds.ac.uk/10.3390/app11199327