Low Effectiveness of the Introduction of pmaxGFP into Primary Human Coronary Endothelial Cells Using Cell-Penetrating Peptides and Nuclear-Localization Sequences in Non-Covalent Interactions

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
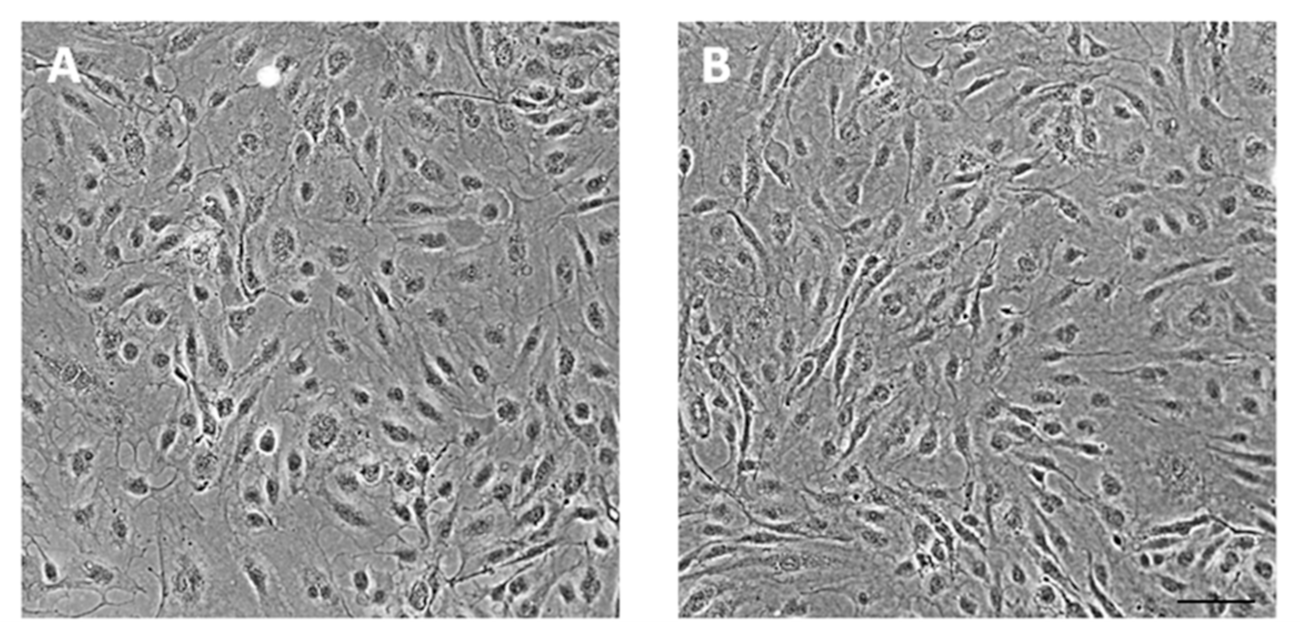
2.1. Cell Culture
2.2. Formation of Electrostatic Interactions between Plasmid and Penetrating Proteins
2.3. Results Evaluation
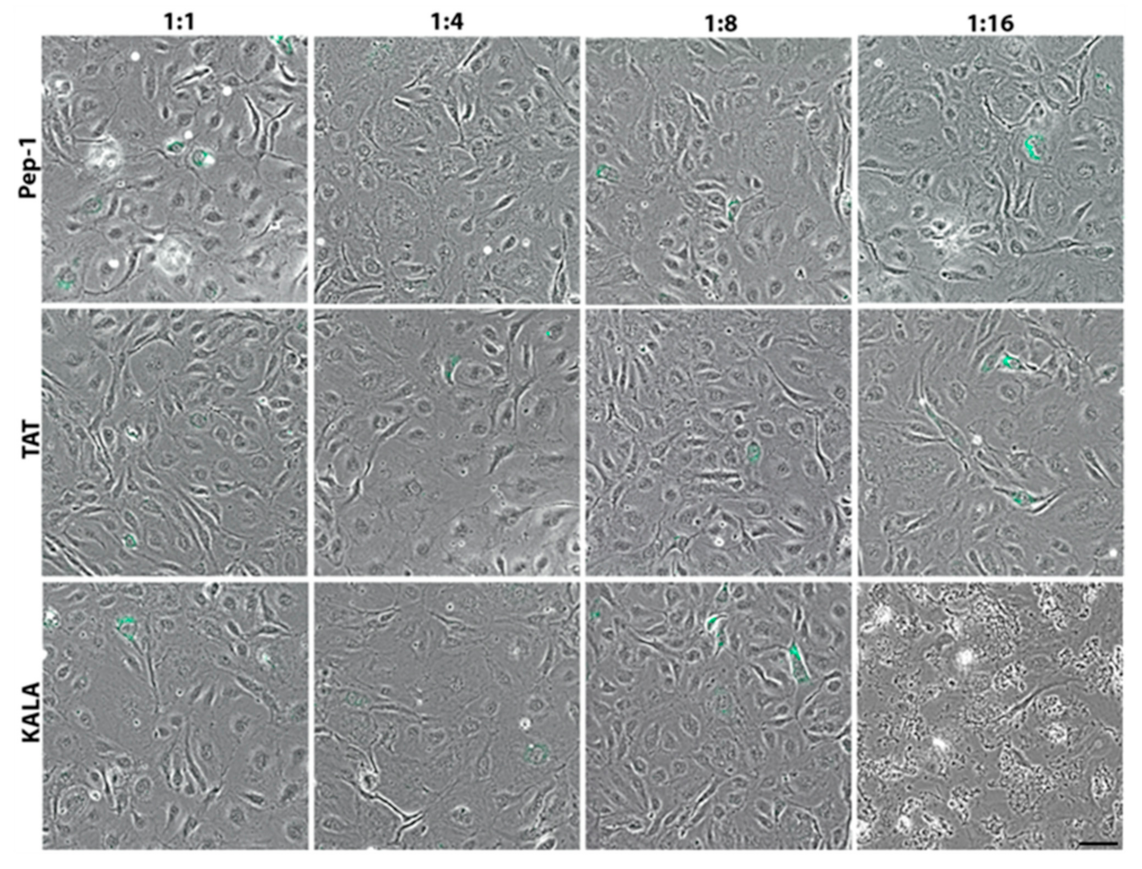
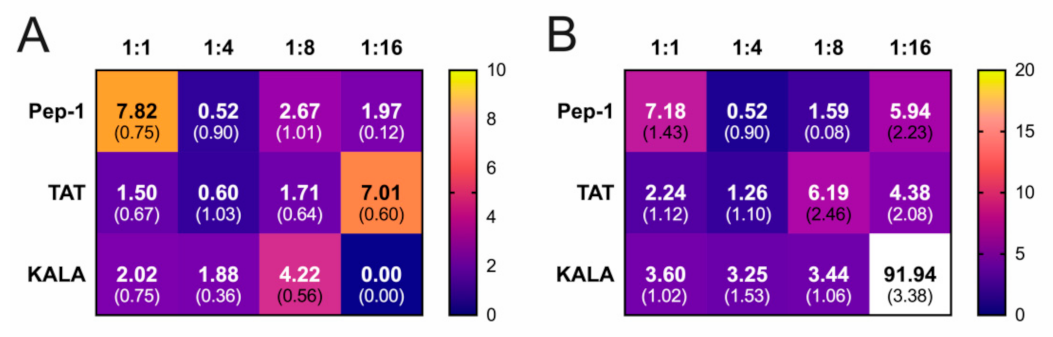
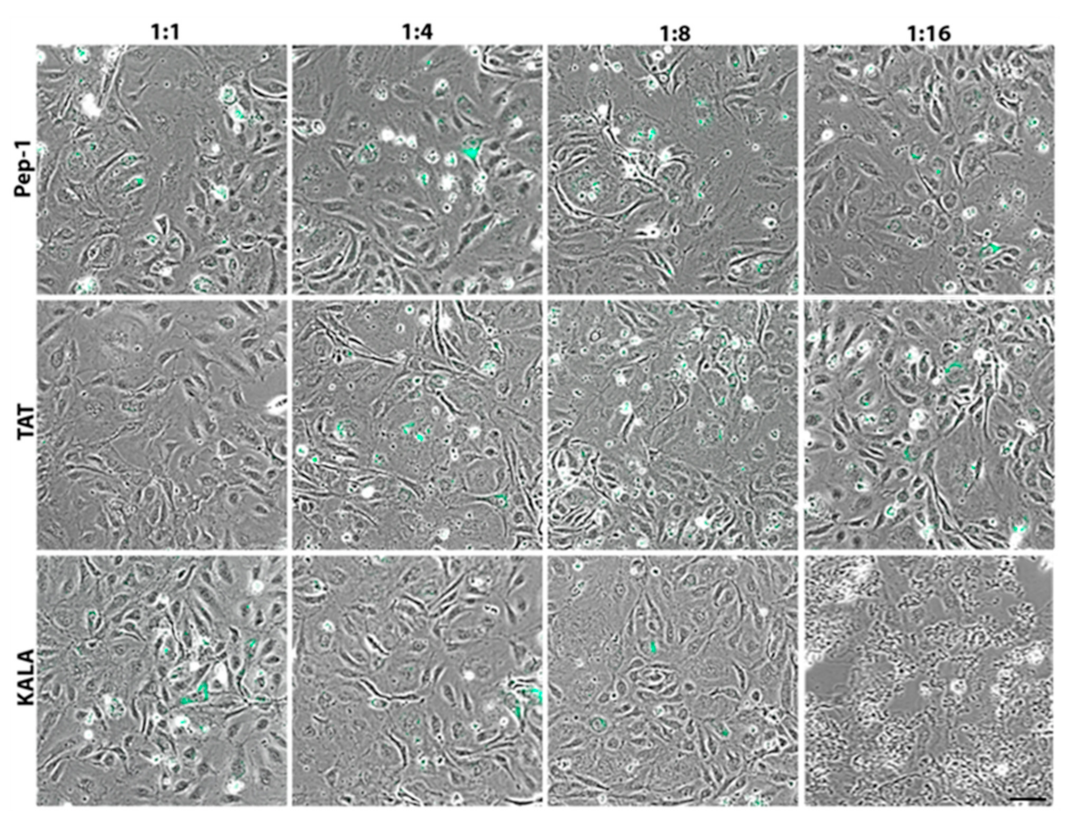
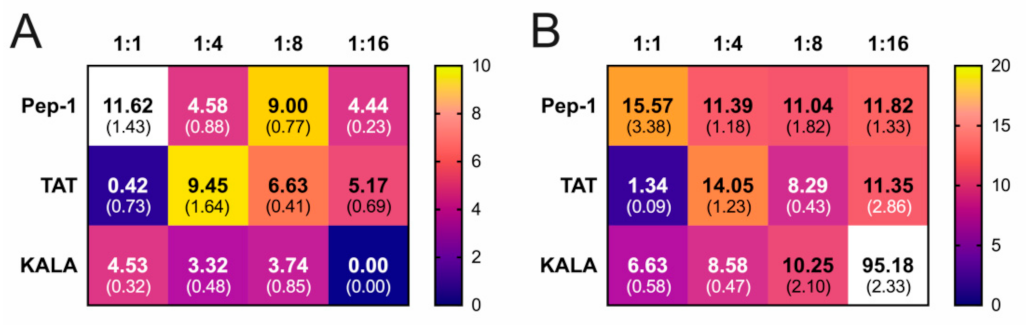
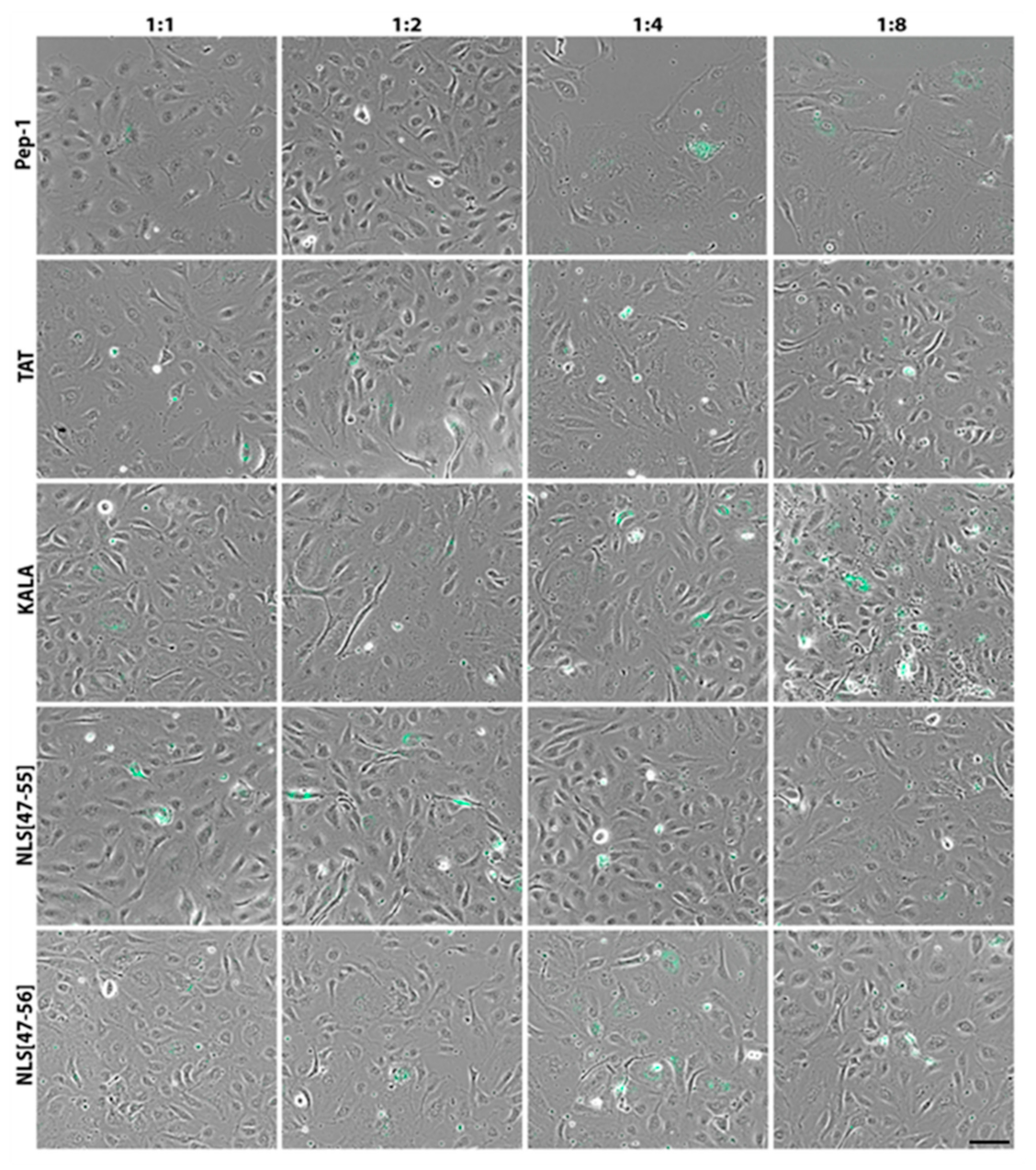
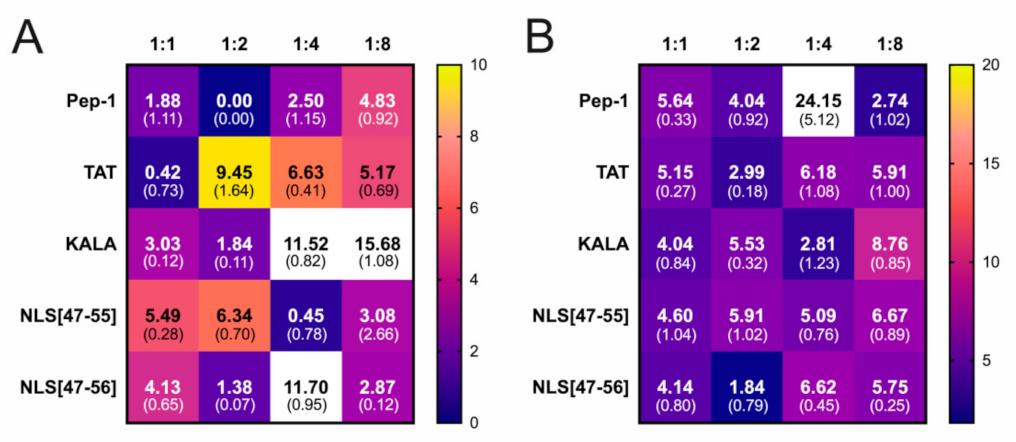
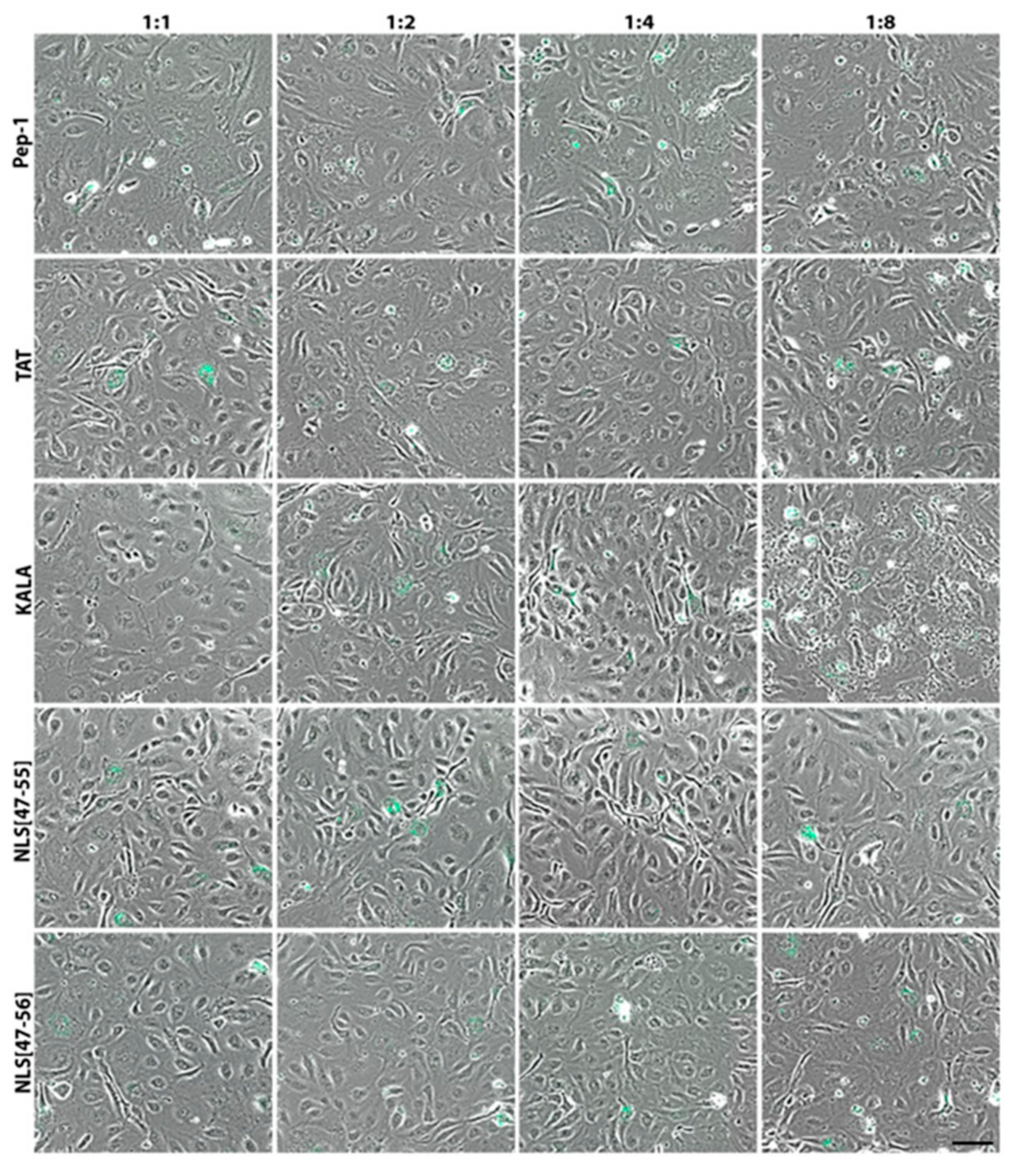
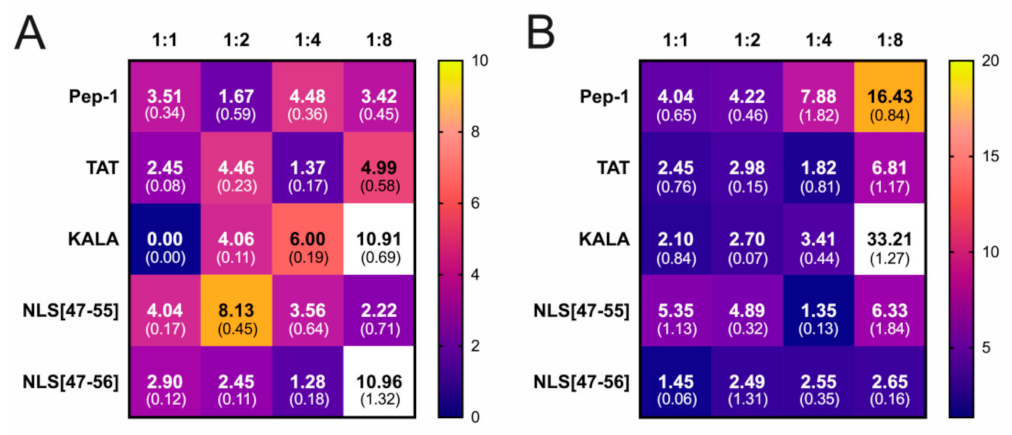
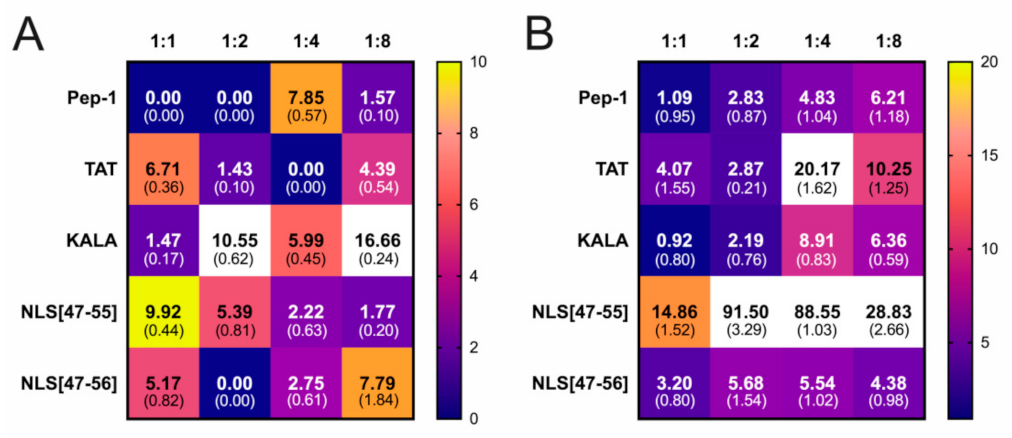
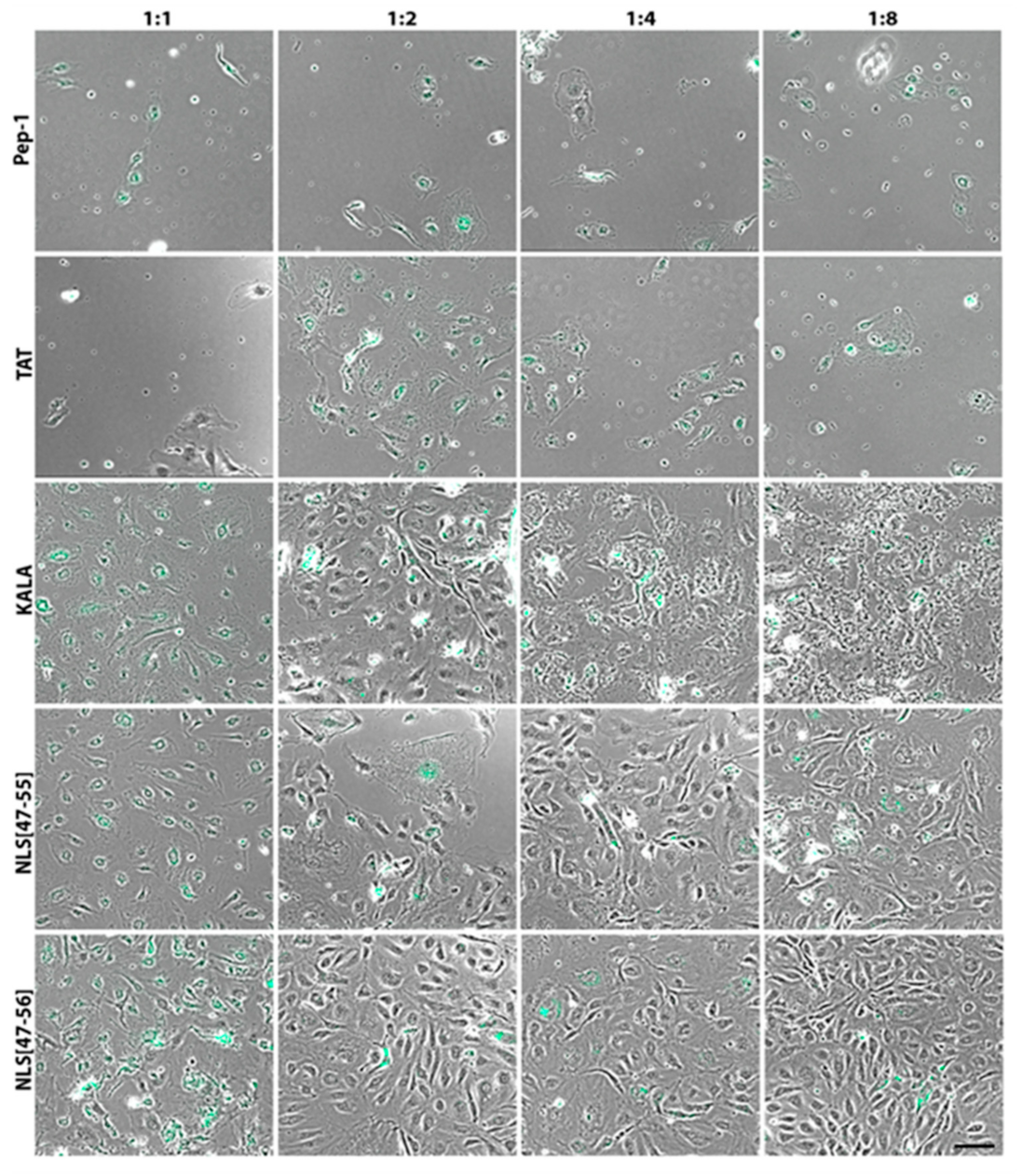
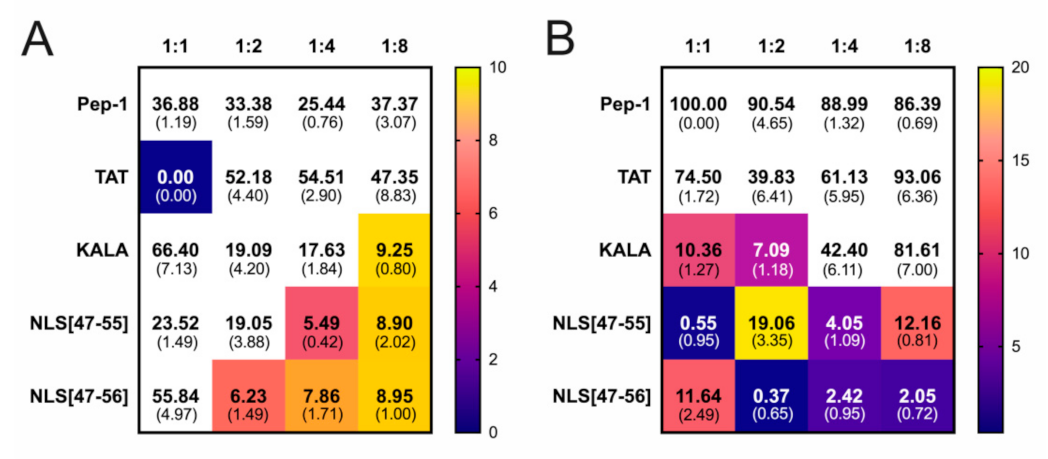
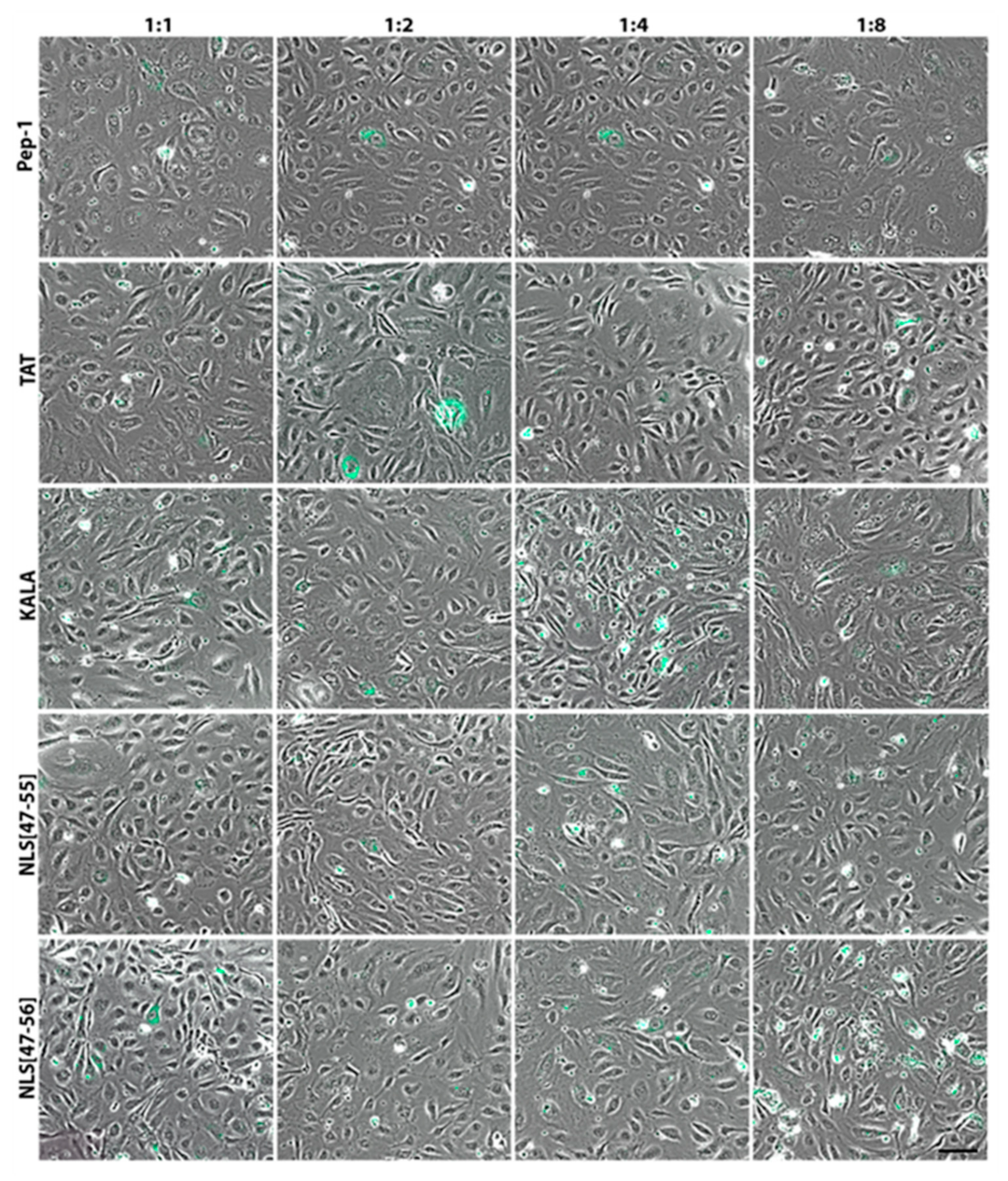
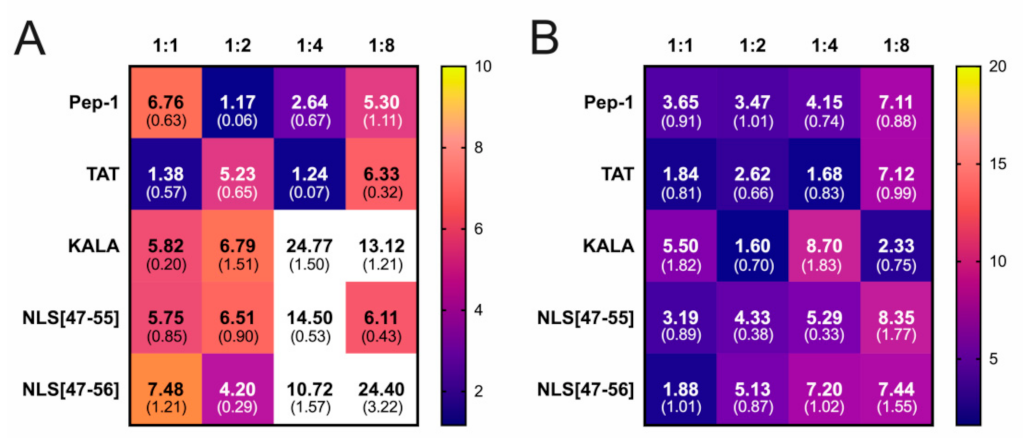
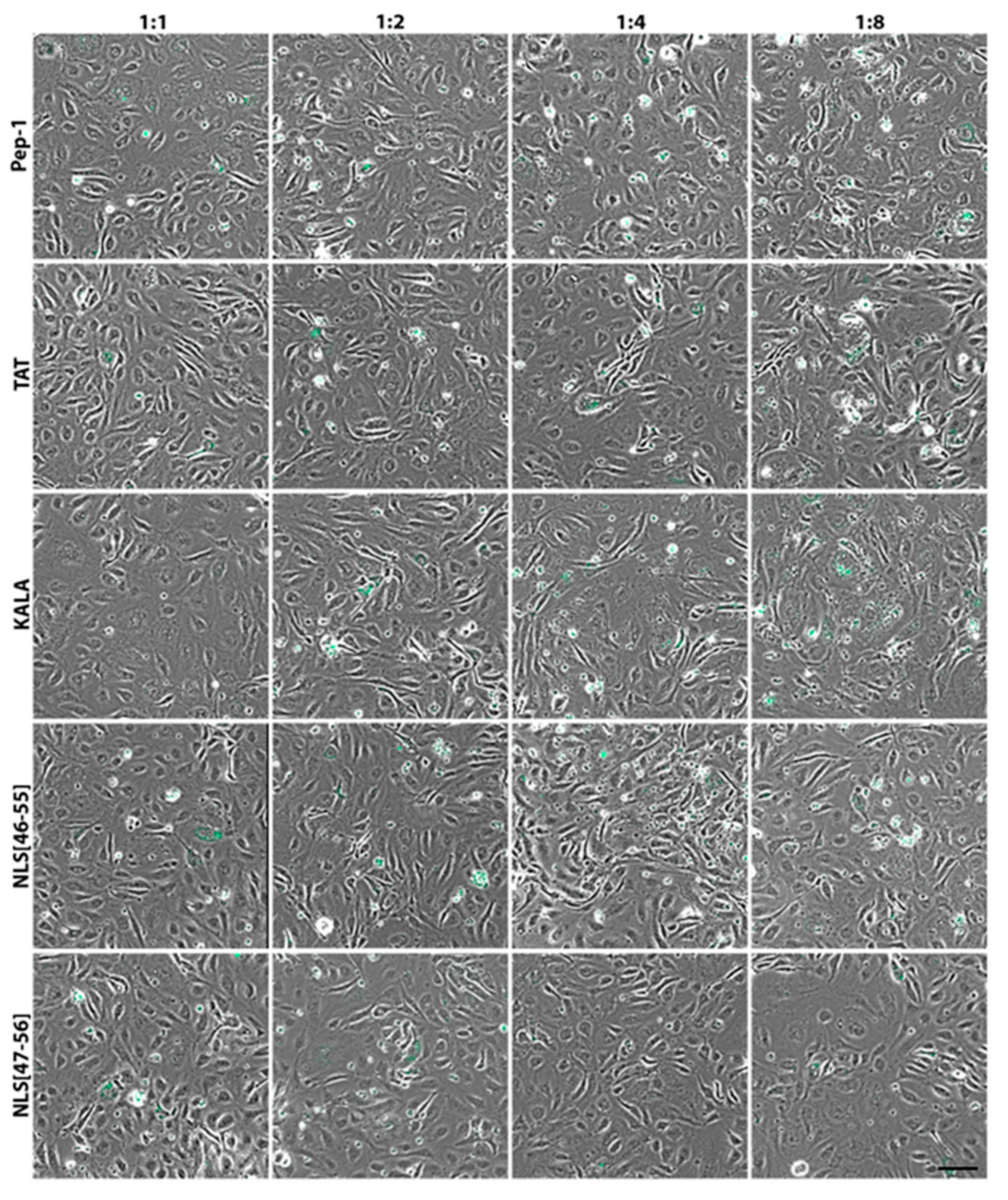
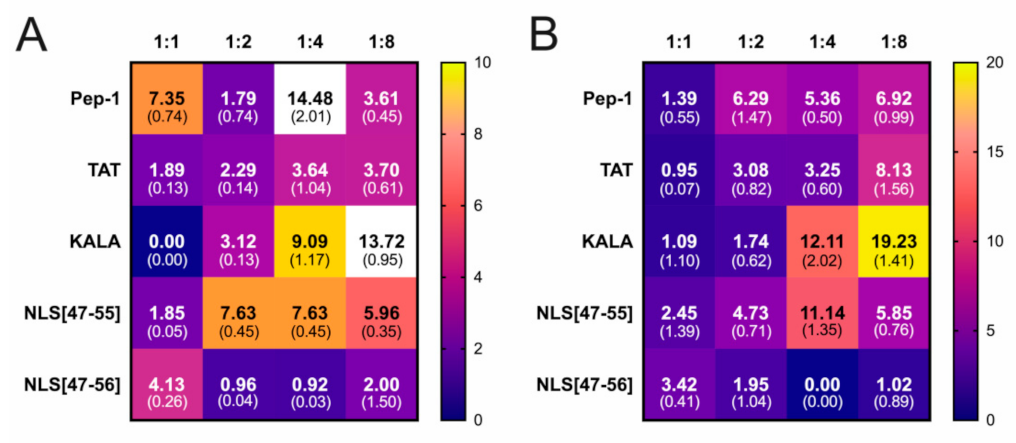
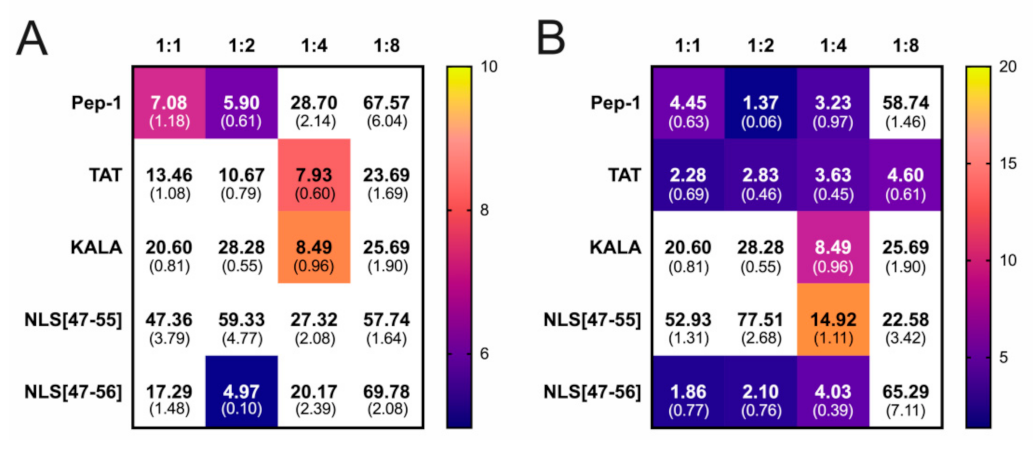
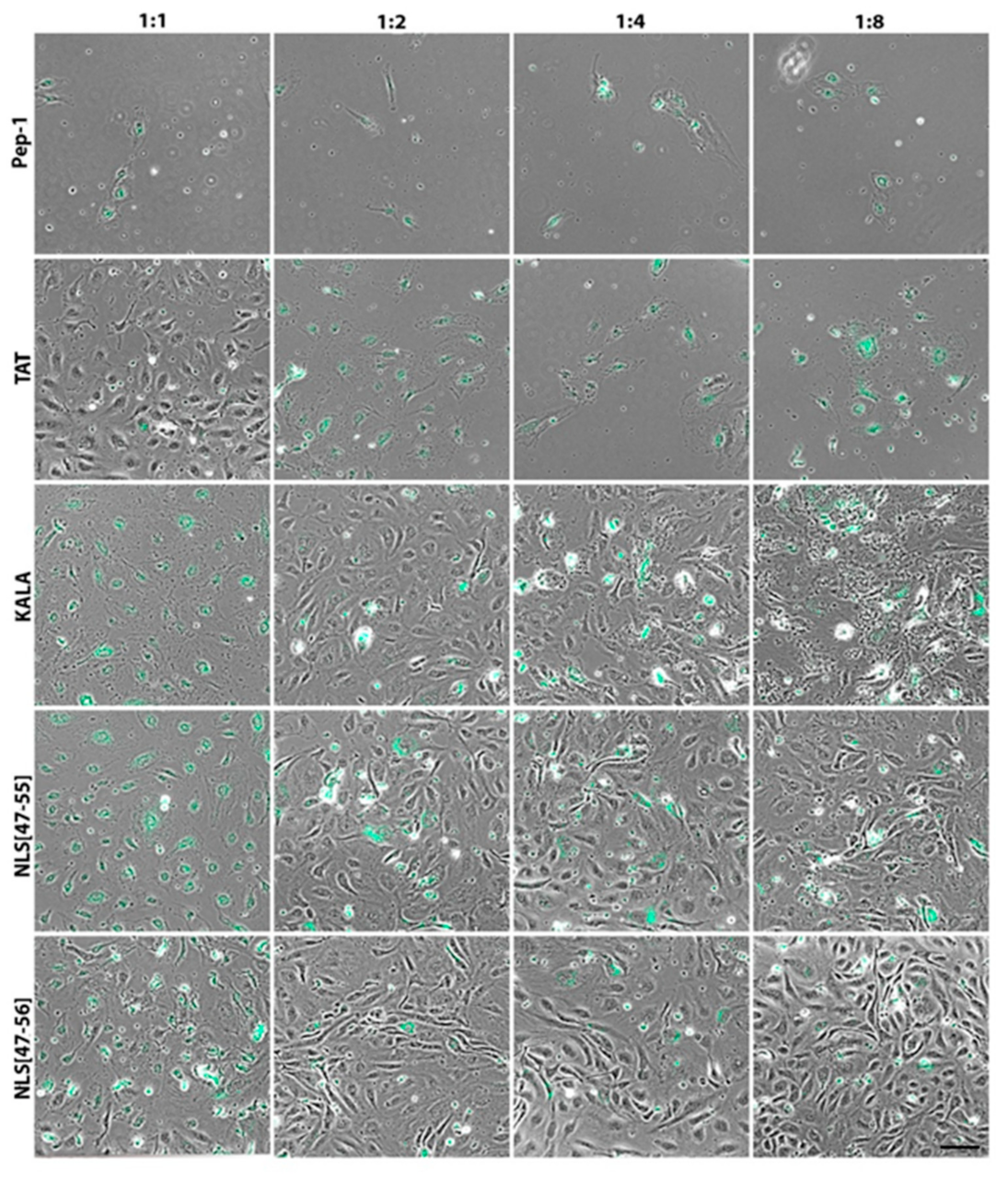
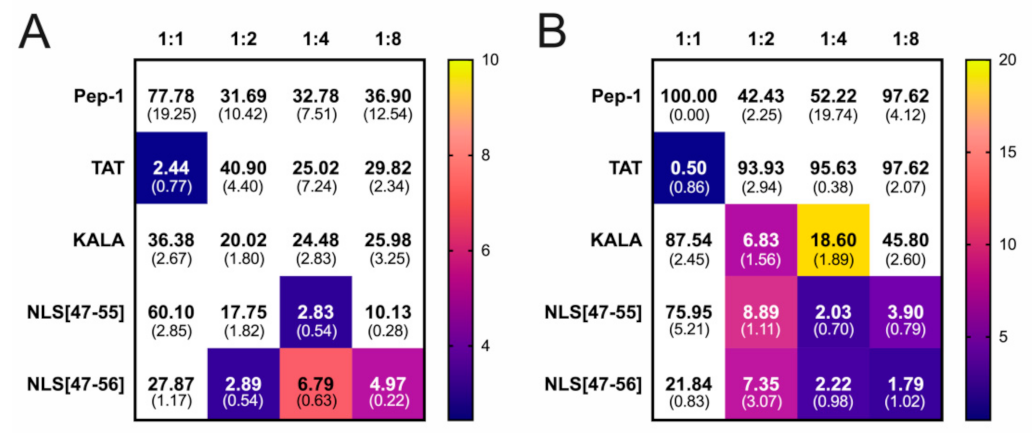
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gagat, M.; Zielińska, W.; Grzanka, A. Cell-penetrating peptides and their utility in genome function modifications (Review). Int. J. Mol. Med. 2017, 40, 1615–1623. [Google Scholar] [CrossRef] [Green Version]
- Meade, B.R.; Dowdy, S.F. Exogenous siRNA delivery using peptide transduction domains/cell penetrating peptides. Adv. Drug Deliv. Rev. 2007, 59, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chen, Y.; Liu, E.; Gong, J.; Shin, M.C.; Huang, Y. CPP-mediated protein delivery in a noncovalent form: Proof-of-concept for percutaneous and intranasal delivery. Protein Pept. Lett. 2014, 21, 1129–1136. [Google Scholar] [CrossRef] [PubMed]
- Iwase, Y.; Kamei, N.; Khafagy, E.S.; Miyamoto, M.; Takeda-Morishita, M. Use of a non-covalent cell-penetrating peptide strategy to enhance the nasal delivery of interferon beta and its PEGylated form. Int. J. Pharm. 2016, 510, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Pärnaste, L.; Arukuusk, P.; Langel, K.; Tenson, T.; Langel, Ü. The Formation of Nanoparticles between Small Interfering RNA and Amphipathic Cell-Penetrating Peptides. Mol. Ther. Nucleic Acids 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Afshar, S. In Vitro Assays: Friends or Foes of Cell-Penetrating Peptides. Int. J. Mol. Sci. 2020, 21, 4719. [Google Scholar] [CrossRef]
- Bucci, M.; Gratton, J.P.; Rudic, R.D.; Acevedo, L.; Roviezzo, F.; Cirino, G.; Sessa, W.C. In vivo delivery of the caveolin-1 scaffolding domain inhibits nitric oxide synthesis and reduces inflammation. Nat. Med. 2000, 6, 1362–1367. [Google Scholar] [CrossRef] [PubMed]
- US National Library of Medicine. ClinicalTrials.gov. 2019. Available online: https://clinicaltrials.gov/ct2/show/NCT00914914 (accessed on 12 January 2021).
- Tripathi, P.P.; Arami, H.; Banga, I.; Gupta, J.; Gandhi, S. Cell penetrating peptides in preclinical and clinical cancer diagnosis and therapy. Oncotarget 2018, 9, 37252–37267. [Google Scholar] [CrossRef] [Green Version]
- Incalza, M.A.; D’Oria, R.; Natalicchio, A.; Perrini, S.; Laviola, L.; Giorgino, F. Oxidative stress and reactive oxygen species in endothelial dysfunction associated with cardiovascular and metabolic diseases. Vascul. Pharmacol. 2018, 100, 1–19. [Google Scholar] [CrossRef]
- Widmer, R.J.; Allison, T.G.; Keane, B.; Dallas, A.; Bailey, K.R.; Lerman, L.O.; Lerman, A. Workplace Digital Health Is Associated with Improved Cardiovascular Risk Factors in a Frequency-Dependent Fashion: A Large Prospective Observational Cohort Study. PLoS ONE 2016, 11, e0152657. [Google Scholar] [CrossRef] [Green Version]
- Poh, K.K.; Lu, P.; Qin, G.; Silver, M.; Losordo, D.W.; Mendelsohn, M.E.; Zhu, Y. Endothelial dysfunction and systemic hypertension by selective cGMP-dependent protein kinase I inhibition using novel cell-penetrating peptide delivered in vivo. Int. J. Cardiol. 2013, 167, 2114–2119. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Wang, J.N.; Huang, Y.Z.; Guo, L.Y. Expression and purification of PEP-1-EGFP fusion protein and its transduction into human umbilical vein endothelial cells. Nan Fang Yi Ke Da Xue Xue Bao 2006, 26, 1114–1117. [Google Scholar] [PubMed]
- US National Library of Medicine. ClinicalTrials.gov. 2019. Available online: https://www.clinicaltrials.gov/ct2/show/NCT00451256 (accessed on 12 January 2021).
- Guidotti, G.; Brambilla, L.; Rossi, D. Cell-Penetrating Peptides: From Basic Research to Clinics. Trends Pharmacol. Sci. 2017, 38, 406–424. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Yan, Y.; Ni, D.; Wu, S.; Chen, Y.; Chen, X.; Xiong, X.; Liu, G. TAT-functionalized PEI-grafting rice bran polysaccharides for safe and efficient gene delivery. Int. J. Biol. Macromol. 2020, 146, 1076–1086. [Google Scholar] [CrossRef] [PubMed]
- Miura, N.; Shaheen, S.M.; Akita, H.; Nakamura, T.; Harashima, H. A KALA-modified lipid nanoparticle containing CpG-free plasmid DNA as a potential DNA vaccine carrier for antigen presentation and as an immune-stimulative adjuvant. Nucleic Acids Res. 2015, 43, 1317–1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, A.A.; Mussbach, F.; Breitling, R.; Hemmerich, P.; Schaefer, B.; Lorkowski, S.; Reissmann, S. Relationships between Cargo, Cell Penetrating Peptides and Cell Type for Uptake of Non-Covalent Complexes into Live Cells. Pharmaceuticals 2013, 6, 184–203. [Google Scholar] [CrossRef] [Green Version]
- Gros, E.; Deshayes, S.; Morris, M.C.; Aldrian-Herrada, G.; Depollier, J.; Heitz, F.; Divita, G. A non-covalent peptide-based strategy for protein and peptide nucleic acid transduction. Biochim. Biophys. Acta 2006, 1758, 384–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mo, R.H.; Zaro, J.L.; Shen, W.C. Comparison of cationic and amphipathic cell penetrating peptides for siRNA delivery and efficacy. Mol. Pharm. 2012, 9, 299–309. [Google Scholar] [CrossRef]
- Morris, M.C.; Vidal, P.; Chaloin, L.; Heitz, F.; Divita, G. A new peptide vector for efficient delivery of oligonucleotides into mammalian cells. Nucleic Acids Res. 1997, 25, 2730–2736. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, C.; Plank, C.; Lausier, J.; Schillinger, U.; Müller, R.H.; Rosenecker, J. Oligomers of the arginine-rich motif of the HIV-1 TAT protein are capable of transferring plasmid DNA into cells. J. Biol. Chem. 2003, 278, 11411–11418. [Google Scholar] [CrossRef] [Green Version]
- Ramakrishna, S.; Kwaku Dad, A.B.; Beloor, J.; Gopalappa, R.; Lee, S.K.; Kim, H. Gene disruption by cell-penetrating peptide-mediated delivery of Cas9 protein and guide RNA. Genome Res. 2014, 24, 1020–1027. [Google Scholar] [CrossRef] [Green Version]
- Wan, Y.; Moyle, P.M.; Christie, M.P.; Toth, I. Nanosized, peptide-based multicomponent DNA delivery systems: Optimization of endosome escape activity. Nanomedicine 2016, 11, 907–919. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Huang, J.; Holden, M.A.; Chen, M. Peptide-Mediated Membrane Transport of Macromolecular Cargo Driven by Membrane Asymmetry. Anal. Chem. 2017, 89, 12369–12374. [Google Scholar] [CrossRef] [PubMed]
- Ben-Dov, N.; Korenstein, R. The uptake of HIV Tat peptide proceeds via two pathways which differ from macropinocytosis. Biochim. Biophys. Acta 2015, 1848, 869–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurzawa, L.; Pellerano, M.; Morris, M.C. PEP and CADY-mediated delivery of fluorescent peptides and proteins into living cells. Biochim. Biophys. Acta 2010, 1798, 2274–2285. [Google Scholar] [CrossRef]
- Upadhya, A.; Sangave, P.C. Hydrophobic and electrostatic interactions between cell penetrating peptides and plasmid DNA are important for stable non-covalent complexation and intracellular delivery. J. Pept. Sci. 2016, 22, 647–659. [Google Scholar] [CrossRef]
- Koo, J.H.; Yoon, H.; Kim, W.J.; Lim, S.; Park, H.J.; Choi, J.M. Cell membrane penetrating function of the nuclear localization sequence in human cytokine IL-1α. Mol. Biol. Rep. 2014, 41, 8117–8126. [Google Scholar] [CrossRef] [PubMed]























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of CPP/NLS | CR 1 | Amount of Plasmid [μg] | Time of Incubation [min] | Time of Culture [h] | Temperature of Incubation [°C] | ||||
|---|---|---|---|---|---|---|---|---|---|
| TAT | 1:1 | 1 | 2 | 15 | 30 | 24 | 48 | 21 | 37 |
| 1:2 | 2 | 15 | 30 | 24 | 48 | 21 | 21 | ||
| 1:4 | 1 | 2 | 15 | 24 | 24 | 48 | 21 | 37 | |
| 1:8 | 1 | 2 | 15 | 30 | 24 | 48 | 21 | 37 | |
| 1:16 | 1 | 15 | 30 | 24 | 48 | 21 | 37 | ||
| Pep-1 | 1:1 | 1 | 2 | 15 | 30 | 24 | 48 | 21 | 37 |
| 1:2 | 2 | 15 | 30 | 24 | 48 | 21 | 37 | ||
| 1:4 | 1 | 2 | 15 | 30 | 24 | 48 | 21 | 21 | |
| 1:8 | 1 | 2 | 15 | 30 | 24 | 48 | 21 | 37 | |
| 1:16 | 1 | 15 | 30 | 24 | 48 | 21 | 37 | ||
| KALA | 1:1 | 1 | 2 | 15 | 30 | 24 | 48 | 21 | 21 |
| 1:2 | 2 | 15 | 30 | 24 | 48 | 21 | 37 | ||
| 1:4 | 1 | 2 | 15 | 30 | 24 | 48 | 21 | 37 | |
| 1:8 | 1 | 2 | 15 | 30 | 24 | 48 | 21 | 21 | |
| 1:16 | 1 | 15 | 30 | 24 | 48 | 21 | 37 | ||
| NLS [47–55] | 1:1 | 2 | 15 | 30 | 24 | 48 | 21 | 37 | |
| 1:2 | 2 | 15 | 30 | 24 | 48 | 21 | 21 | ||
| 1:4 | 2 | 15 | 30 | 24 | 48 | 21 | 37 | ||
| 1:8 | 2 | 15 | 30 | 24 | 48 | 21 | 37 | ||
| NLS [47–56] | 1:1 | 2 | 15 | 30 | 24 | 48 | 21 | 21 | |
| 1:2 | 2 | 15 | 30 | 24 | 48 | 21 | 37 | ||
| 1:4 | 2 | 15 | 30 | 24 | 48 | 21 | 37 | ||
| 1:8 | 2 | 15 | 30 | 24 | 48 | 21 | 37 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zielińska, W.; Gagat, M.; Mikołajczyk, K.; Hałas-Wiśniewska, M.; Grzanka, A. Low Effectiveness of the Introduction of pmaxGFP into Primary Human Coronary Endothelial Cells Using Cell-Penetrating Peptides and Nuclear-Localization Sequences in Non-Covalent Interactions. Appl. Sci. 2021, 11, 1997. https://0-doi-org.brum.beds.ac.uk/10.3390/app11051997
Zielińska W, Gagat M, Mikołajczyk K, Hałas-Wiśniewska M, Grzanka A. Low Effectiveness of the Introduction of pmaxGFP into Primary Human Coronary Endothelial Cells Using Cell-Penetrating Peptides and Nuclear-Localization Sequences in Non-Covalent Interactions. Applied Sciences. 2021; 11(5):1997. https://0-doi-org.brum.beds.ac.uk/10.3390/app11051997
Chicago/Turabian StyleZielińska, Wioletta, Maciej Gagat, Klaudia Mikołajczyk, Marta Hałas-Wiśniewska, and Alina Grzanka. 2021. "Low Effectiveness of the Introduction of pmaxGFP into Primary Human Coronary Endothelial Cells Using Cell-Penetrating Peptides and Nuclear-Localization Sequences in Non-Covalent Interactions" Applied Sciences 11, no. 5: 1997. https://0-doi-org.brum.beds.ac.uk/10.3390/app11051997