
Microbial Life on the Surface of Microplastics in Natural Waters
by
 , , and
, , and
Olena Stabnikova
1,2,*, 
Viktor Stabnikov
2,
Andriy Marinin
1,
Maris Klavins
3,*,
Linards Klavins
3 and
Ashok Vaseashta
4,5,6 1
Advanced Research Laboratory, National University of Food Technologies, 01601 Kyiv, Ukraine
2
Department of Biotechnology and Microbiology, National University of Food Technologies, 01601Kyiv, Ukraine
3
Department of Environmental Science, University of Latvia, LV 1586 Riga, Latvia
4
International Clean Water Institute, Manassas, VA 26250, USA
5
Faculty of Chemistry, Riga Technical University, LV 2156 Riga, Latvia
6
Ghitu Institute of Electronic Engineering and Nanotechnologies, 373-22 Chisinau, Moldova
*
Authors to whom correspondence should be addressed.
Appl. Sci. 2021, 11(24), 11692; https://0-doi-org.brum.beds.ac.uk/10.3390/app112411692
Submission received: 10 November 2021
/
Revised: 6 December 2021
/
Accepted: 7 December 2021
/
Published: 9 December 2021
(This article belongs to the Special Issue Environmental Chemical: Pollution, Analysis and Restoration)
Abstract
:Major water-polluting microplastics (for example, polyethylene, polypropylene and others) have lower density than water. Therefore, they are concentrated in the neustonic layer near the water-air interface altogether with dissolved or colloidal natural organic matter, hydrophobic cells and spores of bacteria. This can cause environmental and public health problems because the floating micro- and nanoparticles of plastics could be coated with biofilm of hydrophobic and often putative pathogenic bacteria. Biofilm-coated microplastics are more attractive for consumption by aquatic animals than pure microplastics, and that increases the negative impacts of microplastics. So, impacts of even small quantities of microplastics in aquatic environments must be accounted for considering their accumulation in the micro-layer of water-air interphase and its interaction with bacterioneuston. Microorganisms attached to the surface of microplastic particles could interact with them, use them as substrates for growth, to change properties and biodegrade. The study of microbial life on the surface of microplastic particles is one of the key topics to understanding their role in the environment.
1. Introduction
Plastic pollution has become one of the most widespread recalcitrant environmental contaminants. In 2019 annual plastic production was 368 million tons, and it is expected that its production will increase up to 33 billion tons by 2050 [1,2]. It is estimated that 76% of total plastics produced are landfilled or spread in the natural environment [3]. The total quantity of waste plastics in the marine environment per year is approximately 8 million tons [4] and according to Bowley with co-authors [5] the number of plastic pieces in the ocean was between 15 and 51 trillion. Small sized particles of plastic wastes (microplastics, nanoplastics) are considered as a group of environmental pollutants with significant adverse impacts on the environment. The term “microplastics” first appeared in a study of small plastic particles in 2004 [6]. The definition by Frias and Nash [7] covers major aspects that could potentially describe what a microplastics are: “Microplastics are any synthetic solid particle or polymeric matrix, with regular or irregular shape and with size ranging from 1 μm to 5 mm, of either primary or secondary manufacturing origin, which are insoluble in water”. Another classification approach of small-sized particles in the environment is based on the categorization by the size of particles as macroplastics (>2 cm), mesoplastics (5 mm—2 cm), microplastics (<5 mm), and nanoplastics (<1 μm) [8]. Small-sized plastic particles can be further classified as primary microplastics (plastics directly released into the environment in the form of small particulates) and secondary microplastics (plastics originating from the degradation of larger plastic items into smaller plastic fragments during mechanical, chemical and biological degradation in the environment) [9]. The source of environmental pollution with small plastic particles is non-sustainable use of plastics—discarding of plastic wastes in the environment and relatively lean implementation of recycling and use of biodegradable plastics [10] as well as high stability of several types of plastics, especially hydrocarbon and halogen atoms containing plastics [11]. It was reported that the quantity of microplastic particles in marine aquatic systems can reach 140 particles/m3 and 8766 particles/m3 in water and sediment, respectively [12]. A lot of microplastics are produced due to physical disintegration and chemical or biological destruction of plastic materials, arising mainly from plastic tableware, single-use beverage bottles and cosmetics which disintegrate into fibers and microspheres that enter the environment through discarding on land or water. The main polymer constituents of microplastics found in waters have been identified as polyethylene, polypropylene, polystyrene, and polyethylene terephthalate, accounting for 70% of the total, but polyvinylchloride, polyacrylonitrile, rubber, different copolymers are also common [13], however significant differences exist between studied environments (inland and marine waters, soils, sediments and others) as well as different world regions [14]. MPs are constantly present in fresh and marine water systems. A lot of microplastics debris were found even in oceanic surface waters of the Antarctic Peninsula [15]. It is considered that microplastics have become a main source of anthropogenic pollution of the oceans [5] and MPs concentration in highly contaminated rivers could be up to 100 mg/L. It is evident that the quantity of microplastics will increase over the next decade, so the fate and biological impact on the environment of this contaminant are in focus of scientific research. Microplastics have been found also in freshwater [16], drinking water [17], soil [18] as well as in food articles [19]. It has been demonstrated that there are adverse impacts of microplastics on aquatic and soil living organisms due to impaired reproduction, malnutrition, internal abrasions, and blockages [20,21,22], and also adverse human health impacts have been identified [23,24,25]. Considering their high surface area and hydrophobicity of their surfaces, microplastics can act as sorbents for other environmental pollutants, persistent organic pollutants [26], hydrocarbons [27], pharmaceuticals [28] and other pollutants [29]. Desorption of pollutants, if the particles enter the living body, is one of the microplastic toxicity mechanisms [22]. Sorption of pollutants onto microplastic particles influences polymer, additives to it, particle size and surface area, as well as ageing and supporting development of functional groups [30]. Another major group of factors affecting microplastics—pollutant interaction are the properties of corresponding environments and, in this respect, of key importance are the presence of natural organic matter (NOM) and their refractory part—humic substances. As the concentrations of NOM in aquatic environments are by order higher than those of microplastics, it can be expected that they play a major role at the interaction with pollutants (Figure 1) as well as support the development of microbial coverage on the surface of particles of microplastics. Recently, the attention of researchers has been directed towards the study of nanoplastics which are produced during degradation of microplastic [31].
Microbial life on the surface of microplastic particles is one of the key topics to understanding their role in the environment. Microplastics serve as habitats for microorganisms including pathogenic ones, places for biofilms formation, and means for microbes’ transportation in new areas. Microorganisms developed on microplastics can cause an impact on the environment and human health. Microorganisms attached to the surface of microplastic particles could interact with them, be used as substrates for growth, change properties and biodegrade. However, as it was indicated in work [32], while the problems of plastics and microplastic pollution are widely discussed, the impact of plastic on microbial life remains poorly understood.
The aim of the article is to review the impacts of microorganisms on the surface of microplastic particles to understand the role of these interactions in the environment and the impact on the fate of microplastics.
2. Bacterial Neuston as a Source of Microorganisms for Interaction with Microplastics
The term neuston was proposed by Swedish hydrobiologist E. Naumann [33] for determination of the inhabitants of the surface film of a body of small reservoirs with fresh water. Life of the near-surface layer of the sea remained unexplored for a long time. It is believed that a number of factors, namely, solar radiation, shortwave ultraviolet light penetrating the depths of water just a few centimeters, heaving of the sea, unsteady-state temperature, especially seasonal temperature changes, create impossible living conditions for marine inhabitants. However, Y. Zaitsev in 1959 discovered this when studying the distribution of mullet roe in marine neustons in the Black Sea [34,35,36]: a significant concentration of bacteria in the surface film of different natural bodies of water made it possible to assert the existence of bacterioneuston [37,38]. So, bacterioneuston is the community of Bacteria present within the neuston in the thin surface microlayer between the atmosphere and deeper layers of water in natural water bodies. This surface microlayer is thick and is considered to be the top 1 mm or less of natural water bodies [35,39], but usually the thickness of biofilm is from 10 to 100 µm [40]. The concentration of hydrophobic and surface-active substance and bacterial cells in the bacterial neuston may be some thousand times higher than in the bulk water. This may be due to the increased concentration of organic substances because of their poor water solubility, surface activity and hydrophobic properties in the surface film [41]. It was shown that the sea surface microlayer contains carbohydrates, proteins, lipids, and humic substances in relatively high concentrations [42,43]. The minimum concentration of organic components at which reproduction of saprophytes occurs is in the range from 0.001 to 0.01%. In lake water, their content varies from 1 to 50 mg/L; in the sea from 1 to 5 and can reach 40 mg/L, but in the surface film increases significantly. The study of the distribution of 14C-glucose, 14C-carbonate and 32P-phosphate introduced into water showed that after 24 h they concentrated in the surface layer. The thickness of the biofilm formed depends on the content of organic matter in the water. In addition, the conditions in the biofilm promote the existence of aerobic or microaerobic organisms. For example, the concentration of bacteria in the surface film in the Volga estuary was up to 300 million per ml, which was 100–1000 times higher than in the water column. Most often under natural conditions, cells are in a state of starvation, which is accompanied by an increase of the concentration of bacteria in the film. A correlative relationship between the degree of oligotrophy of lakes and the number of bacteria in the surface layer was shown [44]. The main physico-chemical parameter of the cell surface which determines the interaction of the cell with air-water interface is the hydrophobicity of the cell surface [45,46]. Bacteria concentration can be increased by the addition of a source of nutrients with hydrophobic properties in water.
There are a lot of studies on bacterioneuston composition. Surprisingly, it was found that bacterioneuston in the sea water was characterized with a significantly lower bacterial diversity than the pelagic seawater, and just two groups of organisms were determined, Vibrio spp., 68%, and Pseudoalteromonas spp., 21% of clones [47]. Another study showed the similarity between bacterial compositions of sea bacterioneuston and bacterioplankton communities with a predominance of representatives of two families Flavobacteriaceae and Cryomorphaceae [48]. No significant difference between the bacterial compositions of bacterioneuston and underlying waters cell was found in Mediterranean coastal stations with Proteobacteria as dominated phylum [49] and in communities of neuston in seven sites located across the northern Yellow Sea and Bohai Sea in China [50]. However, there are also a lot of studies that indicate the difference in bacterial compositions of neuston and the subsurface water [51]. Bacterial strains isolated from the surface biofilm were represented in the genera Bacterium, Chromobacterium, Pseudomonas, Flavobacterium, Caulobacter, Corynebacterium and the Enterobacteriaceae family [41,51,52]. Analysis of bacterial neuston compositions in six lakes showed that despite differences in environmental conditions, the fraction of bacteria was very similar and the class of Gram-negative bacteria Betaproteobacteria, the phylum Proteobacteria, was predominant, followed with the phylum of mostly Gram-positive bacteria Actinobacteria [35]. Significantly higher numbers of Gram-negative heterotrophic bacteria in the neuston than in the underlying water layers were found in freshwater lakes [53]. Bacterial composition of bacterial neuston in high mountain lake in the Pyrenees consisted of Betaproteobacteria and Actinobacteria (>75% of the community composition), Gammaproteobacteria (between 8% and 12%), Alphaproteobacteria (between 1% and 5%), and Firmicutes (1%) [54]. So, generally it seems that the bacterial community of bacterioneuston is similar in most cases to bacteria occurred in the water column, however, due to different factors such as physico-chemical, specific environmental and meteorological conditions the development of bacterial population of neuston could result in change of its structure.
There are many cases when the biological hazards and water quality of natural aquatic systems is determined by the biodiversity of the bacterioneuston. At the water—air interface along with bacterioneuston there is pathobacterioneuston and, moreover, the concentration of pathogenic bacteria in the surface film may exceed 10–1000 times their content in the underlying layers of water. A significant number of pathogenic strains were isolated from the surface film of seawater including Aeromonas hydrophila, Escherichia coli, Pseudomonas aeruginosa, Salmonella spp., Staphylococcus aureus, Streptococcus faecalis, Klebsiella pneumonia, Alcaligenes spp., Micrococcus spp., and viruses [36,55,56,57].
Apparently, bacterial neuston plays a special role in the interaction of microplastics and microorganisms. Among microplastics floating in the ocean, the largest amounts belong to polyethylene, 54.5%, and polypropylene, 16.5% from the total quantity of MPs debris. They have lower density than water, have hydrophobic surfaces, and are present on the surface of water bodies [58]. Microbial cells of bacterioneuston, including pathogenic ones which are concentrated at the air-water interface, could attach to microplastic particles followed by biofilm formation.
3. Microplastics as Substrate for the Microbial Colonization in Marine and Freshwater Environments
The main kinds of plastic, approximately 75% of total plastic production, are polyethylene (PE), polypropylene (PP), polystyrene (PS), polyethylene terephthalate (PETE), polyvinyl chloride (PVC), and polyurethane (PU) [1]. Polyethylene, polystyrene and polypropylene particles were identified in the majority of plastic debris found on the surface of North Sea (UK) [59]. Different kinds of microplastic have their own density, g/cm3: polypropylene and polyethylene (0.91); polystyrene (0.96–1.05); polyurethane (1.13); polyamide (1.13–1.35); polyethylene terephthalate (1.38), and polyvinyl chloride (1.47). The difference in plastic density specified its distribution in the water column: plastic with density lower than density of water, particularly, polypropylene and polyethylene floated on the water surface, while polystyrene, polyethylene terephthalate, and polyvinyl chloride with a higher density than water were deposited through the water column [60]. So, presence of microplastic can be expected in every layer of marine systems. However, the character of plastic and microplastic distribution in the layers of water can be changed in time due to its colonization by microorganisms such as bacteria, fungi and algae forming biofilm on plastic surfaces that cause an increase of microplastic density. For example, it was shown that the befouling of plastic particles can result in their settling down and, according to the model based on settling, biofilm growth, ocean depth profiles for light, water density, temperature, salinity, and viscosity, the authors predict that the larger particles start settling last, so these particles are present longer at the ocean surface [61].
At present time a number of studies of microbial biofilms covering the microplastics particles in the marine environment have been conducted. However, almost every study has its own research design and detected parameters, and it is difficult to compare obtained data. Microorganisms in seawater are able to rapidly in a few hours attach to a microplastic particle and colonize its surface [62,63]. It was suggested that the development of a microbial community on microplastics progresses in a certain succession which can be divided into early, mid and late colonization periods [64]. It was observed that representatives of Gammaproteobacteria (genera Pseudomonas, Acinetobacter and Alteromonas) dominated in the microbial community during 0–9 h on acryl samples placed in seawater; meanwhile the members of Alphaproteobacteria (genera Loktanella, Methylobacterium, and Pelagibacter), became predominated during 24–36 h of biofilm formation. However, the same succession was observed for biofilms developed on glass and steel, notwithstanding that bacterial communities were different in biofilms developed on different substances [65]. The study of biofilm development on different substances such as polycarbonate and stainless steel in estuarine water showed that at the beginning the representative form Alphaproteobacteria, mainly the order Rhodobacterales, were abundant members in both stainless steel and polycarbonate biofilms, but after one week of growth the differences were observed [66]. Members of the Gammaproteobacteria, namely from the genus Oleibacter, dominated up to 59.1% among microbial communities in biofilm during the first day of their formation in coastal marine on the samples of polyvinyl chloride, but were quickly replaced by the representatives of Alphaproteobacteria, mainly represented by Rhodobacteraceae, and Flavobacteriia [67]. The composition of microbial communities of biofilms attached to polypropylene and polyvinyl chloride microplastics in seawater varied depending on location and time of exposure, but bacteria from the Alphaproteobacteria class, mainly Rhodobacteraceae, dominated, followed by the Gammaproteobacteria class [68]. Study of bacterial community composition of biofilms developed in seawater on the low- and high-density polyethylene, and polypropylene showed that differences in biofilm composition were stronger after one week of incubation and were not so dramatically at the latest stages of the biofilm formation when representatives of families such as Flavobacteriaceae, Rhodobacteraceae, Planctomycetaceae and Phyllobacteriaceae were abundant on all surfaces [69]. Study of microbial community composition on fragments of polyethylene terephthalate bottles in the North Sea (UK) provided with denaturing gradient gel electrophoresis and sequencing analysis showed that it comprised of bacteria belonging to Bacteroidetes, Proteobacteria and Cyanobacteria [70]. Analysis of microbial communities from biofilms on microplastics (polyethylene, polypropylene, and polystyrene), floating in a bay of the island of Elba in the Mediterranean Sea showed that plastisphere of all MP particles consisted of representatives from the orders Flavobacteriales, Rhodobacterales, Cytophagales, Rickettsiales, Alteromonadales, Chitinophagales, and Oceanospirillales [71]. So, according to data summarized in Table 1, all dominated bacteria in microbial communities in biofilms covered microplastic particles belong to Gram-negative bacteria (Table 1).
There are only several studies devoted to the compositions of biofilms of microplastic found in freshwater systems [72]. Meanwhile, it was shown that microplastics in rivers provide attractive niches for microorganisms which can be transported by rivers to marine environments and serve a vector of bacterial assemblages [73]. According to results of Hoellein with coauthors [74] bacterial communities colonizing different substrates (5 × 5 cm pieces of ceramic tile, glass, aluminum, polyethylene terephthalate, leaf litter, and cardboard) incubated in the river, pond and artificial recirculating laboratory stream were well differentiated from each other, and there were significant differences in the microbial composition of biofilms depending on sites, but there were no differences in the microbial composition of biofilm developed on plastic in comparison with other solid substrates. Dominant bacterial phyla for all biofilms were the same, namely, Proteobacteria, Bacteroidetes, Firmicutes and Verrucomicrobia. However, river biofilms had higher relative abundances of the phylum Acidobacteria and the genera Nitrospira and lower relative abundances of Proteobacteria; meanwhile the biofilms formed in ponds had significantly higher relative abundances of the phyla Proteobacteria, Firmicutes and Chloroflexi. Higher relative abundance of the families Burkholderiaceae, Nitrospiraceae and Nitrosomonadaceae and lower abundance of the family Erythrobacteraceae was observed in the river biofilms.
There are only a few reports about the presence of fungi in biofilms of microplastic in water systems. Species form fungal phylum Ascomycota were found on plastic debris in marine environments [75]. Members of Chytridiomycota, Cryptomycota and Ascomycota dominated in community composition of fungi found on the surface of polyethylene (PE) and polystyrene (PS) particles, which were incubated in different aquatic systems including sea, river, and a wastewater treatment plant [76]. So, the role of fungi in microplastic biofilms is one of the topics for future study.
Additionally, formation of biofilm could lead to change of distribution of microplastics in the water column. So, biofilms covered microplastic particles of polyethylene terephthalate (PET) and polyvinyl chloride (PVC) being placed for 44 days in freshwater systems contained different biomass levels, and increased MPs density which, supposedly, can change MPs particles buoyancy, and stimulate their sinking and transport behavior [77]. A possible influence of biofilm formation on change of polyethylene density and sinking in coastal waters of the North Sea was also shown [78]. However, the authors suggested that formation of bacterial biofilms is not sufficient to induce sinking of microplastic, and only the presence of multicellular organisms can cause sinking of plastic pieces.
The difference between freshwater and marine habitats may play an important role in formation and activities of the microbial biofilm community on microplastics floating in water bodies. However, there is no systematic study to compare microbial communities of microplastic biofilm in different surroundings. Plastic having a long life in comparison with natural surfaces might transport biofilms for long distances [74]. Wind, current, water movement, and weather conditions also affect movement of MPs with adsorbed microbial cells, so, MPs may play the role of vectors for microorganism’s distribution.
4. Microplastics for Human and Animal Pathogens Colonization
It was suggested that drifting plastic debris is a potential vector for spreading of a harmful microalga [79]. A similar question applies to the role of microplastics in relation to microorganisms. It is considered that microorganisms in microplastic-associated biofilms have an important role in pathogens transfer. The microbial biofilm formed on microplastics particles is called “plastisphere”, a term which was proposed by E. Zettler with coauthors [80]. Plastisphere in ocean water had in its composition mainly members of the phylum Proteobacteria (31–45%) which may include pathogenic species [81]. Potentially pathogenic Vibrio spp., Gram-negative bacteria, facultative anaerobes, typically living in saline environment, several species of which could cause foodborne diseases to animals and humans, have been detected on the polypropylene debris from the North Atlantic [80] and on the floating microplastic particles (polyethylene, polypropylene and polystyrene) in the North and Baltic Sea [82]. It was found that bacteria from the family Burkholderiaceae were present in the biofilms covered polyethylene terephthalate placed in the river [74]. The amount of bacteria from order Burkholderiales was much higher on plastic particles such as polyethylene, polypropylene and polystyrene than on natural and inert substrates placed in ambient Baltic bacterioplankton for 2 weeks [83]. Meanwhile some strains from the genus Burkholderia are opportunistic pathogens of humans [84].
Biofilm-coated microplastics which have taste, smell, color, and have a size similar to plankton, are attractive for consumption by aquatic animals especially if food resources in the surrounding environment are scarce [85]. A lot of marine species are harmed and even killed by using plastic debris which poses a threat to their survival. Additionally, microorganisms including pathogens are concentrated on plastic particles, and consumption of such litters could infect the host. A plastic particle consumed by ocean habitat passes through its gut remaining intact and being excreted serves as a source of nutrients to microbiota of biofilm covered the plastic debris including the opportunistic pathogens such as Vibrio spp., that survive in the animal digestive tract [78]. Disposal waters from wastewater treatment plants could be another source of potential pathogens to freshwater bodies [86]. It was shown that microplastic could be carriers of Vibrio in waters with salinity even higher than 26% which poses a risk for marine environments [87]. Bacterial assemblages colonizing microplastic within the river included plastic decomposing organisms and pathogens [73]. Members of the family Campylobacteraceae, microaerophilic Gram-negative bacteria, which can be causative agents of enteritis in humans were found on microplastics in the river in Chicago, USA [77]. Analysis of biofilms of food-related marine macroplastic litter collected from zone of the shoreline on the Northern Ireland that was above water at low tide and underwater at high tide, showed the presence of such Gram-negative potential pathogenic bacteria: Achromobacter xylosoxidans (agent of respiratory infections); Aeromonas salmonicida sp. salmonicida (pathogen in cultured and wild salmonid causing furunculosis); Erwinia rhapontici (an opportunistic plant pathogen); Lelliottia amnigena (infrequent opportunistic in humans causing urinary tract infection); Neisseria flava, N. perflava, N. subflava (rare opportunistic pathogens of humans); Pantoea agglomerans (plant pathogen and opportunistic human pathogen); Providencia rettgeri (urinary tract and eye infections); Pseudomonas fluorescens (rare affects patients with compromised immune systems); Ps. chlororaphis (rarely pathogenic); Ps. oleovorans (rarely pathogenic); Ps. viridiflava (plant pathogen); Rahnella aquatilis (a causal agent of bacteremia in humans), and Serratia fontic (cause of skin and soft tissue infections) [88]. These bacteria strains belonging to ten bacterial genera and thirteen species showed high resistance to the beta-lactam antibiotics (ampicillin, ceftazidime and cefpodoxime) (98.1% resistant) and least resistance to the tetracycline group, minocycline (16.1% resistant) [88]. Thus, among the bacterial species in the microbial community attached to plastic particles, antibiotic-resistant ones could be present. So, these authors conclude that microplastic is the potential threat for the environment, being a means for spreading pathogens into new areas [73,82].
However, there is another point of view on this problem. No higher content of potential pathogens in biofilms covered polystyrene and polyethylene microplastic particles compared to biofilm communities of wood pellets was observed, and it was shown that wooden particles and other natural materials were colonized by Vibrio bacteria even much higher than microplastics [86]. These authors considered that the majority of microorganisms forming the plastisphere are opportunistic colonists, so, their abilities to attach artificial and natural substrates do not differ significantly, and microplastics do not represent a higher threat to surrounding environment than natural particles as possible carriers of potential harmful microorganisms [89]. Studies of bacterial assemblages on polystyrene (PS) comparable with these on glass beads passed through the digestive tract of the lugworm Arenicola marina did not show a distinct enrichment of potential pathogens on PS [90] and no difference in the taxonomic composition of the biofilms on polyamide and chitin after their passage through the gut of the blue mussel Mytilus edulis and not the presence of potential pathogens exclusively on plastic was observed [91]. However, in the areas highly contaminated with microplastics, unpredictable effects on ecosystems may be observed, such as a strong negative impact on marine inhabitants leading to accumulation of microplastic in fish and sea salt.
5. Wastewater Treatment Plant Effluent as a Point Source of Microplastic
According to the role of effluents from wastewater treatment plants (WWTPs) in the transportation of sewage-treated microorganisms into the water environment, there are different positions. It was shown that wastewater treatment plant effluent can be a source of microplastic [73,92]. In experimental studies of McCormick with co-authors [73] microplastic was collected from a river not far from the point where not disinfected effluent from activated sludge plant-treated domestic water was released. It was found that microplastic was covered with dense microbial biofilm, but microscopic study showed the absence of fungal hyphae or algal cells in it. Taxonomic composition of bacterial assemblages covered microplastic differed from those from suspended organic substances and the water column. Bacterial assemblages from microplastic biofilm were less diverse, and pathogens as well as plastic degrading bacteria were more abundant in it.
Microplastics enter WWTPs with influent and exit in sludge or effluent. It was shown for two wastewater treatment plants that microplastic from influent was accumulated in sludge (>99%), and microplastic from sludge had a more diverse spectrum of bacteria species [93,94]. Microplastic from effluent was characterized also by the presence of opportunistic pathogenic bacteria belonging to the genus Acinetobacter, and bacteria was the genera Pseudomonas and Sphingomonas which are considered to be capable of degrading plastic. The authors showed that biofilms on microplastics within sewage have more diverse bacterial assemblages, so WWTPs can be a source of sewage-related microorganisms entered with MPs into the environment. It agrees with the results published [73,89]. Since then, MPs from WWTPs and MPs passing through the guts of aquatic organisms have been hypothesized to play a particular role as vectors for pathogenic microorganisms [59,73].
Biofilms on polyethylene (PE) and polystyrene (PS) samples of microplastic placed in estuary river close to the discharge of effluent from WWTP contained bacteria from the family Enterobacteriaceae [86] that include several potentially pathogenic genera [95]. It was found that content of certain types of bacteria could be associated with its content in the surrounding environment, so, the highest relative abundance of Enterobacteriaceae in biofilms of polystyrene microparticles in water samples with higher abundances of this family [86,95]. The amount of fecal indicator bacteria (Escherichia coli and Enterococci) in the biofilm colonizing polyethylene microplastic were proportionally higher in the environment influenced by sewage discharge [96]. However, it was found that only the low number of potential pathogens were attached to the microplastic particles and relative abundances of Enterobacteriaceae on microplastic were significantly lower in comparison with the surrounding water communities. So, no enrichment with potential pathogens on microplastic particles was observed. However, authors concluded that due to constant long-term discharge of effluent from wastewater treatment plants into freshwater systems, attention should be given to possible increase of levels of microbial pollution [86].
6. Factors Influencing the Microbial Composition of Plastisphere
Once in the aquatic environment, the microplastics become the object for microbial colonization and biofilm development. At the present time there is no clear understanding of what and how different factors affect the composition of biofilms and their function. We shall try to briefly summarize what is known. It was shown that the structure and taxonomic composition of microbial communities on plastic debris depend on plastic material, geographical location and season [59].
Factor 1. Geographic origins. Geographical location has a significant influence on the microbial composition of “Plastisphere”. It was shown that bacterial communities of plastic samples from the North Pacific and North Atlantic subtropical gyres differed between ocean basins [78]. Study of composition of microbial communities from the surfaces of polypropylene and polyvinyl chloride microplastics being incubated for one year in seawater in China showed significant differences depending on geographical locations.
Factor 2. Location of plastic in the water body. Composition of marine plastic bacterial communities showed a great variability depending on plastic locations (sediment, seawater, and resin pellets sampled on the beach) across the Belgian part of the North Sea [97].
Factor 3. Surrounding Environment and seasonal variation. Such factors as salinity and nutrient concentration affect biofilm formation and microbial composition [78,89]. Salinity is considered to be the most essential factor affecting bacterial diversity in the biofilms developed on microplastic particles [87]. Low nutrient levels trigger the attachment to surfaces in many bacterial species, while under high nutrient levels biofilm formation seems less advantageous [98]. The temperature of water obviously plays an important role in formation of the “Plastisphere”, as well as oxygen content in the surrounding environment [99]. It was found that microbial communities on fragments of polyethylene terephthalate bottles in the North Sea (UK) varied from season and location [59,70].
Factor 4. Plastic properties. Physical characteristics of plastic such as density play an important role in its ability to float on the water surface and vertical distribution in the water column. Microplastics with density lower than the density of water are floating on the water surface and could be displaced over long distances; meanwhile microplastics particles with a density higher than the density of water sink in the water column and distribute attached microorganisms in the bulk water. The hydrophobic surface of plastic and its long half-life make it an attractive substrate for microbial colonization [80]. Microplastic has a large specific surface area that enhances microbial cells adhesion on it. Composition of microbial communities depends on the composition of the polymer type they colonize. Significant differences in the community composition of biofilms formed on polyethylene in comparison with polypropylene and polystyrene floating in the Mediterranean Sea were found [71]. It was shown that bacterial communities of biofilm on polystyrene had different bacterial communities than polyethylene and propylene [100]. The same results were obtained previously by Amaral-Zettler with co-authors, who found considerable differences in composition of bacterial communities between polystyrene and polyethylene and between polystyrene and polypropylene [78]. Similar results were obtained by Li and co-authors [87] showed selectiveness of bacterial genera to colonize different types of microplastics such as polyvinyl chloride, polypropylene, polyethylene, polystyrene, and polyurethane. The difference between microbial communities of the biofilms from plastic materials such as polyethylene, polypropylene and polystyrene, native material cellulose and inert glass beads particles placed in ambient Baltic bacterioplankton for 2 weeks was shown [83]. Amount of bacteria from order Burkholderiales was twice higher on plastics, meanwhile the amount of bacteria from genera of Actinobacteria and Cytophagia was much higher on non-plastic materials. Analysis of the presence of pathogenic bacteria in polyethylene terephthalate (PET), polyethylene (PE), and polypropylene (PP) microplastics-associated communities showed their higher on PET, especially potentially pathogenic members of Vibrio, Tenacibaculum, and Escherichia [101].
Transportation and release of microplastics with ballast water may be a serious threat posed by potential pathogens occurring in MPs plastisphere. Firstly, ships’ ballast water could contain bacterial species which are alien or opportunistic pathogens and, in the case of uncontrolled discharge of such water in coastal areas, undesirable microorganisms could be released in the local environment. It was reported that ballast water contained from 0.7 to 39.5% of eubacteria, from 0 to 2.5% of enterobacteria, from 0.2 to 35.8% of Vibrio spp., and from 0 to 2.5% of Escherichia coli from total number of bacterial cells [92]. So, during transportation in the presence of microplastic particles these bacteria could become part of the community of the plastisphere, and their number may increase significantly. Secondly, ballast water transports microplastics with biofilms across different world areas, and incidences of bacterial infections may occur in the recipient country [93].
The hydrophobicity of the substrate surface had a great influence on community structure. The ability of bacterial cells to attach to plastic depends on its structure, level of roughness, and the presence of hydrophilic groups on the structure of the polymer surface [63,102,103]. It was found the positive correlation between hydrophobicity of polymethacrylates and bacterial adhesion [98].
Evaluation of the influence of size (diameter 3 mm and 100 µm) and shape (spherical or irregular) polyethylene and polylactide acid microplastics submerged in sea water for 2 months showed the absence of differences in abundance and diversity in microbial communities, but had an effect on bacterial activity. The higher activity was detected in the biofilm with size 100 µm [104]. It was suggested that bacterial colonization may depend on such intrinsic plastic properties such as pigment content [99].
7. Role Microorganisms of Degradation of Plastic
It is known that most plastics are non-biodegradable. For example, the biodegradation of polythene bags consisted of just 4.2% after 9 months of incubation in mangrove soil, and microbial species involved in the process of biodegradation were representatives of bacterial genera Streptococcus, Staphylococcus, Micrococcus, Moraxella, and Pseudomonas and fungal species of the genus Aspergillus [105]. Plastic is considered as non-degradable material due to (1) high molecular weight which makes impossible transportation of large molecules through the cell membrane of microorganisms; (2) high hydrophobicity which prevent attachment of microbial cells to hydrophilic surfaces; (3) lack of proper functional groups [63,106].
Although many recent studies have focused on the microbial biodegradation of microplastic in water, there are no positive results proving participation of marine microorganisms in this process. However, it was found that sequences from such bacterial genera as Klebsiella, Pseudomonas and Sphingomonas were more abundant in biofilm on microplastic in effluent from WWTP than in sewage, so it can be due to the ability of these microorganism to use microplastics as growth substrates which leads to their degradation [94]. Thus, sequences from the genus Sphingomonas were 10-fold more abundant on microplastic taken from effluent compared to microplastic found in sewage. Meanwhile, bacteria from the genus Sphingomonas can degrade plastic. Sphingomonas paucimobilis isolated from Lake Nakuru, Kenya, degrade polythene and plastics, 37.5% and 17.5%, respectively, after ninety days of degradation [107]. However, several recent studies showed that some bacteria and fungi are capable of degrading certain types of plastic. It was reported that bacterial strain with high cell-surface hydrophobicity Rhodococcus ruber C208 can degrade polyethylene film up to 8%, and the addition of mineral oil increased biodegradation by about 50% [108].
Some authors reported that microorganisms capable of decomposition and consuming inert materials which are very difficult to degrade in the environment could be found in the biofilms on microplastic particles. Thus, microorganisms capable of degrading hydrocarbons with the content up to 17% were found in the biofilms on polypropylene, and polystyrene microplastic particles in the Mediterranean Sea [71]. Microorganisms able to degrade lignin were found in biofilms covered with microplastic particles [70,83]. Very often bacteria capable of degrading hydrocarbon contaminants were present in the microbial communities attached to MRs. Several hydrocarbon-degrading bacteria were identified in the plastisphere of plastic marine debris collected in the North Atlantic [80]. Bacteria of genera Arcobacter and Colwellia which degrade hydrocarbon contaminants within low-temperature marine environments were detected on low-density polyethylene microplastics [108]. Bacteria from the genus Oleibacter were found to be dominated among microbial communities in microplastic biofilm [67]; meanwhile it is known that this genus includes species capable of degrading hydrocarbons [109,110].
Biofilm of microplastic from effluent of WWTP had more representatives of genera Klebsiella, Pseudomonas, and Sphingomonas compared to influent microplastic [94]. Meanwhile, it was found that Pseudomonas and Sphingomonas strains are capable of degrading polycyclic aromatic hydrocarbons [111]; strain Sphingomonas paucimobilis Sy isolated from mangrove sediment degrades dimethyl terephthalate that is used in the manufacturing of polyethylene terephthalate (PET) [112], and bacterial strain Klebsiella pneumoniae was able to degrade high-density polyethylene (HDPE) [113]. The bacterial genus Erythrobacter, known for the ability to utilize polycyclic aromatic hydrocarbons, was found on microplastics [86]. So, the possibility of specific selection of bacterial strains able to break down the microplastic and use it as a source of nutrients in microplastic-associated assemblages was suggested [94]. The data according to occurrence of bacteria belonging to some genera and families in the biofilms covered microplastics which were found in different geographic zones, and the ability of representatives of these genera and families to biodegrade plastic materials or hydrocarbons, are shown in Table 2.
An interesting fact is that marine deuteromycete Zalerion maritimum can use polyethylene as substrate and has been considered as a potential biodegrader [137].
8. Risk of Microplastic for Animals and Humans
Role of microplastic in marine life and its impact on marine inhabitants has been intensively studied during recent years, however a restricted amount of research is done to investigate microplastic impact on human health. It was estimated that humans consume several milligrams of microplastic daily, which consist of up to millions of MP particles a year [138]. Drinking water which comes from freshwater natural resources containing microplastic particles is a source of this contaminant in the human diet, because treatment of water with coagulation-flocculation processes removes only up to 88% of MPs [139]. Tap water in China contains microplastic particles, mostly polyethylene and polypropylene, from 440 ± 275 particles/L [140]. Analysis for microplastic of water taken from different natural water sources showed that its content ranged from 1473 to 3605 particles in 1 L, and MPs content in the treated drinking water varied from 338 to 628 particles in 1 L [139]. PET (polyethylene terephthalate), PP (polypropylene) and PE (polyethylene) consisted of 70% of the total microplastics amount. Generally, microplastics and nanoplastics can enter the human body by indigestion, inhalation and through contact with skin [25]. It was evaluated that one person consumes between 39,000 and 52,000 particles of microplastic with food annually [141], and the consumption of sea food containing microplastic contributes in these numbers.
Microplastic is consumed by various marine organisms from zooplankton to higher trophic levels (fish and marine mammals) and from contaminated prey to predators [142,143]. Trophic transfer, bioaccumulation and biomagnification of microplastic in seafood create a real way for intake of MRs by humans. Plastic debris has been found in fishes, shellfish, mussels and cockles designated for human consumption [144,145,146,147,148].
A lot of microplastic is found in sea salt. The authors [149] proposed to consider sea salt to be an indicator of microplastic pollution of the environment and showed the presence of microplastic particles ranging from 0 to 1674 n/kg in 28 sea salt brands from 16 countries/regions on six continents. The microplastic content, mainly polyethylene terephthalate, polyethylene, and cellophane, in sea salts was from 550 to 681 particles/kg and from 43 to 364 particles in lake salts from supermarkets throughout China [64].
Effects of microplastic consumption on human health are still not well known and need to be investigated. At the present time there is no scientifically proven data that microplastics impact human health. However, the removal of microplastic from organisms is difficult and MPs particles can be accumulated [149]. It is considered that microplastic being consumed by humans can cause inflammation of the gastrointestinal system and in cases when MPs particles serve as carrier for environmental chemicals it can cause toxic effects and organ damage. The presence of microplastics in a human’s body will increase the risk of cancer [82]. However, only microplastics particles with a size of less than 20 µm are capable of penetrating organs and particles with size less than 10 µm could access all organs and cross cell membranes [150,151].
9. Conclusions
The majority of microplastics (MPs), for example, polyethylene, polypropylene, polystyrene and others are accumulated in the upper layers of natural waters due to their low density where they can interact with bacteria inhabiting the bacterioneuston as well as natural organic matter. Bacterioneuston, a thin surface film between the atmosphere and the underlying layers of water in aquatic ecosystems, is characterized by a much higher concentration of bacteria, including pathogenic ones, than the bulk of water. Currently, bacterioneuston and microbial fouling processes of microplastics are being studied independently of each other, while the effect of the bacterioneuston composition on the composition of microplastics microbial assembling is obvious. Bacterioneuston interaction with microplastic can be considered as a key step, influencing the interaction of microplastic particles with environmental pollutants as well as the fate of plastic particles, at first changing their surface properties and disintegration. Thus, a prospective direction of future research includes study of impacts of natural organic matter in microbial life on the surface of microplastic particles which can help to understand their role in the environment and their interaction with other pollutants.
Author Contributions
Conceptualization, V.S. and A.V.; methodology, O.S.; and M.K.; formal analysis, L.K.; writing—original draft preparation, O.S.; writing—review and editing, M.K.; supervision, A.M.; funding acquisition, V.S. and M.K. All authors have read and agreed to the published version of the manuscript.
Funding
Research supported by Latvian-Ukrainian Joint Programme of Scientific and Technological Cooperation project “The microbially-coated microplastics in neustonic water-air interphase and their environmental impacts on aquatic biotopes”.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bellasi, A.; Binda, G.; Pozzi, A.; Galafassi, S.; Volta, P.; Bettinetti, R. Microplastic contamination in freshwater environments: A review, focusing on interactions with sediments and benthic organisms. Environments 2020, 7, 30. [Google Scholar] [CrossRef] [Green Version]
- Plastics Europe. 2020. Available online: https://plasticseurope.org/knowledge-hub/plastics-the-facts-2020 (accessed on 6 November 2021).
- Geyer, R. Production, use, and fate of synthetic polymers. In Plastic Waste and Recycling. Environmental Impact, Societal Issues, Prevention, and Solutions; Letcher, T.M., Ed.; Academic Press: London, UK, 2020; pp. 13–32. [Google Scholar] [CrossRef]
- Rodrigues, J.P.; Duarte, A.C.; Santos-Echeandía, J.; Rocha-Santos, T. Significance of interactions between microplastics and POPs in the marine environment: A critical overview. Trends Analyt. Chem. 2019, 111, 252–260. [Google Scholar] [CrossRef]
- Bowley, J.; Baker-Austin, C.; Porter, A.; Hartnell, R.; Lewis, C. Oceanic hitchhikers—Assessing pathogen risks from marine microplastic. Trends Microbiol. 2021, 29, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.C.; Olsen, Y.; Mitchell, R.P.; Davis, A.; Rowland, S.J.; John, A.W.; McGonigle, D.; Russell, A.E. Lost at sea: Where is all the plastic? Science 2004, 304, 838. [Google Scholar] [CrossRef]
- Frias, J.P.G.L.; Nash, R. Microplastics: Finding a consensus on the definition. Mar. Pollut. Bull. 2019, 138, 145–147. [Google Scholar] [CrossRef]
- Blettler, M.C.; Ulla, M.A.; Rabuffetti, A.P.; Garello, N. Plastic pollution in freshwater ecosystems: Macro-, meso-, and microplastic debris in a floodplain lake. Environ. Monit. Assess. 2017, 189, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Boucher, J.; Friot, D. Primary Microplastics in the Oceans: A Global Evaluation of Sources; IUCN: Gland, Switzerland, 2017; Volume 10. [Google Scholar]
- European Commission. Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions. A European Strategy for Plastics in a Circular Economy; European Commission: Brussels, Belgium, 2018. [Google Scholar]
- Avio, C.G.; Gorbi, S.; Regoli, F. Plastics and microplastics in the oceans: From emerging pollutants to emerged threat. Mar. Environ. Res. 2016, 128, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Thushari, G.G.N.; Senevirathna, J.D.M. Plastic pollution in the marine environment. Heliyon 2020, 6, e04709. [Google Scholar] [CrossRef]
- Li, C.; Busquets, R.; Campos, L.C. Assessment of microplastics in freshwater systems: A review. Sci. Total Environ. 2020, 707, 135578. [Google Scholar] [CrossRef]
- Akdogan, Z.; Guven, B. Microplastics in the environment: A critical review of current understanding and identification of future research needs. Environ. Pollut. 2019, 254, 113011. [Google Scholar] [CrossRef]
- Audrézet, F.; Zaiko, A.; Lear, G.; Wood, S.A.; Tremblay, L.A.; Pochon, X. Biosecurity implications of drifting marine plastic debris: Current knowledge and future research. Mar. Pollut. Bull. 2020, 162, 111835. [Google Scholar] [CrossRef]
- Wagner, M.; Scherer, C.; Alvarez-Muñoz, D.; Brennholt, N.; Bourrain, X.; Buchinger, S.; Fries, E.; Grosbois, C.; Marti, T.; Rodriguez-Mozaz, S.; et al. Microplastics in freshwater ecosystems: What we know and what we need to know. Environ. Sci. Eur. 2014, 26, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eerkes-Medrano, D.; Heather, A.L.; Brian, Q. Microplastics in drinking water: A review and assessment. Curr. Opin. Environ. Sci. Health 2019, 7, 69–75. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.J.; Huang, X.P.; Xiang, L.; Wang, Y.Z.; Li, Y.W.; Li, H.; Cai, Q.Y.; Mo, C.H. Wong, M.H. Source, migration and toxicology of microplastics in soil. Environ. Int. 2020, 137, 105263. [Google Scholar] [CrossRef] [PubMed]
- Rainieri, S.; Barranco, A. Microplastics, a food safety issue? Trends Food Sci. Technol. 2019, 84, 55–57. [Google Scholar] [CrossRef]
- Kumar, M.; Xiong, X.; He, M.; Tsang, D.C.; Gupta, J.; Khan, E.; Bolan, N.S. Microplastics as pollutants in agricultural soils. Environ. Pollut. 2020, 265, 114980. [Google Scholar] [CrossRef]
- Gola, D.; Tyagi, P.K.; Arya, A.; Chauhan, N.; Agarwal, M.; Singh, S.K.; Gola, S. The impact of microplastics on marine environment: A review. Environ. Nanotechnol. Monit. Manag. 2021, 16, 100552. [Google Scholar] [CrossRef]
- Sun, T.; Zhan, J.; Li, F.; Ji, C.; Wu, H. Effect of microplastics on aquatic biota: A hormetic perspective. Environ. Pollut. 2021, 285, 117206. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.; Love, D.C.; Rochman, C.M.; Neff, R.A. Microplastics in seafood and the implications for human health. Curr. Environ. Health Rep. 2018, 5, 375–386. [Google Scholar] [CrossRef] [Green Version]
- Prata, J.C.; da Costa, J.P.; Lopes, I.; Duarte, A.C.; Rocha-Santos, T. Environmental exposure to microplastics: An overview on possible human health effects. Sci. Total Environ. 2020, 702, 134455. [Google Scholar] [CrossRef]
- Yee, M.S.L.; Hii, L.W.; Looi, C.K.; Lim, W.M.; Wong, S.F.; Kok, Y.Y.; Tan, B.K.; Wong, C.Y.; Leong, C.O. Impact of microplastics and nanoplastics on human health. Nanomaterials 2021, 11, 496. [Google Scholar] [CrossRef]
- Bakir, A.; Rowland, S.J.; Thompson, R.C. Competitive sorption of persistent organic pollutants onto microplastics in the marine environment. Mar. Pollut. Bull. 2012, 64, 2782–2789. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Wu, X.; Song, X.; Shi, C.; Zhang, Z. Sorption and desorption of petroleum hydrocarbons on biodegradable and nondegradable microplastics. Chemosphere 2021, 273, 128553. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Liu, F.; Brookes, P.C.; Xu, J. Microplastics play a minor role in tetracycline sorption in the presence of dissolved organic matter. Environ. Pollut. 2018, 240, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Yang, C.; Zhu, Z.; Bai, X.; Ma, J. Adsorption behavior of organic pollutants and metals on micro/nanoplastics in the aquatic environment. Sci. Total Environ. 2019, 694, 133643. [Google Scholar] [CrossRef] [PubMed]
- Torres, F.G.; Dioses-Salinas, D.C.; Pizarro-Ortega, C.I.; De-la-Torre, G.E. Sorption of chemical contaminants on degradable and non-degradable microplastics: Recent progress and research trends. Sci. Total Environ. 2020, 757, 143875. [Google Scholar] [CrossRef] [PubMed]
- Gigault, J.; ter Halle, A.; Baudrimont, M.; Pascal, P.Y.; Gauffre, F.; Phi, T.L.; Phi, T.L.; Hadri, H.E.; Reynaud, B.G.S. Current opinion: What is a nanoplastic? Environ. Pollut. 2018, 235, 1030–1034. [Google Scholar] [CrossRef]
- Lear, G.; Kingsbury, J.M.; Franchini, S.; Gambarini, V.; Maday, S.D.M.; Wallbank, J.A.; Weaver, L.; Pantos, O. Plastics and the microbiome: Impacts and solutions. Environ. Microbiome 2021, 16, 2. [Google Scholar] [CrossRef] [PubMed]
- Naumann, E. Some aspects of the ecology of the limnoplankton, with special reference to the phytoplankton. Sven. Bot. Tidskr. 1919, 13, 129–163. Available online: http://hdl.handle.net/1834/22386 (accessed on 26 October 2021).
- Gladyshev, M. Biophysics of the Surface Microlayer of Aquatic Ecosystems; IWA Publishing: London, UK, 2005; Volume 4, pp. 1–148. [Google Scholar] [CrossRef]
- Hörtnagl, P.; Pérez, M.T.; Sommaruga, R. Living at the border: A community and single-cell assessment of lake bacterioneuston activity. Limnol. Oceanogr. 2010, 55, 1134–1144. [Google Scholar] [CrossRef] [Green Version]
- Zaitsev, Y.P. Marine Neustonologia; Naukova Dumka: Kyiv, Ukraina, 1977; pp. 1–264. [Google Scholar]
- Zavarsin, G.A. Bacterial population of the surface water film in natural reservoirs the Volga delta. Proc. Ins. Microbiol. 1955, 4, 196–201. [Google Scholar]
- Harvey, G.M. Microlayer collection from the sea surface: A new method and initial results. Limnol. Oceanogr. 1966, 11, 608–613. [Google Scholar] [CrossRef]
- Cunliffe, M.; Whiteley, A.S.; Newbold, L.; Oliver, A.; Schäfer, H.; Murrell, J.C. Comparison of bacterioneuston and bacterioplankton dynamics during a phytoplankton bloom in a fjord mesocosm. Appl. Environ. Microbiol. 2009, 75, 7173–7181. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, V. Monitoring the bacterial neuston. In Rapid Methods for Analysis of Biological Materials in the Environment; NATO ASI Series; Stopa, P.J., Bartoszcze, M.A., Eds.; Springer: Dordrecht, The Netherlands, 2000; Volume 30, pp. 67–72. [Google Scholar] [CrossRef]
- Stabnikova, E.V.; Gordienko, A.S.; Ksenofontov, B.S.; Poberezhniy, V.Y.; Stavskaya, T.Z.; Ivanov, V.N. Interaction between the Cells and Water–Air Interface; Naukova Dumka: Kiev, Ukraine, 1991; pp. 1–196. [Google Scholar]
- Gasparovic, B.; Kozarac, Z.; Saliot, A.; Cosovi, B.; Mobius, D. Physicochemical characterization of natural and ex-situ reconstructed sea-surface microlayers. J. Colloid Interface Sci. 1998, 208, 191–202. [Google Scholar] [CrossRef] [Green Version]
- Zhengbin, Z.; Liansheng, L.; Zhijian, W.; Jun, L.; Haibing, D. Physicochemical studies of the sea surface microlayer. J. Colloid Interface Sci. 1998, 204, 294–299. [Google Scholar] [CrossRef]
- Crawford, R.L.; Johnson, L.; Martinson, M. Bacterial enrichment in surface films of freshwater lakes. J. Great Lake Res. 1982, 8, 323–325. [Google Scholar] [CrossRef]
- Stabnikova, E.V.; Gregirchak, N.N.; Ivanov, V.N. The age specificity of interaction between bacterial cells and liquid—Gas interface. Microbiology 1992, 61, 1038–1042. [Google Scholar]
- Stabnikova, E.V.; Ivanov, V.N.; Gregirchak, N.N.; Dulgerov, A.N. Utilization of neuston forms of bacilli for treatment and decontamination of surface waters. Mikrob. Zh. 1993, 55, 88–94. [Google Scholar]
- Franklin, M.P.; McDonald, I.R.; Bourne, D.G.; Owens, N.J.P.; Upstill-Goddard, R.C.; Murrell, J.C. Bacterial diversity in the bacterioneuston (sea surface microlayer): The bacterioneuston through the looking glass. Environ. Microbiol. 2005, 7, 723–736. [Google Scholar] [CrossRef]
- Zäncker, B.; Cunliffe, M.; Engel, A. Bacterial community composition in the sea surface microlayer off the Peruvian coast. Front. Microbiol. 2018, 9, 2699. [Google Scholar] [CrossRef]
- Agogué, H.; Casamayor, E.O.; Bourrain, M.; Obernosterer, I.; Joux, F.; Herndl, G.J.; Lebaron, P. A survey on bacteria inhabiting the sea surface microlayer of coastal ecosystems. FEMS Microbiol. Ecol. 2005, 54, 269–280. [Google Scholar] [CrossRef]
- Fan, J.; Li, H.; Chang, Y.; Wang, X.; Ming, H.; Su, J.; Li, H. Spatial analysis of bacterioneuston and bacterioplankton diversity in the north China Sea. Aquat. Ecosyst. Health Manag. 2018, 21, 1–9. [Google Scholar] [CrossRef]
- Donderski, W.; Walczak, M.; Mudryk, Z.; Kobylinski, M. Neustonic bacteria number, biomass and taxonomy. Pol. J. Environ. Stud. 1999, 8, 137–141. [Google Scholar]
- Fiebig, A. Role of Caulobacter cell surface structures in colonization of the air-liquid interface. J. Bacteriol. 2019, 201, e00064-19. [Google Scholar] [CrossRef] [Green Version]
- Jones, G.W.; Baines, L.; Genthner, F.J. Heterotrophic bacteria of the freshwater neuston and their ability to act as plasmid recipients under nutrient deprived conditions. Microb. Ecol. 1991, 22, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Heinrich-Salmeron, A.; Cordi, A.; Brochier-Armanet, C.; Halter, D.; Pagnout, C.; Abbaszadeh-Fard, E.; Montaut, D.; Seby, F.; Bertin, P.N.; Bauda, P.; et al. Unsuspected diversity of arsenite-oxidizing bacteria as revealed by widespread distribution of the aoxB gene in prokaryotes. App. Environ. Microb. 2011, 77, 4685–4692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kjelleberg, S. Mechanisms of bacterial adhesion at gas-liquid interfaces. In Bacterial Adhesion; Savage, D.C., Fletcher, M., Eds.; Plenum Publication Corp: New York, NY, USA, 1985; pp. 163–194. [Google Scholar]
- Rahlff, J. The virioneuston: A review on viral/bacterial associations at air/water interfaces. Viruses 2019, 11, 191. [Google Scholar] [CrossRef] [Green Version]
- Walczak, M.; Donderski, W.; Mudryk, Z.; Skórczewski, P. Aromatic hydrocarbon decomposition by neustonic bacteria. Part, I. Single-ring hydrocarbon biodegradation. Pol. J. Environ. Stud. 2000, 9, 471–474. [Google Scholar]
- Issac, M.N.; Kandasubramanian, B. Effect of microplastics in water and aquatic systems. Environ. Sci. Pollut. Res. 2021, 28, 19544–19562. [Google Scholar] [CrossRef]
- Oberbeckmann, S.; Loeder, M.G.; Gerdts, G.; Osborn, A.M. Spatial and seasonal variation in diversity and structure of microbial biofilms on marine plastics in Northern European waters. FEMS Microbiol. Ecol. 2014, 90, 478–492. [Google Scholar] [CrossRef]
- Guo, X.; Wang, J.L. The chemical behaviors of microplastics in marine environment: A review. Mar. Pollut. Bull. 2019, 142, 1–14. [Google Scholar] [CrossRef]
- Harrison, P.; Sapp, M.; Schratzberger, M.; Osborn, A.M. Interactions between microorganisms and marine microplastics: A call for research. Mar. Technol. Soc. J. 2011, 45, 12–20. [Google Scholar] [CrossRef]
- Kooi, M.; van Nes, E.H.; Scheffer, M.; Koelmans, A.A. Ups and downs in the ocean: Effects of biofouling on vertical transport of microplastics. Environ. Sci. Technol. 2017, 51, 7963–7971. [Google Scholar] [CrossRef] [Green Version]
- Urbanek, A.K.; Rymowicz, W.; Mirończuk, A.M. Degradation of plastics and plastic-degrading bacteria in cold marine habitats. Appl. Microbiol. Biotechnol. 2018, 102, 7669–7678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.; Shi, H.; Li, L.; Li, J.; Jabeen, K.; Kolandhasamy, P. Microplastic pollution in table salts from China. Environ. Sci. Technol. 2015, 49, 13622–13627. [Google Scholar] [CrossRef]
- Lee, J.W.; Nam, J.H.; Kim, Y.H.; Lee, K.H.; Lee, D.H. Bacterial communities in the initial stage of marine biofilm formation on artificial surfaces. J. Microbiol. 2008, 46, 174–182. [Google Scholar] [CrossRef]
- Jones, P.R.; Cottrell, M.T.; Kirchman, D.L.; Dexter, S.C. Bacterial community structure of biofilms on artificial surfaces in an estuary. Microb. Ecol. 2007, 53, 153–162. [Google Scholar] [CrossRef]
- Pollet, T.; Berdjeb, L.; Garnier, C.; Durrieu, G.; Le Poupon, C.; Misson, B.; Briand, J.F. Prokaryotic community successions and interactions in marine biofilms: The key role of Flavobacteria. FEMS Microbiol. Ecol. 2018, 94, fiy083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Wang, S.; Gao, F.; Li, J.; Zheng, L.; Sun, C.; He, C.; Wang, Z.; Qu, L. Marine microplastic-associated bacterial community succession in response to geography, exposure time, and plastic type in China’s coastal seawaters. Mar. Pollut. Bull. 2019, 145, 278–286. [Google Scholar] [CrossRef]
- Pinto, M.; Langer, T.M.; Huffer, T.; Hofmann, T.; Herndl, G.J. The composition of bacterial communities associated with plastic biofilms differs between different polymers and stages of biofilm succession. PLoS ONE 2019, 14, e0217165. [Google Scholar] [CrossRef] [Green Version]
- Oberbeckmann, S.; Osborn, A.M.; Duhaime, M.B. Microbes on a bottle: Substrate, season and geography influence community composition of microbes colonizing marine plastic debris. PLoS ONE 2016, 11, e0159289. [Google Scholar] [CrossRef]
- Vaksmaa, A.; Knittel, K.; Asbun, A.A.; Goudriaan, M.; Ellrott, A.; Witte, H.J.; Vollmer, I.; Meirer, F.; Lott, C.; Weber, M.; et al. Microbial communities on plastic polymers in the Mediterranean sea. Front. Microbiol. 2021, 12, 673553. [Google Scholar] [CrossRef]
- Harrison, J.P.; Hoellein, T.J.; Sapp, M.; Tagg, A.S.; Ju-Nam, Y.; Ojeda, J.J. Microplastic-associated Biofilms: A comparison of freshwater and marine environments. In Freshwater Microplastics; Wagner, M., Lambert, S., Eds.; Springer: Cham, Switzerland, 2018; Volume 58, pp. 181–201. [Google Scholar] [CrossRef] [Green Version]
- McCormick, A.; Hoellein, J.; Mason, S.A.; Schluep, J.; Kelly, J.J. Microplastic is an abundant and distinct microbial habitat in an urban river. Environ. Sci. Technol. 2014, 48, 11863–11871. [Google Scholar] [CrossRef]
- Hoellein, T.; Rojas, M.; Pink, A.; Gasior, J.; Kelly, J. Anthropogenic litter in urban freshwater ecosystems: Distribution and microbial interactions. PLoS ONE 2014, 9, e98485. [Google Scholar] [CrossRef] [PubMed]
- De Tender, C.; Devriese, L.I.; Haegeman, A.; Maes, S.; Vangeyte, J.; Cattrijsse, A.; Dawyndt, P.; Ruttink, T. Temporal dynamics of bacterial and fungal colonization on plastic debris in the North Sea. Environ. Sci. Technol. 2017, 51, 7350–7360. [Google Scholar] [CrossRef]
- Kettner, M.T.; Rojas-Jimenez, K.; Oberbeckmann, S.; Labrenz, M.; Grossart, H.P. Microplastics alter composition of fungal communities in aquatic ecosystems. Environ. Microbiol. 2017, 19, 4447–4459. [Google Scholar] [CrossRef]
- Miao, L.; Gao, Y.; Adyel, T.M.; Huo, Z.; Liu, Z.; Hoa, J. Effects of biofilm colonization on the sinking of microplastics in three freshwater environments. J. Hazard. Mater. 2021, 413, 125370. [Google Scholar] [CrossRef] [PubMed]
- Amaral-Zettler, L.A.; Zettler, E.R.; Mincer, T.J.; Klaassen, M.A.; Gallager, S.M. Biofouling impacts on polyethylene density and sinking in coastal waters: A macro/micro tipping point? Water Res. 2021, 201, 117289. [Google Scholar] [CrossRef]
- Maso, M.; Garcés, E.; Pages, F.; Camp, J. Drifting plastic debris as a potential vector for dispersing harmful algal bloom (HAB) species. Sci. Mar. 2003, 67, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Zettler, E.R.; Mincer, T.J.; Amaral-Zettler, L.A. Life in the “Plastisphere”: Microbial communities on plastic marine debris. Environ. Sci. Technol. 2013, 47, 7137–7146. [Google Scholar] [CrossRef] [PubMed]
- Dudek, K.L.; Cruz, B.N.; Polidoro, B.; Neuer, S. Microbial colonization of microplastics in the Caribbean Sea. L&O Lett. 2020, 5, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Kirstein, I.V.; Kirmizi, S.; Wichels, A.; Garin-Fernandez, A.; Erler, R.; Löder, M.; Gerdtsa, G. Dangerous hitchhikers? Evidence for potentially pathogenic Vibrio spp. on microplastic particles. Mar. Environ. Res. 2016, 120, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Ogonowski, M.; Motiei, A.; Ininbergs, K.; Hell, E.; Gerdes, Z.; Udekwu, K.I.; Bacsik, Z.; Gorokhova, E. Evidence for selective bacterial community structuring on microplastics. Environ. Microbiol. 2018, 20, 2796–2808. [Google Scholar] [CrossRef] [PubMed]
- Parke, J.L.; Gurian-Sherman, D. Diversity of the Burkholderia cepacia complex and implications for risk assessment of biological control strains. Annu. Rev. Phytopathol. 2001, 39, 225–258. [Google Scholar] [CrossRef]
- Derraik, J.G. The pollution of the marine environment by plastic debris: A review. Mar. Pollut. Bull. 2002, 44, 842–852. [Google Scholar] [CrossRef]
- Oberbeckmann, S.; Kreikemeyer, B.; Labrenz, M. Environmental factors support the formation of specific bacterial assemblages on microplastics. Front. Microbiol. 2018, 8, 2709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.J.; Zhang, Y.; Wu, N.; Zhao, Z.; Xu, W.A.; Ma, Y.Z.; Niu, Z.G. Colonization characteristics of bacterial communities on plastic debris influenced by environmental factors and polymer types in the Haihe estuary of Bohai Bay, China. Environ. Sci. Technol. 2019, 53, 10763–10773. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.E.; Millar, B.C.; Moore, J.E. Antimicrobial resistance (AMR) and marine plastics: Can food packaging litter act as a dispersal mechanism for AMR in oceanic environments? Mar. Pollut. Bull. 2020, 150, 10702. [Google Scholar] [CrossRef]
- Oberbeckmann, S.; Labrenz, M. Marine microbial assemblages on microplastics: Diversity, adaptation, and role in degradation. Ann. Rev. Mar. Sci. 2020, 12, 209–232. [Google Scholar] [CrossRef] [Green Version]
- Kesy, K.; Hentzsch, A.; Klaeger, F.; Oberbeckmann, S.; Mothes, S.; Labrenz, M. Fate and stability of polyamide-associated bacterial assemblages after their passage through the digestive tract of the blue mussel Mytilus edulis. Mar. Pollut. Bull. 2017, 125, 132–138. [Google Scholar] [CrossRef]
- Kesy, K.; Oberbeckmann, S.; Müller, F.; Labrenz, M. Polystyrene influences bacterial assemblages in Arenicola marina-populated aquatic environments in vitro. Environ. Pollut. 2016, 219, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, V. Bacteriological monitoring of ships’ ballast water in Singapore and its potential importance for the management of coastal ecosystems. WIT Trans. Biomed. Health 2006, 10, 5. [Google Scholar] [CrossRef] [Green Version]
- Naik, R.K.; Naik, M.M.; D’Costa, P.M.; Shaikh, F. Microplastics in ballast water as an emerging source and vector for harmful chemicals, antibiotics, metals, bacterial pathogens and HAB species: A potential risk to the marine environment and human health. Mar. Pollut. Bull. 2019, 149, 110525. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.J.; London, M.G.; McCormick, A.R.; Rojas, M.; Scott, J.W.; Hoellein, T.J. Wastewater treatment alters microbial colonization of microplastics. PLoS ONE 2021, 16, e0244443. [Google Scholar] [CrossRef] [PubMed]
- Octavia, S.; Lan, R. The family Enterobacteriaceae. In The Prokaryotes: Gammaproteobacteria; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 225–286. [Google Scholar] [CrossRef]
- Pazos, R.S.; Suárez, J.C.; Gómez, N. Study of the plastisphere: Biofilm development and presence of faecal indicator bacteria on microplastics from the Río de la Plata estuary. Ecosistemas 2020, 29, 2069. [Google Scholar] [CrossRef]
- Harrison, J.P.; Schratzberger, M.; Sapp, M.; Osborn, A.M. Rapid bacterial colonization of low-density polyethylene microplastics in coastal sediment microcosms. BMC Microbiol. 2014, 14, 232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanni, O.; Chang, C.Y.; Anderson, D.G.; Langer, R.; Davies, M.C.; Williams, P.M.; Williams, P.; Alexander, M.R.; Hook, A.L. Bacterial attachment to polymeric materials correlates with molecular flexibility and hydrophilicity. Adv. Healthc. Mater. 2015, 4, 695–701. [Google Scholar] [CrossRef]
- De Tender, C.A.; Devriese, L.I.; Haegeman, A.; Maes, S.; Ruttink, T.; Dawyndt, P. Bacterial community profiling of plastic litter in the Belgian part of the North Sea. Environ. Sci. Technol. 2015, 49, 9629–9638. [Google Scholar] [CrossRef] [PubMed]
- Frere, L.; Maignien, L.; Chalopin, M.; Huvet, A.; Rinnert, E.; Morrison, H.; Kerninon, S.; Cassone, A.L.; Lambert, C.; Reveillaud, J.; et al. Microplastic bacterial communities in the Bay of Brest: Influence of polymer type and size. Environ. Pollut. 2018, 242 Pt A, 614–625. [Google Scholar] [CrossRef] [Green Version]
- Hou, D.; Hong, M.; Wang, Y.; Dong, P.; Cheng, H.; Yan, H.; Yao, Z.; Li, D.; Wang, K.; Zhang, D. Assessing the risks of potential bacterial pathogens attaching to different microplastics during the summer–autumn period in a mariculture cage. Microorganisms 2021, 9, 1909. [Google Scholar] [CrossRef] [PubMed]
- Donlan, R.M. Biofilms: Microbial life on surfaces. Emerg. Infect. Dis. 2002, 8, 881–890. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Peng, C.; Li, H.; Zhang, P.; Liu, X. The impact of microplastic-microbe interactions on animal health and biogeochemical cycles: A mini-review. Sci. Total Environ. 2021, 773, 145697. [Google Scholar] [CrossRef]
- Cheng, J.; Jacquin, J.; Conan, P.; Barbe, V.; George, M.; Fabre, P.; Bruzaud, S.; Halle, A.T.; Meistertzheim, A.L.; Ghiglione, J.F. Relative influence of plastic debris size and shape, chemical composition and phytoplankton-bacteria interactions in driving seawater plastisphere abundance, diversity and activity. Front. Microbiol. 2021, 11, 610231. [Google Scholar] [CrossRef]
- Kathiresan, K. Polythylene and plastics-degrading microbes in an Indian mangrove soil. Rev. Biol. Trop. 2003, 51, 629–633. [Google Scholar]
- Wilkes, R.A.; Aristilde, L. Degradation and metabolism of synthetic plastics and associated products by Pseudomonas sp.: Capabilities and challenges. J. Appl. Microbiol. 2017, 123, 582–593. [Google Scholar] [CrossRef] [Green Version]
- Wanjohi, L.; Mwamburi, L.; Too, E.; Kosgei, J. Bacterial Diversity in Lake Nakuru and their Potential Utilization in Agriculture and Environment Management. In Proceedings of the 27th Soil Science Society of East Africa and the 6th African Soil Science Society, Nakuru, Kenya, 20–25 October 2013. [Google Scholar]
- Gilan, I.; Hadar, Y.; Sivan, A. Colonization, biofilm formation and biodegradation of polyethylene by a strain of Rhodococcus ruber. Appl. Microbiol. Biotechnol. 2004, 65, 97–104. [Google Scholar] [CrossRef]
- Liu, J.; Zheng, Y.; Lin, H.; Wang, X.; Li, M.; Liu, Y.; Yu, M.; Zhao, M.; Pedentchouk, N.; Lea-Smith, D.J.; et al. Proliferation of hydrocarbon-degrading microbes at the bottom of the Mariana Trench. Microbiome 2019, 7, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teramoto, M.; Ohuchi, M.; Hatmanti, A.; Darmayati, Y.; Widyastuti, Y.; Harayama, S.; Fukunaga, Y. Oleibacter marinus gen. nov., sp. nov., a bacterium that degrades petroleum aliphatic hydrocarbons in a tropical marine environment. Int. J. Syst. Evol. Microbiol. 2011, 61, 375–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bastiaens, L.; Springael, D.; Wattiau, P.; Harms, H.; deWachter, R.; Verachtert, H.; Diels, L. Isolation of adherent polycyclic aromatic hydrocarbon (PAH)-degrading bacteria using PAH-sorbing carriers. Appl. Environ. Microbiol. 2000, 66, 834–1843. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Gu, J.D.; Yao, J.H. Degradation of dimethyl terephthalate by Pasteurella multocida Sa and Sphingomonas paucimobilis Sy isolated from mangrove sediment. Int. Biodeter. Biodegr. 2005, 56, 158–165. [Google Scholar] [CrossRef]
- Awasthi, S.; Srivastava, P.; Singh, P.; Tiwary, D.; Mishra, P.K. Biodegradation of thermally treated high-density polyethylene (HDPE) by Klebsiella pneumoniae CH001. 3 Biotech 2017, 75, 332. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Singh, A.K.; Bhatt, K. Biodegradation of polyethylene by bacteria isolated from soil. Int. J. Res. Dev. Pharm. Life Sci. 2016, 5, 2056–2062. [Google Scholar]
- Pathak, V.M.; Navneet. Review on the current status of polymer degradation: A microbial approach. Bioresour. Bioprocess. 2017, 4, 15. [Google Scholar] [CrossRef]
- Wu, X.; Pan, J.; Li, M.; Li, Y.; Bartlam, M.; Wang, Y. Selective enrichment of bacterial pathogens by microplastic biofilm. Water Res. 2019, 165, 114979. [Google Scholar] [CrossRef]
- Asmita, K.; Shubhamsingh, T.; Tejashree, S. Isolation of plastic degrading microorganisms from soil samples collected at various locations in Mumbai, India. Curr. World Environ. 2015, 4, 77–85. [Google Scholar]
- Curren, E.; Leong, S.C.Y. Profiles of bacterial assemblages from microplastics of tropical coastal environments. Sci. Total Environ. 2019, 655, 313–320. [Google Scholar] [CrossRef]
- Tamnou, E.M.; Arfao, A.T.; Nougang, M.; Metsopkeng, C.; Ewoti, O.V.N.; Moungang, L.M. Nana, P.A.; Takang-Etta, L.R.A.; Perrière, F.; Sime-Ngando, T.; et al. Biodegradation of polyethylene by the bacterium Pseudomonas aeruginosa in acidic aquatic microcosm and effect of the environmental temperature. Environ. Chall. 2021, 3, 100056. [Google Scholar] [CrossRef]
- Kesy, K.; Oberbeckmann, S.; Kreikemeyer, B.; Labrenz, M. Spatial environmental heterogeneity determines young biofilm assemblages on microplastics in Baltic Sea. Front. Microbiol. 2019, 10, 1665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, B.; Olanrewaju-Kehinde, D.; Popoola, O.; James, E. Degradation of plastic and polythene materials by some selected microorganisms isolated from soil. World Appl. Sci. J. 2015, 33, 1888–1891. [Google Scholar] [CrossRef]
- Kertesz, M.A.; Kawasaki, A.; Stolz, A. Aerobic hydrocarbon-degrading alphaproteobacteria: Sphingomonadales. In Taxonomy, Genomics and Ecophysiology of Hydrocarbon-Degrading Microbes; McGenity, T.J., Ed.; Springer: Cham, Switzerland, 2017; pp. 1–21. [Google Scholar]
- Gao, X.; Gao, W.; Cui, Z.; Han, B.; Yang, P.; Sun, C.; Zheng, L. Biodiversity and degradation potential of oil-degrading bacteria isolated from deep-sea sediments of South Mid-Atlantic Ridge. Mar. Pollut. Bull. 2015, 97, 373–380. [Google Scholar] [CrossRef]
- Matallana-Surget, S.; Villette, C.; Intertaglia, L.; Joux, F.; Bourrain, M.; Lebaron, P. Response to UVB radiation and oxidative stress of marine bacteria isolated from South Pacific Ocean and Mediterranean Sea. J. Photochem. Photobiol. 2012, B117, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Atanasova, N.; Stoitsova, S.; Paunova-Krasteva, T.; Kambourova, M. Plastic degradation by extremophilic bacteria. Int. J. Mol. Sci. 2021, 22, 5610. [Google Scholar] [CrossRef]
- Ozaki, S.; Kishimoto, N.; Fujita, T. Change in the predominant bacteria in a microbial consortium cultured on media containing aromatic and saturated hydrocarbons as the sole carbon source. Microbes Environ. 2007, 22, 128–135. [Google Scholar] [CrossRef] [Green Version]
- Sivan, A.; Szanto, M.; Pavlov, V. Biofilm development of the polyethylene degrading bacterium Rhodococcus ruber. Appl. Microbiol. Biotechnol. 2006, 72, 346–352. [Google Scholar] [CrossRef]
- Bacosa, H.P.; Erdner, D.L.; Rosenheim, B.E.; Kiley, P.S.; Shetty, P.; Seitz, K.W.; Baker, B.J.; Liu, Z. Hydrocarbon degradation and response of seafloor sediment bacterial community in the northern Gulf of Mexico to light Louisiana sweet crude oil. ISME J. 2018, 12, 2532–2543. [Google Scholar] [CrossRef]
- Dang, H.; Li, T.; Chen, M.; Huang, G. Cross-Ocean distribution of Rhodobacterales bacteria as primary surface colonizers in temperate coastal marine waters. Appl. Environ. Microbiol. 2008, 74, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Mor, R.; Sivan, A. Biofilm formation and partial biodegradation of polystyrene by the actinomycetes Rhodococcus ruber. Biodegradation 2008, 19, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Al-Amshawee, S.; Yunus, M.Y.B.M.; Vo, D.V.N.; Yunus, N.H. Biocarriers for biofilm immobilization in wastewater treatments: A review. Environ. Chem. Lett. 2020, 18, 1925–1945. [Google Scholar] [CrossRef]
- Trivedi, P.; Hasan, A.; Akhtar, S.; Siddiqui, M.H.; Sayeed, U.; Khan, M.K.A. Role of microbes in degradation of synthetic plastics and manufacture of bioplastics. J. Chem. Pharm. Res. 2016, 8, 211–216. [Google Scholar]
- Brakstad, O.G.; Bonaunet, K. Biodegradation of petroleum hydrocarbons in seawater at low temperatures (0–5°C) and bacterial communities associated with degradation. Biodegradation 2006, 17, 71–82. [Google Scholar] [CrossRef]
- Thompson, H.F.; Summers, S.; Yuecel, R.; Gutierrez, T. hydrocarbon-degrading bacteria found tightly associated with the 50–70 μm cell-size population of eukaryotic phytoplankton in surface waters of a Northeast Atlantic region. Microorganisms 2020, 8, 1955. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, V.; Natarajan, K.; Hemambika, B.; Ramesh, N.; Sumathi, C.S.; Kottaimuthu, R.; Kannan, V.R. High-density polyethylene (HDPE)-degrading potential bacteria from marine ecosystem of Gulf of Mannar, India. Lett. Appl. Microbiol. 2010, 51, 205–211. [Google Scholar] [CrossRef]
- Satlewal, A.; Soni, R.; Zaidi, M.G.H.; Shouche, Y.; Goel, R. Comparative biodegradation of HDPE and LDPE using an indigenously developed microbial consortium. J. Microbiol. Biotechnol. 2008, 18, 477–482. [Google Scholar]
- Paco, A.; Duarte, K.; da Costa, J.P.; Santos, P.S.M.; Pereira, R.; Pereira, M.E.; Freitas, A.C.; Duarte, A.C.; Rocha-Santos, T.A.P. Biodegradation of polyethylene microplastics by the marine fungus Zalerion maritimum. Sci. Total Environ. 2017, 586, 10–15. [Google Scholar] [CrossRef]
- Kannan, K.; Vimalkumar, K. A review of human exposure to microplastics and insights into microplastics as obesogens. Front. Endocrinol. 2021, 12, 724989. [Google Scholar] [CrossRef] [PubMed]
- Pivokonsky, M.; Cermakova, L.; Novotna, K.; Peer, P.; Cajthaml, T.; Janda, V. Occurrence of microplastics in raw and treated drinking water. Sci. Total Environ. 2018, 643, 644–651. [Google Scholar] [CrossRef]
- Tong, H.; Jiang, Q.; Hu, X.; Zhong, X. Occurrence and identification of microplastics in tap water from China. Chemosphere 2020, 252, 126493. [Google Scholar] [CrossRef]
- Cox, K.D.; Covernton, G.A.; Davies, H.L.; Dower, J.F.; Juanes, F.; Dudas, S.E. Human consumption of microplastics. Environ. Sci. Technol. 2019, 53, 7068–7074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, E.; Piazza, V.; Lavorano, S.; Faimali, M.; Garaventa, F.; Gambardella, C. Trophic transfer of microplastics from copepods to jellyfish in the marine environment. Front. Environ. Sci. 2020, 4, 158. [Google Scholar] [CrossRef]
- Farrell, P.; Nelson, K. (Trophic level transfer of microplastic: Mytilus edulis (L.) to Carcinus maenas (L.). Environ. Pollut. 2013, 177, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Rochman, C.M.; Tahir, A.; Williams, S.L.; Baxa, D.V.; Lam, R.; Miller, J.T.; The, F.C.; Werorilangi, S.; Teh, S.J. Anthropogenic debris in seafood: Plastic debris and fibers from textiles in fish and bivalves sold for human consumption. Sci. Rep. 2015, 5, 14340. [Google Scholar] [CrossRef]
- Davidson, K.; Dudas, S.E. Microplastic ingestion by wild and cultured Manila clams (Venerupis philippinarum). Arch. Environ. Contam. Toxicol. 2016, 71, 147–156. [Google Scholar] [CrossRef]
- Giani, D.; Baini, M.; Galli, M.; Casini, S.; Fossi, M.C. Microplastics occurrence in edible fish species (Mullus barbatus and Merluccius merluccius) collected in three different geographical sub-areas of the Mediterranean Sea. Mar. Pollut. Bull. 2019, 140, 129–137. [Google Scholar] [CrossRef]
- Gurjar, U.R.; Xavier, M.; Nayak, B.B.; Ramteke, K.; Deshmukhe, G.; Jaiswar, A.K.; Shukla, S. Microplastics in shrimps: A study from the trawling grounds of north eastern part of Arabian Sea. Environ. Sci. Pollut. Res. 2021, 28, 1–11. [Google Scholar] [CrossRef]
- Hermabessiere, L.; Paul-Pont, I.; Cassone, A.L.; Himber, C.; Receveur, J.; Jezequel, R.; El Rakwe, M.; Rinnert, E.; Rivière, G.; Lambert, C.; et al. Microplastic contamination and pollutant levels in mussels and cockles collected along the channel coasts. Environ. Pollut. 2019, 250, 807–819. [Google Scholar] [CrossRef]
- Kim, J.S.; Lee, H.J.; Kim, S.K.; Kim, H.J. Global pattern of microplastics (MPs) in commercial food-grade salts: Sea salt as an indicator of seawater MP pollution. Environ. Sci. Technol. 2018, 52, 12819–12828. [Google Scholar] [CrossRef]
- Campanale, C.; Massarelli, C.; Savino, I.; Locaputo, V.; Uricchio, V.F. A Detailed review study on potential effects of microplastics and additives of concern on human health. Int. J. Environ. Res. Public Health 2020, 17, 1212. [Google Scholar] [CrossRef] [Green Version]
- Vaseasta, A.; Ivanov, V.; Stabnikov, V.; Marinin, A. Environmental safety and security investigations of neustonic microplastic aggregates near water-air interphase. Pol. J. Environ. Stud. 2021, 30, 3457–3469. [Google Scholar] [CrossRef]
Figure 1.
Concept of interaction between microplastic particles, bacterioneuston, considering its location in water column and natural organic matter.
Figure 1.
Concept of interaction between microplastic particles, bacterioneuston, considering its location in water column and natural organic matter.


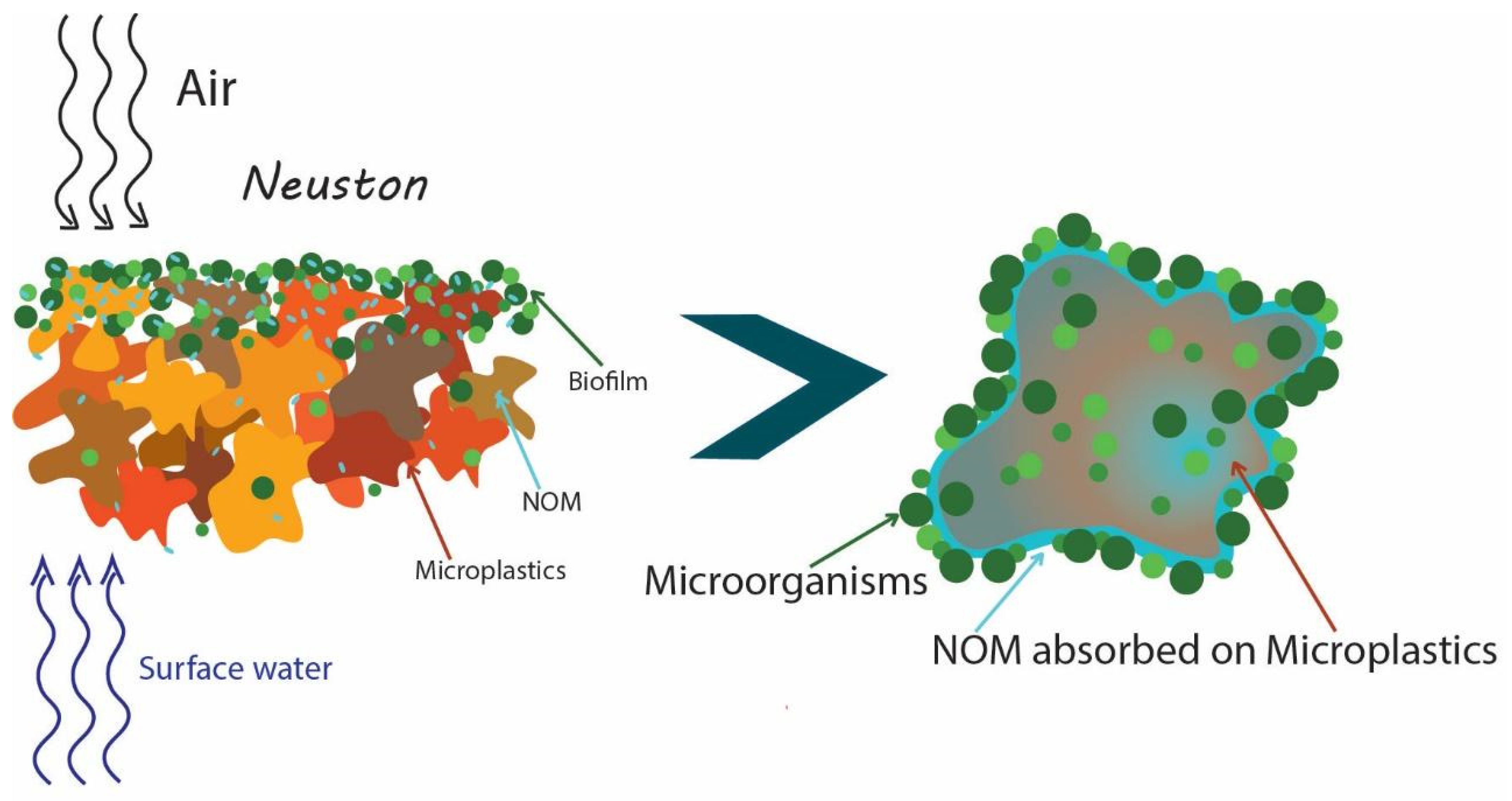{kind=link}
Table 1.
Dominated microorganisms in biofilms on microplastics found in salinity waters.
| Microorganisms Belonging to | Characteristics | Type of Plastic, Location | Time of Biofilm Development | References |
|---|---|---|---|---|
| Gammaproteobacteria genera Pseudomonas Acinetobacter Alteromonas | Gram-negative | Acryl in seawater | Dominated during 0–9 h of biofilm formation | [65] |
| Gram-negative | ||||
| Gram-negative | ||||
| Gram-negative | ||||
| Alphaproteobacteria | Gram-negative | Predominated during 24–36 h of biofilm formation | ||
| genera Loktanella | Gram-negative | |||
| Methylobacterium | Gram-negative | |||
| Pelagibacter | Gram-negative | |||
| Gammaproteobacteria namely genus Oleibacter | Gram-negative | Polyvinyl chloride in coastal marine | Dominated up to 59.1% in the first hours, but | [67] |
| Gram-negative | ||||
| mainly genera Alphaproteobacteria | Gram-negative | were quickly replaced by | ||
| Rhodobacteraceae | Gram-negative | |||
| Flavobacteriia | Gram-negative | |||
| Alphaproteobacteria | Gram-negative | Polypropylene and polyvinyl chloride in seawater | Formed biofilm | [68] |
| mainly genus Rhodobacteraceae | Gram-negative | |||
| followed by the Gammaproteobacteria | Gram-negative | |||
| Flavobacteriaceae | Gram-negative | Polyethylene, polypropylene in seawater | Latest stages of the biofilm formation | [69] |
| Rhodobacteraceae | Gram-negative | |||
| Phyllobacteriaceae | Gram-negative | |||
| Planctomycetaceae | Gram-negative | |||
| Bacteroidetes | Gram-negative | Polyethylene terephthalate in the North Sea | Formed biofilm | [70] |
| Proteobacteria | Gram-negative | |||
| Cyanobacteria | Gram-negative | |||
| Alphaproteobacteria | Gram-negative | Polycarbonate in estuarine water | At the beginning the most abundant members | [66] |
| mainly the Rhodobacterales | Gram-negative | |||
| Flavobacteriales | Gram-negative | Polyethylene, polypropylene, polystyrene in the Mediterranean Sea | Formed biofilm | [71] |
| Rhodobacterales | Gram-negative | |||
| Cytophagales | Gram-negative | |||
| Rickettsiales | Gram-negative | |||
| Alteromonadales | Gram-negative | |||
| Chitinophagales | Gram-negative | |||
| Oceanospirillales | Gram-negative |
Table 2.
Microorganisms found abundant on microplastics and the ability of representatives of these genera to degrade plastic or hydrocarbons.
Table 2.
Microorganisms found abundant on microplastics and the ability of representatives of these genera to degrade plastic or hydrocarbons.
| Microorganisms Belonging To | Type of Plastic, Location | Ref. | Microorganisms Belonging To | Degrade Plastics or Hydrocarbons | Ref. |
|---|---|---|---|---|---|
| Pseudomonas | MP, WWTP, Illinois, USA | [94] | Pseudomonas | PAHs | [111] |
| Pseudomonas sp. | acryl | Pseudomonas | PE | [114] | |
| Pseudomonas | MP, river, Chicago, USA | [73] | Pseudomonas | Polyester | [115] |
| Pseudomonas cepacia | PE, Ps, Lake Nakuru, Kenya | [107] | Pseudomonas cepacia | Plastic | [107] |
| Pseudomonas | PVC, water from Haihe River, China | [116] | Pseudomonas aeruginosa | PET, PS | [117] |
| Pseudomonas | MP, the Singapore Strait | [118] | Pseudomonas aeruginosa | PE | [119] |
| Pseudomonas | PE, PS, Baltic Sea | [120] | Pseudomonas fluorescens | PE | [121] |
| Sphingomonas | MP, WWTP, Illinois, USA | [94] | Sphingomonas | DMT | [112] |
| Sphingomonas paucimobilis | PE, Ps, Lake Nakuru, Kenya | [107] | Sphingomonas paucimobilis | Ps, PE | [107] |
| Sphingomonadaceae | PE, PS, Baltic Sea | [120] | Sphingomonadaceae | hydrocarbons | [122] |
| Erythrobacter | PS, PE, Baltic Sea | [86] | Erythrobacter | PAHs | [123] |
| Erythrobacter | Ps, Mediterranean Sea | [89,124] | Erythrobacter | hydrocarbons | [125] |
| Erythrobacter | MP, the Singapore Strait | [118] | Erythrobacter | hydrocarbons | [118] |
| Hyphomonadaceae | Ps, the North Atlantic | [80] | Hyphomonadaceae | hydrocarbons | [126] |
| Klebsiella | MP, WWTP, Illinois, USA | [94] | Klebsiella pneumoniae | HDPE | [113] |
| Rhodobacteraceae | Ps, North Atlantic | [80] | Rhodococcus ruber | PE | [127] |
| Rhodobacteraceae | Ps, North Sea | [75] | Rhodobacteraceae | hydrocarbons | [128] |
| Rhodobacterales | PVC, Pacific Ocean | [129] | Rhodococcus ruber | PS | [130] |
| Rhodobacteraceae | PE, PP, PS, Mediterranean Sea | [71] | Rhodococcus ruber C208 | PE | [108] |
| Alcaligenes sp. | PE, Ps, Lake Nakuru, Kenya | [107] | Alcaligenes sp. | PE | [107] |
| Alcaligenes faecalis | Polymeric materials, WWTP | [131] | Alcaligenes faecalis | PCL | [132] |
| Oleibacter | Ps, coastal marine | [67] | Oleibacter | hydrocarbons | [109,110] |
| Arcobacter | LDPE, Humber Estuary, UK | [97] | Arcobacter | hydrocarbons | [133] |
| Colwellia | LDPE, Humber Estuary, UK | [97] | Colwellia | hydrocarbons | [128,134] |
| Arthrobacter sp. | LDPE, Caribbean Sea | [81] | Arthrobacter sp. | LDPE, HDPE | [135,136] |
MP—microplastics; Ps—plastics; PE—polyethylene (polythene); PP—polypropylene, PS—polystyrene; PET—polyethylene terephthalate; PVC—polyvinyl chloride; DMT—dimethyl terephthalate; PAHs—polycyclic aromatic hydrocarbons; PCL—polycaprolactone; LDPE—low-density polyethylene microplastics; HDPE—high- density polyethylene microplastics.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Stabnikova, O.; Stabnikov, V.; Marinin, A.; Klavins, M.; Klavins, L.; Vaseashta, A. Microbial Life on the Surface of Microplastics in Natural Waters. Appl. Sci. 2021, 11, 11692. https://0-doi-org.brum.beds.ac.uk/10.3390/app112411692
AMA Style
Stabnikova O, Stabnikov V, Marinin A, Klavins M, Klavins L, Vaseashta A. Microbial Life on the Surface of Microplastics in Natural Waters. Applied Sciences. 2021; 11(24):11692. https://0-doi-org.brum.beds.ac.uk/10.3390/app112411692
Chicago/Turabian StyleStabnikova, Olena, Viktor Stabnikov, Andriy Marinin, Maris Klavins, Linards Klavins, and Ashok Vaseashta. 2021. "Microbial Life on the Surface of Microplastics in Natural Waters" Applied Sciences 11, no. 24: 11692. https://0-doi-org.brum.beds.ac.uk/10.3390/app112411692
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.