
Effect of Allium senescens Extract on Sorafenib Resistance in Hepatocarcinoma Cells


1
Department of Clinical Laboratory Sciences, College of Health Science, Korea University, Seoul 02841, Korea
2
Life Together, 13 Gongdan-ro, Chuncheon-si 24232, Korea
3
Mitosbio, 13, Gongdan-ro, Chuncheon-si 24232, Korea
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Appl. Sci. 2021, 11(8), 3696; https://0-doi-org.brum.beds.ac.uk/10.3390/app11083696
Submission received: 17 March 2021
/
Revised: 14 April 2021
/
Accepted: 16 April 2021
/
Published: 20 April 2021
(This article belongs to the Special Issue Functional Foods and Natural Products: Bioactive Compounds and Beneficial Effects on Health)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Although Allium species are involved in bioactivity, to the best of our knowledge, there is no research on the effects of Allium senescens on drug resistance in hepatocarcinoma. Ultra-high performance liquid chromatography was used to determine the concentration of several bioactive compounds in A. senescens extract; flow cytometry, reverse transcription–quantitative polymerase chain reaction, and siRNA-mediated knockdown to estimate the levels of different markers in HepG2 cells. The quantity of p-coumaric acid in the extract was 4.7291 ± 0.06 μg/mL, and the protein of relevant evolutionary and lymphoid interest (PRELI) in the resistant cells decreased 2.1 times in the presence of p-coumaric acid. The resistant cells strongly downregulated the efflux transporters (ABCB1, ABCC2, and ABCG2) when exposed to the extract or p-coumaric acid and when PRELI was knocked down, in contrast to the influx proteins (OCT-1). Additionally, the extract induced mitochondrial apoptosis and suppressed autophagy. Consequently, the extract and p-coumaric acid attenuated drug resistance of HepG2 cells through the downregulation of PRELI, a key protein associated with the modulation of drug transporter expression, the activation of autophagy, and mitochondrial apoptosis. Our results indicate that A. senescens extract is beneficial in protecting cancer cells against drug resistance and sustaining the efficacy of sorafenib against liver cancer.
1. Introduction
Hepatocellular carcinoma (HCC) is a common liver cancer with approximately 75% of the cases occurring in Asia and Africa [1]. Although various therapies, including liver transplantation, surgical resection, radioembolization, radiofrequency ablation, and trans-arterial chemoembolization [2] have been developed to manage HCC, drug resistance occurs frequently with continuous chemotherapy [3,4] and combinatorial therapy [3,4].
Sorafenib, a multi-target kinase inhibitor, suppresses the proliferation and viability of cancer cells as well as inhibits angiogenesis in breast, skin, pancreatic, colon, and liver cancers [5,6]. It activates the production of reactive oxygen species (ROS), eventually leading to the activation of ROS-mediated apoptotic pathways in HCC cells [7]. Sorafenib exhibits its action by targeting several signaling pathways/molecules: mitogen-activated protein kinase (MAPK) signaling pathway, vascular endothelial growth factor receptors (VEGFRs), platelet-derived growth factor receptor (PDGFR), and autophagy activation through iron regulatory protein (IRE), mammalian target of rapamycin (mTOR), and Beclin [6]. Although sorafenib is an effective HCC treatment option, the survival time of HCC patients can be shortened because of possible drug resistance [5]. In such a case, resistant cells prevent cellular apoptosis through the activation of the phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt) pathway, autophagy, and epithelial–mesenchymal transition [8]. Moreover, resistant cells alter the tumor microenvironment by increasing the expression of hypoxia-inducible factor-1α (HIF-1α), also known as VEGF inducer, and multidrug resistance 1 (MDR1) gene activator [9].
The protein of relevant evolutionary and lymphoid interest (PRELI) plays an important role in cellular protection under several stress conditions, including chemotherapy, desiccation, and osmotic imbalance [10]. Additionally, PRELI contains tandem repeats of the late embryogenesis abundant (LEA) protein [10]. We previously reported that [11] group 3 LEA (G3LEA) induces drug resistance by modulating ATP-binding cassette (ABC) transporters and decreasing of ROS production in Enterococcus faecium.
Allium senescens, commonly known as German garlic from the lily family, is a perennial herb rich in flavonoids [12]. Reportedly, A. senescens extract attenuates carcinogenesis [13], modulates enzyme activities [14], and possesses antioxidant [15], antithrombotic [13], and antimutagenic [16] properties. Although many studies have been documented, to the best of our knowledge, there is none available on the effects of the A. senescens extract on drug resistance.
The present study documents the bioactive substances involved in modulating PRELI expression and the attenuation of drug resistance in HCC cells. Our findings suggest that A. senescens extract and p-coumaric acid can potentially act as functional biomaterials to suppress drug resistance in HCC cells when treated with sorafenib. Although we propose a further in vivo experiment and research on PRELI signaling, the current findings suggest that PRELI is a key protein to attenuate drug resistance and that the extract is an optimal substance for application as a biofunctional material for pharmaceuticals and as a functional food.
2. Materials and Methods
2.1. Cell Culture and Establishment of Treating Dosage
Human HCC cells (HepG2) (KCLB No. 88065, Seoul, Korea) were cultured in Dulbecco’s modified Eagle’s medium (DMEM) high glucose (Invitrogen, Carlsbad, MA, USA) with 10% fetus bovine serum (FBS) (Sigma, Darmstadt, Germany) at 37 °C and 5% CO2. After exposure to A. senescens extract, p-coumaric acid (Sigma), and sorafenib (Sigma) for 3 days, HepG2 cells were stained using the Annexin V-PI apoptosis detection kit (Invitrogen), and cellular viability was evaluated using a flow cytometer (BD FACScalibur, BD Biosciences, CA, USA) and FlowJo 10.6.1 (BD Biosciences) to establish a cytotoxic concentration (CC50) (Figure S2). In all experiments, the exposure time of all substances was 3 days. In all the samples, the concentrations of A. senescens extract, p-coumaric acid, and sorafenib used were 1 μg/L, 0.002 μmol/L, and 0.015 μmol/L, respectively.
2.2. Modeling of a Drug-Resistant Cell
HepG2 cells were cultured in DMEM with sorafenib at serial concentrations (0.005, 0.007, 0.010, and 0.015 μmol/L) for 70 days to model drug-resistant cells. After the exposure, the cellular viability was evaluated using Annexin V-PI (Invitrogen), a flow cytometer (BD FACScalibur, BD Biosciences), and FlowJo 10.6.1 (BD Biosciences).
2.3. Reverse Transcription–Quantitative Polymerase Chain Reaction (RT–qPCR)
Total RNA was isolated from the treated cells using RiboEx reagent (GeneAll, Seoul, Korea), and cDNA was synthesized from the isolated RNA using Maxime RT PreMix (iNtRON, Seongnam, Korea). qPCR was performed with primers (Table S1) at the following cycling parameters: 1 min at 95 °C, followed by 35 cycles of 35 s at 59 °C, and 1 min at 72 °C. The amplified DNA was estimated quantitatively using iBright FL1000 and iBright Analysis Software 4.0.0 (Invitrogen) (Figure S1).
2.4. Ultra-High Performance Liquid Chromatography (UHPLC)
The A. senescens extract was obtained from Life Together (Chuncheon, Korea) and Mitosbio (Chuncheon, Korea); compounds including p-coumaric acid (CAS Number: 501-98-4, Sigma, St. Louis, MO, USA), ferulic acid (CAS Number: 1135-24-6, Sigma, St. Louis, MO, USA), luteolin (CAS Number: 491-70-3, Sigma, St. Louis, MO, USA), naringenin (CAS Number: 67604-48-2, Sigma, St. Louis, MO, USA), and apigenin (CAS Number: 520-36-5, Sigma, St. Louis, MO, USA) in the extract were evaluated using UHPLC (Dionex Ultimate 3000 system, Thermo Fisher Scientific, Waltham, MA, USA) [17]. After air-drying and grinding (35 mesh), the leaves of A. senescens (Ulleung, Korea) were extracted twice with hydration for 90 min. The filtered extract was concentrated (R-3000, BuCHI Labortechnik AG, Germany) at 60 °C and lyophilized using a freeze dryer (FD8505, Ilshin Lab Co., Seoul, Korea).
To prepare the standards, p-coumaric acid was serially diluted (0.0625–1 mmol) with 100% dimethyl sulfoxide (DMSO). The extract (20 μL) and standards (20 μL) were injected into the column (YMC-triart C18, 250 × 4.6 mm I.D. S-5 μm, 12 nm, p/n TA12S05-2546WT). The flow conditions were as follows: Solvent A, 1% acetic acid; Solvent B, acetonitrile; flow rate, 0.7 mL/min; and gradient elution. The chromatograms were evaluated using a UV detector (Thermo Fisher Scientific) at three different wavelengths (272, 280, and 310 nm) with optimal absorbances for the standards.
2.5. Flow Cytometry
To estimate the levels of the markers, including PRELI, ATP-binding cassette C2 (ABCC2), organic cation transporter 1 (OCT-1), and organic anion transporting polypeptide 1B1 (OaTP1B1), the treated cells were fixed with 2% paraformaldehyde for 2 days and then treated with 0.02% Tween 20 for 10 min [18]. After blocking with an Fc blocking solution (BD Biosciences), the fixed cells were stained using FITC anti-PRELI (GeneTex, CA, USA), PE-anti-OCT-1 (Novusbio, CO, USA), APC-anti-OaTP1B1 (Thermo Fisher, MA, USA), and PerCP-anti-ABCC2 (Cell Signaling, MA, USA) for 3 days at 25 °C. Fluorescence labeling was performed using the Lightning-Link Rapid Antibody Labeling Kit (Fluor 488, 555, 647, 680) (Novusbio). Briefly, cells (5 × 107 cells/mL) were stained with fluorochromes and 104 cells in the samples were evaluated for their fluorescence intensity using a flow cytometer (FACScalibur, BD bioscience) and FlowJo 10.6.1 (BD bioscience). False fluorescent signals were eliminated using a negative control.
2.6. PRELI Knockdown
Briefly, the cultured HepG2 cells were exposed to Lipofectamine 2000 reagent (Invitrogen) with PRELI siRNA or control siRNA oligonucleotide (negative and positive) (Bioneer, Daejeon, Korea) for 24 h, and subsequently to sorafenib for 24 h. Data were compensated with the negative and positive controls. For PRELI overexpression, the cultured HepG2 cells were exposed to PRELI Lentiviral Activation Particles (h): sc-411975-LAC (Santa Cruz Biotechnology, TX, USA) with retronectin (Takara, Shiga, Japan).
2.7. Statistical Analysis
All experiments were performed in triplicates (n = 3), and the data were analyzed using one-way analysis of variance (ANOVA) with the post hoc test (Scheffe’s method) and an independent t-test using SPSS v. 26 (IBM, NY, USA) and Prism 7 (GraphPad, CA, USA) (p < 0.05 is significant).
3. Results
3.1. PRELI Expression in Resistant and Susceptible Cells
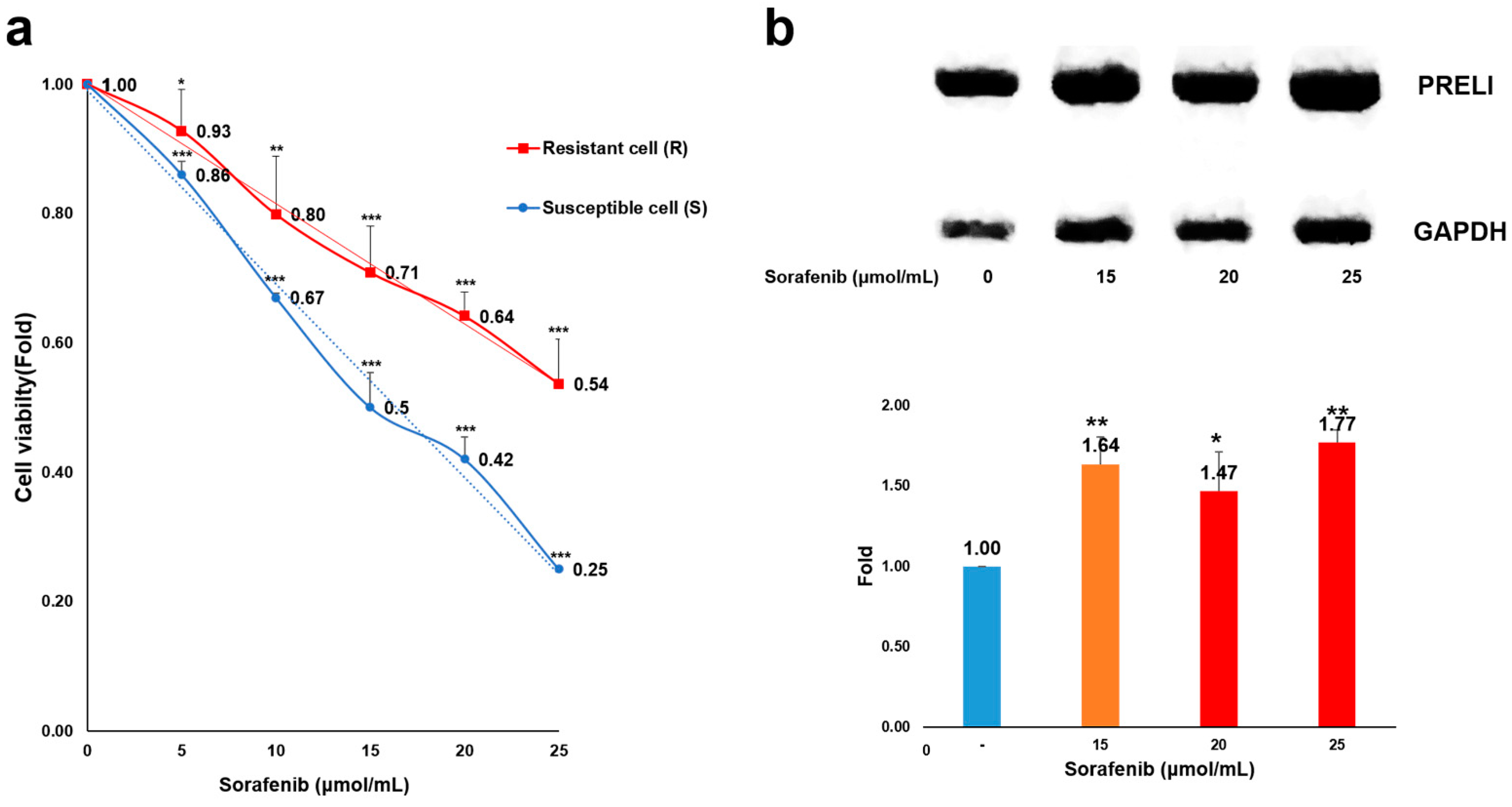
This study documented the biological functions of A. senescens extract and p-coumaric acid with respect to drug resistance in HCC cells. Based on the results of the modeling of drug-resistant cells, the resistant cells enhanced their viability against sorafenib with a proportional increase in PRELI level (Figure 1). At 15 µmol/mL, viability of the resistant cells increased by 42% compared to that of the susceptible cells. Additionally, compared with the viability of susceptible cells, the resistant cells were 2.15 times higher in number at 25 µmol/mL (Figure 1a,b).
3.2. Concentration of p-Coumaric Acid in the Extract
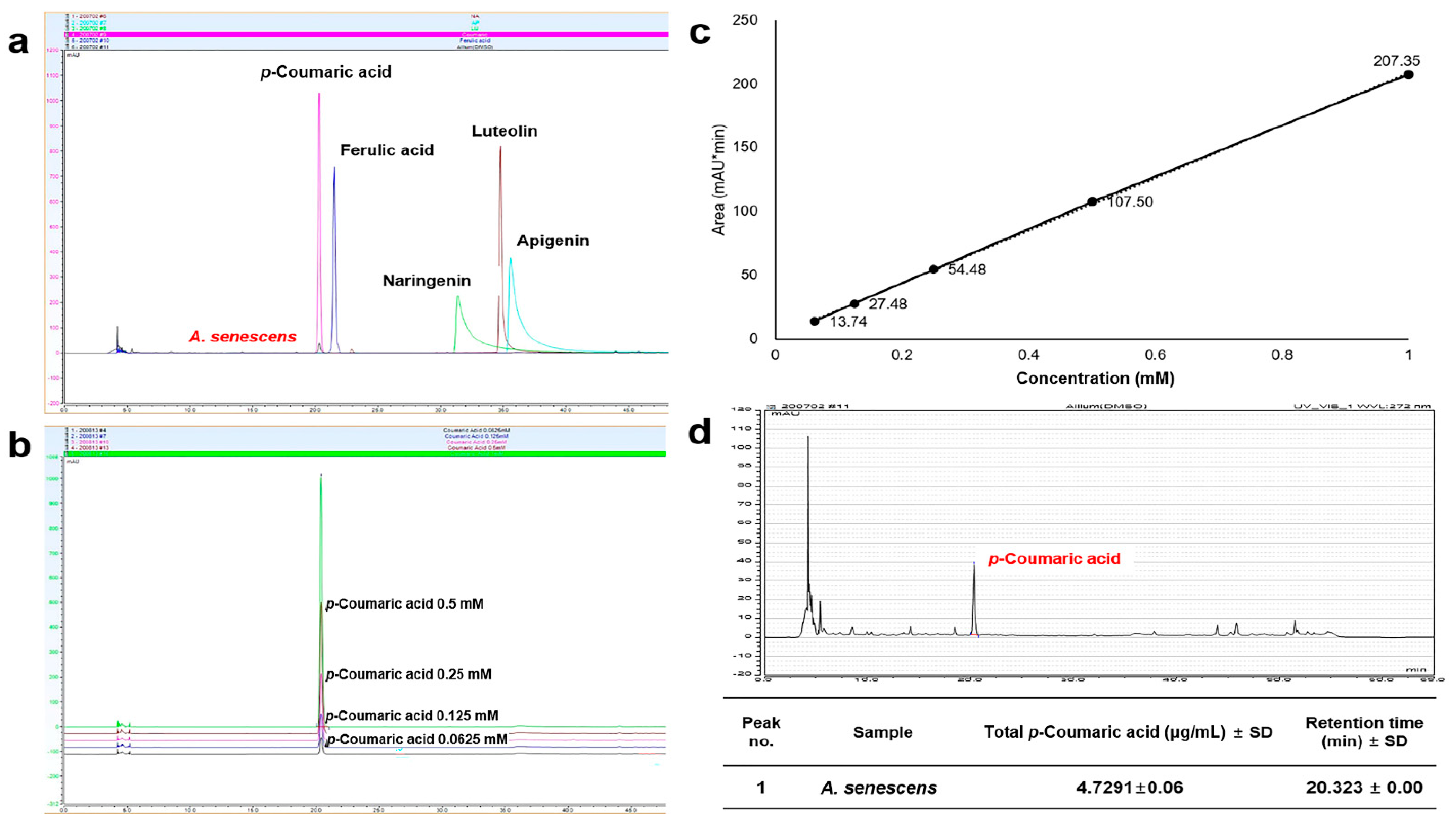
UHPLC showed that the extract contained p-coumaric acid at 4.7291 μg/mL, without the detection of other compounds such as ferulic acid, luteolin, naringenin, and apigenin (Figure 2).
3.3. Effects of p-Coumaric Acid on Drug Resistance
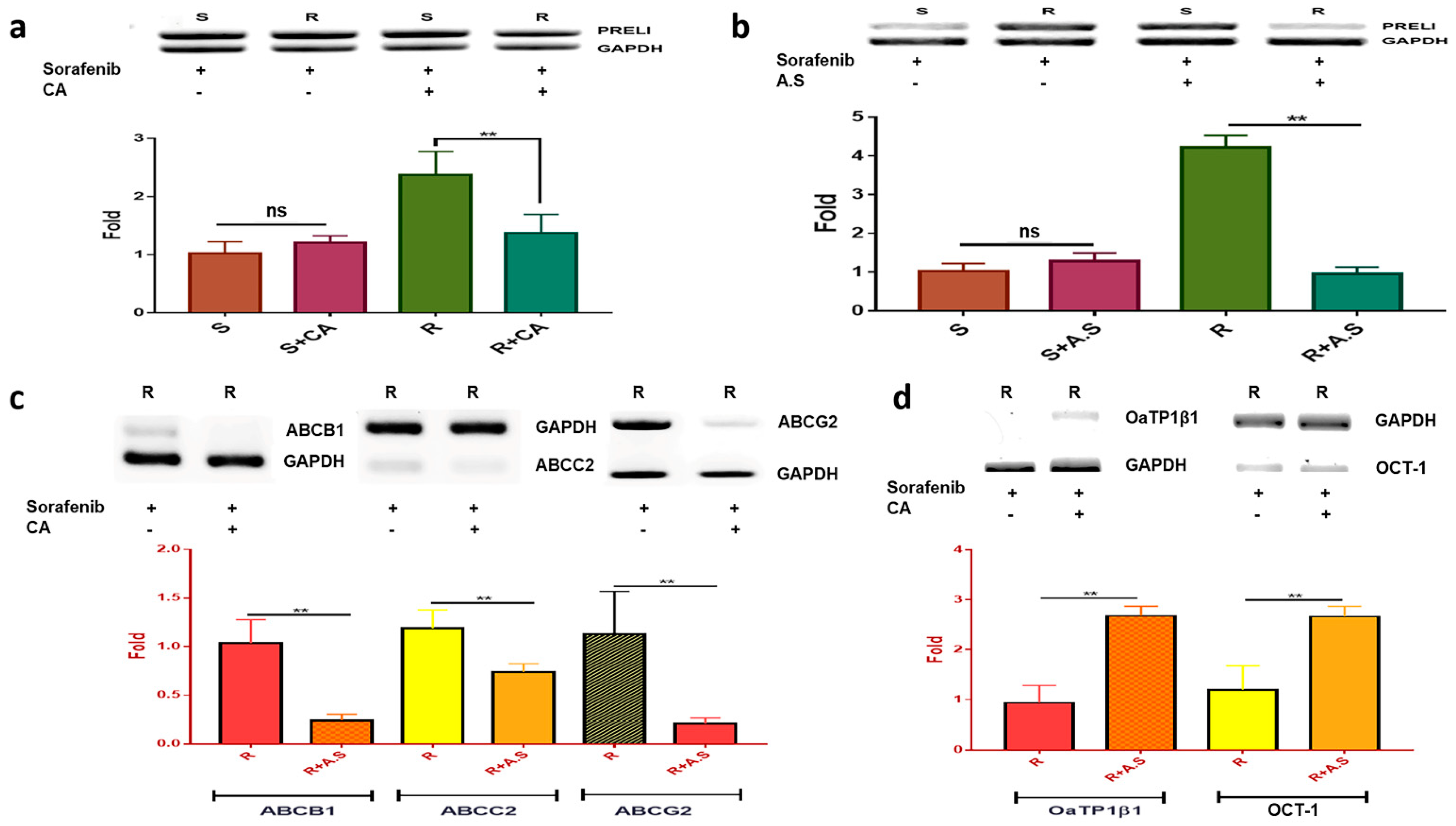
When exposed to p-coumaric acid, although the levels of PRELI were upregulated in the susceptible cells, the levels were downregulated in the resistant cells, similarly to the control (Figure 3a).
Although p-coumaric acid was not effective in considering changes in the PRELI levels in the susceptible cells, the PRELI level was downregulated approximately 2.1 times compared to that in the resistant cells without p-coumaric acid (Figure 3a). Similarity to p-coumaric acid, the extract downregulated PRELI level in the resistant cells, but it was more effective than p-coumaric acid (Figure 3b). p-coumaric acid attenuated the RNA levels of the efflux transporters ABCB1, ABCC2, and ABCG2 (Figure 3c). Contrary to the efflux transporters, the levels of the influx transporters OCT-1 and OaTP1β1 were upregulated in the resistant cells treated with p-coumaric acid (Figure 3d). Notably, the levels of ABCB1 and ABCG2 were 3.3–3.5 times downregulated compared to those in the resistant cells without p-coumaric acid (Figure 3c). The levels of OaTP1β1 were 2.6 times higher than those of the resistant cells when the cells were not exposed to p-coumaric acid (Figure 3d).
3.4. Roles of PRELI and the Extract in Drug Resistance
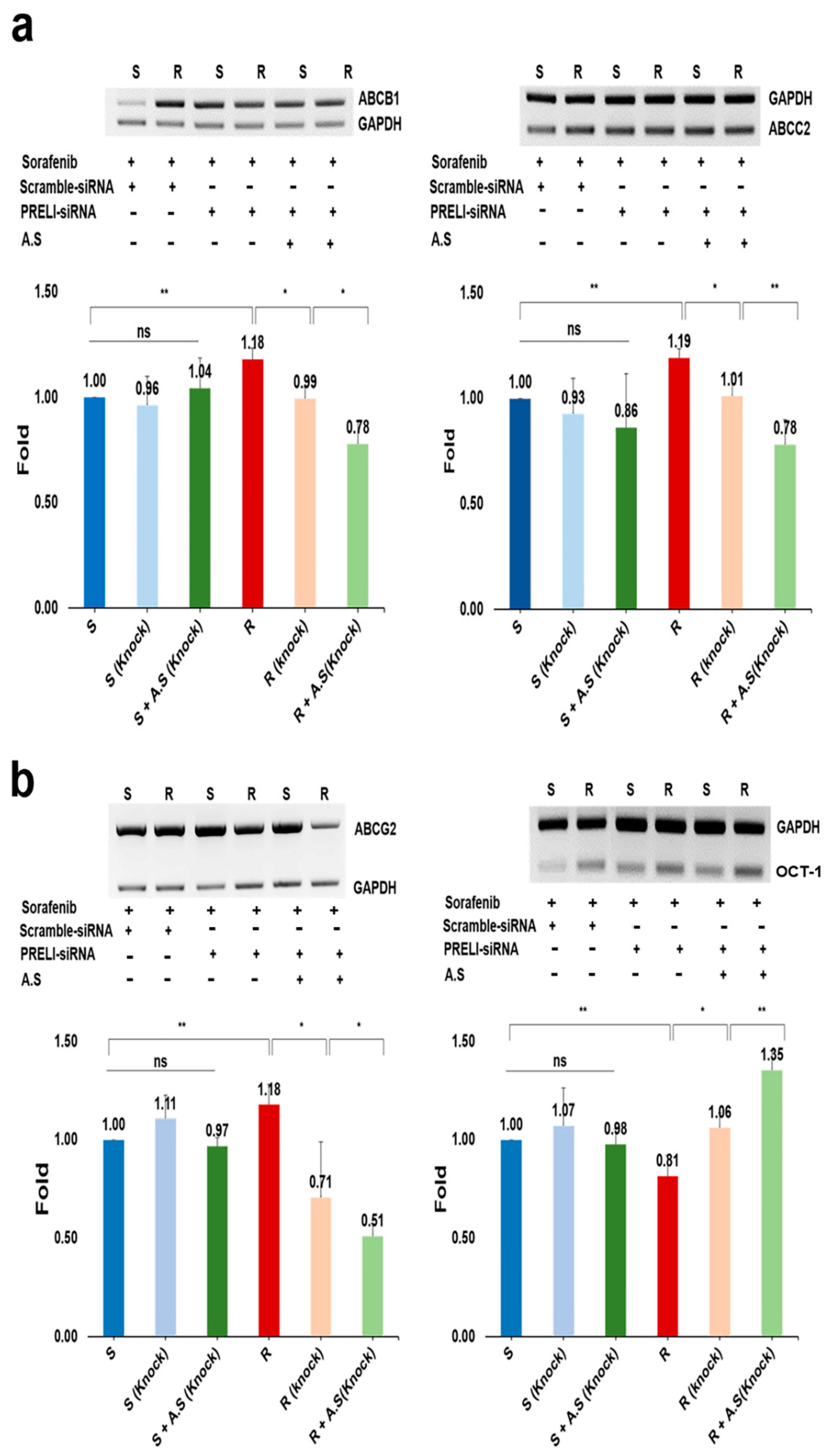
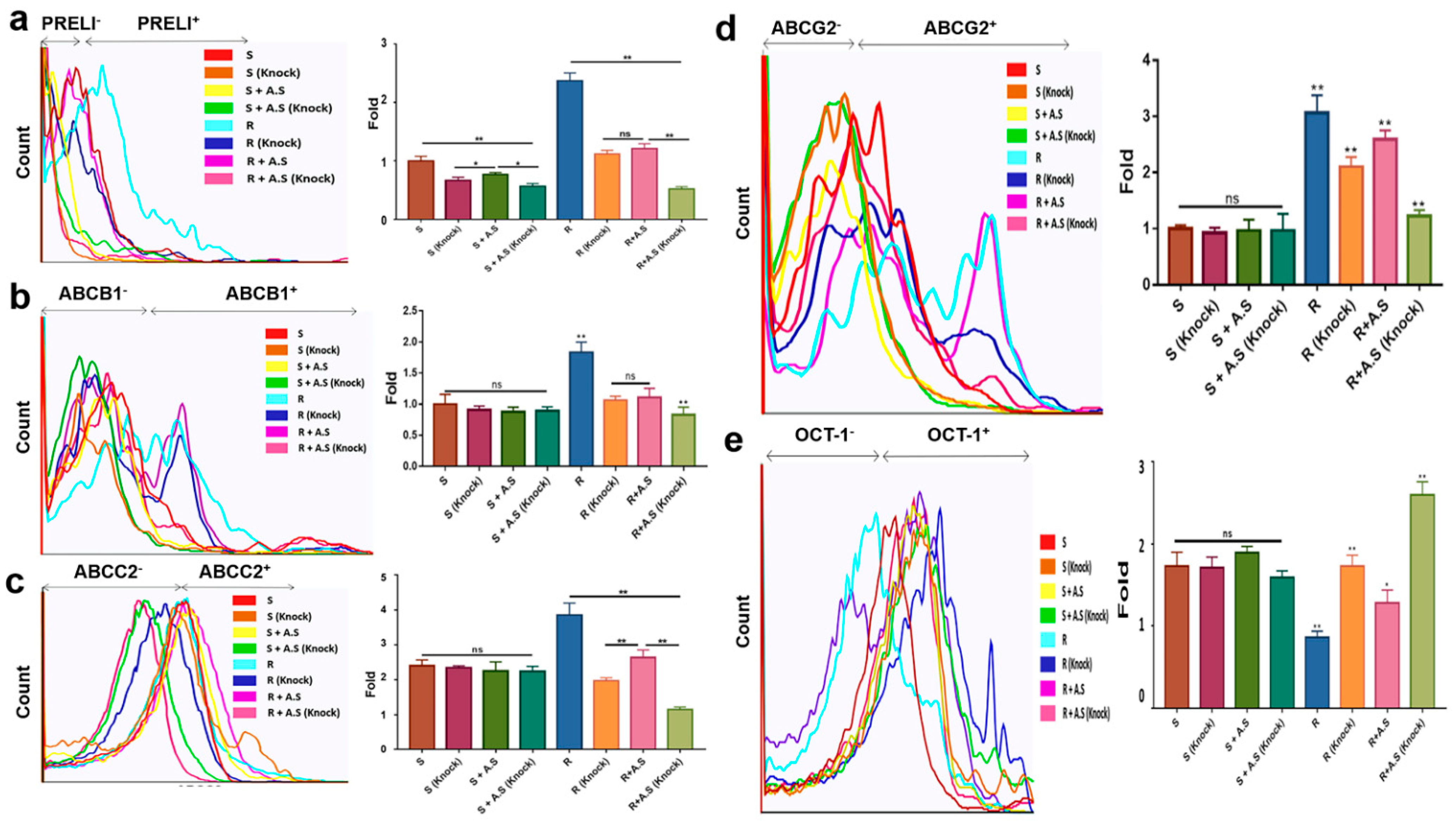
Although the levels of all transporters were not significant in susceptible cells, the transporters without OaTP1β1 were significant in resistant cells under the extract (Figure 4). Following PRELI knockdown (Figure 4a,b), the RNA levels of the efflux transporters were downregulated. Compared with other efflux transporters, alterations in ABCG2 levels were more sensitive to PRELI levels (Figure 4b). Synergistically with PRELI knockdown, the extract induced the downregulation of the RNA levels for efflux transporters in the resistant cells (Figure 4). Corresponding to the RNA levels, PRELI-knocked down resistant cells that showed an attenuated expression of the efflux transporters, and this attenuation was enhanced in the presence of the extract (Figure 5a–d and Table S2). When comparing the expression levels of OCT-1 in the susceptible cells, the resistant cells decreased the levels of the influx transporters by 2.3 times (Figure 5e and Table S2). Although the levels of OCT-1 following PRELI knockdown were not significant, a combination of extract-exposure and knockdown induced an increase in the levels (Figure 5e). Contrary to OCT-1, the levels of OaTP1β1 were not significant in susceptible and resistant cells (S3).
3.5. Role of the Extract in Apoptosis
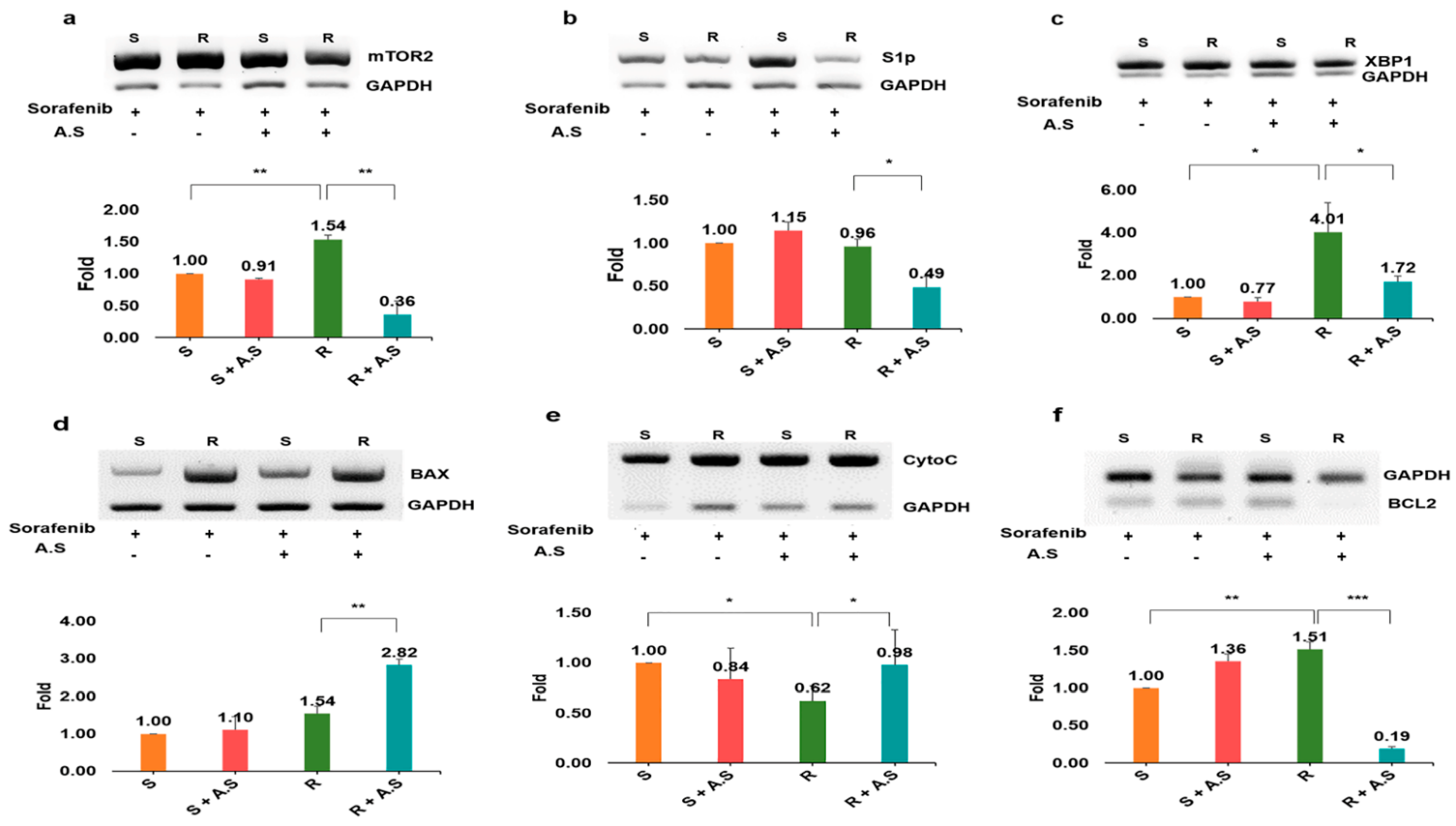
When evaluating the RNA levels of apoptotic markers (Figure 6), unlike sorafenib, the extract dramatically altered the levels of the markers in the resistant cells. Interestingly, although the extract was not effective against susceptible cells, the resistant cells showed attenuated drug resistance through the modulation of autophagy and mitochondrial apoptosis markers when exposed to the extract (Figure 6). Among the autophagy markers mTOR2, sphingosine-1-phosphate (S1P), and x-box-binding protein (XBP1), the level of mTOR2 was dramatically decreased, that is, approximately 9.5 times lower than that of the resistant cells exposed to the extract (Figure 6a–c). Furthermore, contrary to bcl-2-associated X protein (BAX) and cytochrome p450 C (CytoC), the extract suppressed B-cell lymphoma 2 (BCL2) levels in resistant cells (Figure 6d–f).
3.6. Localization of PRELI in HCC cells
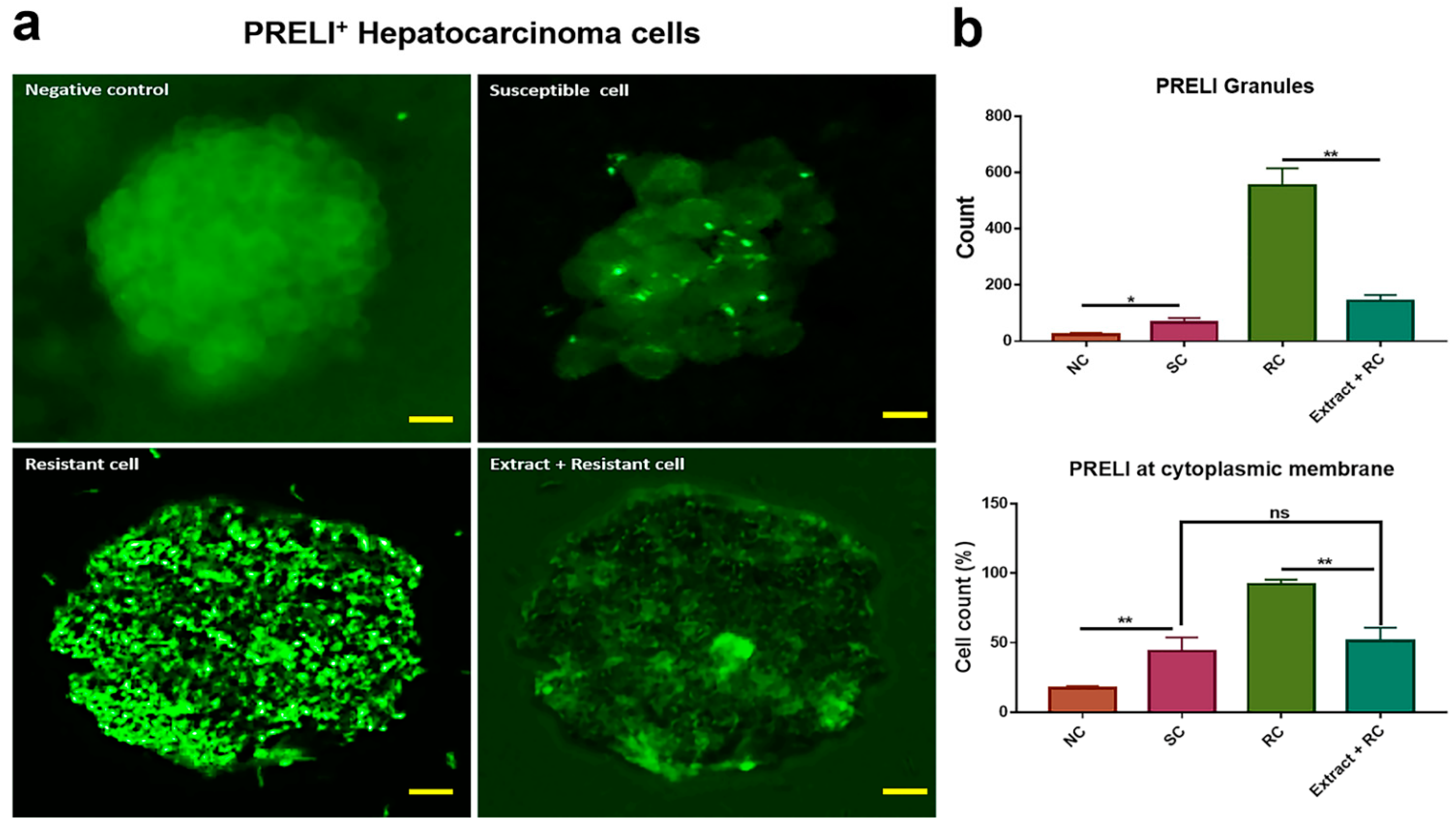
As indicated by Figure 7, although PRELI was stained densely near the cytoplasmic membrane in resistant cells, it was found frequently in the cytoplasm of susceptible cells. Compared with susceptible cells, the number of granules in resistant cells was 10.2 times higher. Furthermore, the counts of the stained cells at the cytoplasmic membrane were 2.1 times higher than those in the susceptible cells. The resistant cells showed results similar to those of the susceptible cells when exposed to the extract.
4. Discussion
HCC, a primary liver cancer, is the second most fatal cancer type worldwide. Obesity is a major causative reason underlying HCC, wherein the modulation of obesity is important to reduce its incidence rate [19]. Notably, drug resistance occurs frequently and adversely affects the efficient HCC treatment. The attenuation of drug resistance is crucial for the effective treatment of HCC and finding a natural bioactive substance that can reduce drug resistance is indeed essential.
Drug-resistant cells express high levels of PRELI, in addition to high cellular viability. In this study (Figure 1), we demonstrated the modeling of resistant cells. Allium species are effective in the treatment of various diseases through their bioactive compounds, including sulfur compounds and polyphenols [20,21]. Notably, A. senescens is involved in suppressing cancer growth and development, it has anti-inflammatory and antioxidative activities, and contains bioactive compounds, such as allicin, alliin, antifungal activators [20,22], and polyphenols (hydroxycinnamic acids such as ferulic acid, p-coumaric acid, and phytosterols) [20]. High-end chromatography revealed that the A. senescens extract contained p-coumaric acid, with the exception of ferulic acid, luteolin, apigenin, and naringenin. As mentioned earlier, sorafenib enhances an increase in cellular ROS levels, and under the influence of sorafenib, polyphenols reinforce resistance to ROS to survive this stress. Unlike the resistant cells with upregulated PRELI levels, cells exposed to p-coumaric acid or the extract showed downregulated levels of PRELI. Notably, the extract strongly downregulated the expression of PRELI (Figure 3) in resistant cells. p-coumaric acid exhibits antioxidant, anti-proliferative, antibacterial, antiangiogenic, antimelanogenic, antimutagenic, and antigenotoxic properties as well as mediates lipoprotein peroxidation [23]. Additionally, although p-coumaric acid has been shown to be cytotoxic to drug-resistant leukemia cell lines [24], the cytotoxic mechanism was not demonstrated in drug-resistant cells. These results indicated that p-coumaric acid attenuates drug resistance through the downregulation of PRELI in resistant cells, wherein bioactive compounds associated with MDR reversing activators, including alkaloids, curcumin, dihydroptychantol A, guggulsterone, phenolic diterpenes, tetrandrine, and vincristine also induce such effects [25].
Contrary to the levels of the influx transporters, the expression of the efflux transporters was downregulated in the extract-exposed-resistant cells (Figure 4 and Figure 5). In addition, at low levels of PRELI, the resistant cells showed upregulated levels of the influx transporters (Figure 4 and Figure 5). Sorafenib is an important anticancer drug that affects HCC cells through OCT-1, and drug-resistant cells break down it to sorafenib glucuronide via cytochrome P450 [26]. The modified sorafenib, sorafenib glucuronide, is excreted by ABC transporters [27]. Resistant cells alter their genetic expression, including increases in ABCB1, ABCC2, and ABCG2, in contrast to OCT-1. In our previous report [11], we found that G3LEA containing the PRELI motif was associated with resistance against vancomycin in bacteria, and that the suppression of PRELI is an effective modulation of efflux and influx transporters in HCC cells. Attenuation of PRELI caused by the A. senescens extract and p-coumaric acid indicates that the two substances are effective in treating HCC without drug resistance. Furthermore, A. senescens was synergized with PRELI knockdown to regulate transporters in HCC cells. In particular, synergistic effects of the extract-exposed and PRELI-knocked down resistant cells are similar to those of the susceptible cells.
In addition to ordinary pathways, sorafenib enhances the apoptosis of HCC cells by altering various pathways, including the 5’ AMP-activated protein kinase (AMPK) pathway, microRNA regulation, sphingolipid metabolism, ER stress, and autophagy [28]. Although autophagic markers were downregulated in the resistant cells after treatment with the extract, mitochondrial apoptosis markers were upregulated in them (Figure 6). Interestingly, PRELI was localized near the cytoplasmic membrane in resistant cells, unlike in susceptible cells (Figure 7). Based on our previous study [11] and the current one, PRELI is involved in the drug resistance of prokaryotic cells, in addition to cellular proliferation, detoxification, and apoptosis. Moreover, many efflux and influx transporters are localized in the cytoplasmic membranes [27]. These results suggest that the extract-attenuated activity of HCC cells occurs through the downregulation of PRELI, which is a crucial protein for the survival of HCC cells under chemical stress.
This study suggests that the A. senescens extract can be applied in further in vivo experiments to elucidate attenuation of drug resistance in HCC. Additionally, the results for PRELI provide directions for further research associated with the signaling pathways and effects of PRELI under stress conditions in HCC cells.
5. Conclusions
This study documents the functions of A. senescens extract with respect to drug-resistant HCC cells. First, the extract enhanced the downregulation of PRELI levels, which acts as a key protein in drug resistance. Second, p-coumaric acid, a compound in the extract, also attenuated the PRELI expression in the resistant cells. Third, the downregulation of PRELI by the extract improved drug resistance in resistant cells by inhibiting autophagy and activating mitochondrial apoptosis. Consequently, A. senescens extract is effective against drug resistance, maintaining the efficacy of sorafenib against HCC. Additionally, microRNA profiling in the induced exosomes of cells exposed to the extract or with knocked down PRELI will provide crucial insights into the development of drugs or biomaterials as functional foods for the prevention and treatment of HCC.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/app11083696/s1, Figure S1: Full gels, and Table S1: Primers. Figure S2: Viability and levels of transporter in HepG2 cells. Table S2: Records from flow cytometer for Figure 5.
Author Contributions
Conceptualization and methodology: S.P., Y.P., and H.S.; writing—original draft preparation: Y.P. and B.K.; writing—review and editing: B.K.; supervision: B.K. and S.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
This study was supported by CO., Ltd., Life Together, and Korea University.
Conflicts of Interest
The authors declare no conflict of interest.
References
- McGlynn, K.A.; Petrick, J.L.; London, W.T. Global epidemiology of hepatocellular carcinoma: An emphasis on demographic and regional variability. Clin. Liver Dis. 2015, 19, 223–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raza, A.; Sood, G.K. Hepatocellular carcinoma review: Current treatment, and evidence-based medicine. World J. Gastroenterol. WJG 2014, 20, 4115. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, N.; Yu, Y.; Walsh, W.R.; Yang, J.L. Molecular targeted therapies for cancer: Sorafenib monotherapy and its combination with other therapies (Review). Oncol. Rep. 2012, 27, 1303–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, M.; Numata, K.; Hara, K.; Nozaki, A.; Fukuda, H.; Chuma, M.; Maeda, S.; Tanaka, K. Treatment of advanced hepatocellular carcinoma after failure of sorafenib treatment: subsequent or additional treatment interventions contribute to prolonged survival postprogression. Gastroenterol. Res. Pract. 2017, 2017, 5728946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.-J.; Zheng, B.; Wang, H.-Y.; Chen, L. New knowledge of the mechanisms of sorafenib resistance in liver cancer. Acta Pharmacol. Sin. 2017, 38, 614–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilhelm, S.M.; Carter, C.; Tang, L.; Wilkie, D.; McNabola, A.; Rong, H.; Chen, C.; Zhang, X.; Vincent, P.; McHugh, M.; et al. BAY 43-9006 exhibits broad spectrum oral antitumor activity and targets the RAF/MEK/ERK pathway and receptor tyrosine kinases involved in tumor progression and angiogenesis. Cancer Res. 2004, 64, 7099–7109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coriat, R.; Nicco, C.; Chereau, C.; Mir, O.; Alexandre, J.; Ropert, S.; Weill, B.; Chaussade, S.; Goldwasser, F.; Batteux, F. Sorafenib-induced hepatocellular carcinoma cell death depends on reactive oxygen species production in vitro and in vivo. Mol. Cancer 2012, 11, 2284–2293. [Google Scholar] [CrossRef] [Green Version]
- Morgensztern, D.; McLeod, H.L. PI3K/Akt/mTOR pathway as a target for cancer therapy. Anti Cancer Drug 2005, 16, 797–803. [Google Scholar] [CrossRef]
- Powis, G.; Kirkpatrick, L. Hypoxia inducible factor-1α as a cancer drug target. Mol. Cancer 2004, 3, 647–654. [Google Scholar]
- Kim, B.Y.; Cho, M.H.; Kim, K.J.; Cho, K.J.; Kim, S.W.; Kim, H.S.; Jung, W.-W.; Lee, B.H.; Lee, B.H.; Lee, S.G. Effects of PRELI in oxidative-stressed HepG2 cells. Ann. Clin. Lab. Sci. 2015, 45, 419–425. [Google Scholar]
- Song, A.; Kim, B.; Kim, E.; Sung, J.H.; Park, Y.; Park, S.; Park, T.; Kim, J.K.; Jeong, Y.; Lee, S.G. Protective Functions of Group 3 Late Embryogenesis Abundant (G3LEA) Proteins in Enterococcus faecium During Vancomycin Treatment. Indian J. Microbiol. 2020, 60, 526–534. [Google Scholar] [CrossRef]
- Zhou, X.F.; Ding, Z.S.; Liu, N.B. Allium vegetables and risk of prostate cancer: Evidence from 132,192 subjects. Asian Pac. J. Cancer Prev. 2013, 14, 4131–4134. [Google Scholar] [CrossRef] [Green Version]
- Thomson, M.; Ali, M. Garlic [Allium sativum]: A review of its potential use as an anti-cancer agent. Curr. Cancer Drug Tar. 2003, 3, 67–81. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, D.M. Role of reversible oxidation-reduction of enzyme thiols-disulfides in metabolic regulation. Annu. Rev. Biochem. 1985, 54, 305–329. [Google Scholar] [CrossRef] [PubMed]
- Hopia, A.; Heinonen, M. Antioxidant activity of flavonol aglycones and their glycosides in methyl linoleate. J. Am. Oil. Chem. Soc. 1999, 76, 139–144. [Google Scholar] [CrossRef]
- Brady, J.F.; Ishizaki, H.; Fukuto, J.M.; Lin, M.C.; Fadel, A.; Gapac, J.M.; Yang, C.S. Inhibition of cytochrome P-450 2E1 by diallyl sulfide and its metabolites. Chem. Res. Toxicol. 1991, 4, 642–647. [Google Scholar] [CrossRef] [PubMed]
- Seal, T. Quantitative HPLC analysis of phenolic acids, flavonoids and ascorbic acid in four different solvent extracts of two wild edible leaves, Sonchus arvensis and Oenanthe linearis of North-Eastern region in India. J. Appl. Pharm. Sci. 2016, 6, 157–166. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.; Lee, K.; Lee, C.; Song, A.; Kim, J.; Kim, B.; Lee, S. Protection and immune modulation of activated human vaginal epithelial cells by Aurea helianthus extract. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Marin, J.J.G.; Macias, R.I.R.; Monte, M.J.; Romero, M.R.; Asensio, M.; Sanchez-Martin, A.; Cives-Losada, C.; Temprano, A.G.; Espinosa-Escudero, R.; Reviejo, M.; et al. Molecular Bases of Drug Resistance in Hepatocellular Carcinoma. Cancers 2020, 12, 1663. [Google Scholar] [CrossRef]
- Putnik, P.; Gabrić, D.; Roohinejad, S.; Barba, F.J.; Granato, D.; Mallikarjunan, K.; Lorenzo, J.M.; Kovačević, D.B. An overview of organosulfur compounds from Allium spp.: From processing and preservation to evaluation of their bioavailability, antimicrobial, and anti-inflammatory properties. Food Chem. 2019, 276, 680–691. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Di Gioia, F.; Polyzos, N.; Tzortzakis, N. Natural antioxidants, health effects and bioactive properties of wild Allium species. Curr. Pharm. Des. 2020, 26, 1816–1837. [Google Scholar] [CrossRef]
- Vlase, L.; Parvu, M.; Parvu, E.A.; Toiu, A. Chemical constituents of three Allium species from Romania. Molecules 2013, 18, 114–127. [Google Scholar] [CrossRef]
- Kilic, I.; Yesiloglu, Y. Spectroscopic studies on the antioxidant activity of p-coumaric acid. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2013, 115, 719–724. [Google Scholar] [CrossRef]
- Wang, S.-Q.; Li, X.; Wang, X.-N.; Wei, N.-N.; Lou, H.-X. Coumarins from Cicuta virosa and their modulating effects on multidrug-resistant (MDR) tumors. Phytochem. Lett. 2011, 4, 97–100. [Google Scholar] [CrossRef]
- Senthilkumar, R.; Bao-An, C.; Xiao-Hui, C.; Rong, F. Anticancer and multidrug-resistance reversing potential of traditional medicinal plants and their bioactive compounds in leukemia cell lines. Chin. J. Nat. Med. 2014, 12, 881–894. [Google Scholar] [CrossRef]
- Han, C.; Liao, X.; Qin, W.; Yu, L.; Liu, X.; Chen, G.; Liu, Z.; Lu, S.; Chen, Z.; Su, H.; et al. EGFR and SYNE2 are associated with p21 expression and SYNE2 variants predict post-operative clinical outcomes in HBV-related hepatocellular carcinoma. Sci. Rep. 2016, 6, 31237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabral, L.K.D.; Tiribelli, C.; Sukowati, C.H. Sorafenib Resistance in Hepatocellular Carcinoma: The Relevance of Genetic Heterogeneity. Cancers 2020, 12, 1576. [Google Scholar] [CrossRef]
- Prieto-Domínguez, N.; Ordóñez, R.; Fernández, A.; García-Palomo, A.; Muntané, J.; González-Gallego, J.; Mauriz, J.L. Modulation of autophagy by sorafenib: Effects on treatment response. Front. Pharmacol. 2016, 7, 151. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Modeling of sorafenib resistance and levels of PRELI in hepatocarcinoma cells: (a) modeling of resistant cells; comparison of cellular viability between susceptible and resistant cells exposed to sorafenib; (b) levels of PRELI in the susceptible and resistant cells (* p < 0.05, ** p < 0.01, *** p < 0.001) (error bars = standard deviation).
Figure 1.
Modeling of sorafenib resistance and levels of PRELI in hepatocarcinoma cells: (a) modeling of resistant cells; comparison of cellular viability between susceptible and resistant cells exposed to sorafenib; (b) levels of PRELI in the susceptible and resistant cells (* p < 0.05, ** p < 0.01, *** p < 0.001) (error bars = standard deviation).

Figure 2.
Quantity of p-coumaric acid in Allium senescens extract: (a) detection of bioactive compounds in A. senescens extract; (b,c) standard curves of p-coumaric acid; and (d) the measurement of p-coumaric acid in the A. senescens extract using ultra-high performance liquid chromatography (UHPLC) (p < 0.05).
Figure 2.
Quantity of p-coumaric acid in Allium senescens extract: (a) detection of bioactive compounds in A. senescens extract; (b,c) standard curves of p-coumaric acid; and (d) the measurement of p-coumaric acid in the A. senescens extract using ultra-high performance liquid chromatography (UHPLC) (p < 0.05).

Figure 3.
mRNA Levels of PRELI and ABC transporters in drug resistant hepatocarcinoma cells treated with p-coumaric acid and Allium senescens extract: (a) levels of PRELI in susceptible and resistant cells treated with p-coumaric acid (CA); (b) levels of PRELI in susceptible and resistant cells treated with A. senescens extract (A.S.); (c) levels of efflux transporters in the resistant cells treated with p-coumaric acid; and (d) levels of influx transporters in the resistant cells treated with A. senescens extract. S, susceptible cells; R, resistant cells; ns, not significant (* p < 0.05, ** p < 0.01) (error bars = standard deviation).
Figure 3.
mRNA Levels of PRELI and ABC transporters in drug resistant hepatocarcinoma cells treated with p-coumaric acid and Allium senescens extract: (a) levels of PRELI in susceptible and resistant cells treated with p-coumaric acid (CA); (b) levels of PRELI in susceptible and resistant cells treated with A. senescens extract (A.S.); (c) levels of efflux transporters in the resistant cells treated with p-coumaric acid; and (d) levels of influx transporters in the resistant cells treated with A. senescens extract. S, susceptible cells; R, resistant cells; ns, not significant (* p < 0.05, ** p < 0.01) (error bars = standard deviation).

Figure 4.
RNA levels of drug transporters in hepatocarcinoma cells following PRELI knockdown: (a) the levels of drug transporters after PRELI knockdown; (b) expressive modulation of drug transporters by the Allium senescens extract (A.S.) in the PRELI-knocked down cells. S, susceptible cells; R, resistant cells; ns, not significant, exposure time for all substances was 3 days (* p < 0.05, ** p < 0.01) (error bars = standard deviation).
Figure 4.
RNA levels of drug transporters in hepatocarcinoma cells following PRELI knockdown: (a) the levels of drug transporters after PRELI knockdown; (b) expressive modulation of drug transporters by the Allium senescens extract (A.S.) in the PRELI-knocked down cells. S, susceptible cells; R, resistant cells; ns, not significant, exposure time for all substances was 3 days (* p < 0.05, ** p < 0.01) (error bars = standard deviation).

Figure 5.
Protein levels of drug transporters in hepatocarcinoma cells following PRELI knockdown; (a) estimating PRELI levels to confirm PRELI knockdown; (b–d) expression of the efflux transporter ABCB1, AGCC2 and ABCC2 in PRELI-knocked down cells; (e) expression of the influx transporter OCT-1 in PRELI-knocked down cells. A.S., Allium senescens extract; S, susceptible cells; R, resistant cells; knock, PRELI knock down; ns, not significant (* p < 0.05, ** p < 0.01) (error bars = standard deviation).
Figure 5.
Protein levels of drug transporters in hepatocarcinoma cells following PRELI knockdown; (a) estimating PRELI levels to confirm PRELI knockdown; (b–d) expression of the efflux transporter ABCB1, AGCC2 and ABCC2 in PRELI-knocked down cells; (e) expression of the influx transporter OCT-1 in PRELI-knocked down cells. A.S., Allium senescens extract; S, susceptible cells; R, resistant cells; knock, PRELI knock down; ns, not significant (* p < 0.05, ** p < 0.01) (error bars = standard deviation).

Figure 6.
RNA levels of apoptotic markers in hepatocarcinoma cells treated with Allium senescens extract. The markers in panels (a–c) are involved in autophagy, while those in panels (d–f) are apoptotic signals in the mitochondria. A.S., Allium senescens extract; ns, not significant (* p < 0.05, ** p < 0.01, *** p < 0.001) (error bars = standard deviation).
Figure 6.
RNA levels of apoptotic markers in hepatocarcinoma cells treated with Allium senescens extract. The markers in panels (a–c) are involved in autophagy, while those in panels (d–f) are apoptotic signals in the mitochondria. A.S., Allium senescens extract; ns, not significant (* p < 0.05, ** p < 0.01, *** p < 0.001) (error bars = standard deviation).

Figure 7.
Localization of PRELI in hepatocarcinoma cells. All panels show the patterns of PRELI expression in various cell types: (a) images for the localization of PRELI in hepatocarcinoma cells; (b) histograms for counting of PRELI granules; the granules indicate accumulated PRELI. nc, negative control (cells without sorafenib); A.S, Allium senescens extract; SC, susceptible cells; RC, resistant cells; CM, cytoplasmic membrane; ns, not significant, scale bars = 30μm, (* p < 0.05, ** p < 0.01) (error bars = standard deviation).
Figure 7.
Localization of PRELI in hepatocarcinoma cells. All panels show the patterns of PRELI expression in various cell types: (a) images for the localization of PRELI in hepatocarcinoma cells; (b) histograms for counting of PRELI granules; the granules indicate accumulated PRELI. nc, negative control (cells without sorafenib); A.S, Allium senescens extract; SC, susceptible cells; RC, resistant cells; CM, cytoplasmic membrane; ns, not significant, scale bars = 30μm, (* p < 0.05, ** p < 0.01) (error bars = standard deviation).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Park, S.; Park, Y.; Shin, H.; Kim, B.; Lee, S. Effect of Allium senescens Extract on Sorafenib Resistance in Hepatocarcinoma Cells. Appl. Sci. 2021, 11, 3696. https://0-doi-org.brum.beds.ac.uk/10.3390/app11083696
AMA Style
Park S, Park Y, Shin H, Kim B, Lee S. Effect of Allium senescens Extract on Sorafenib Resistance in Hepatocarcinoma Cells. Applied Sciences. 2021; 11(8):3696. https://0-doi-org.brum.beds.ac.uk/10.3390/app11083696
Chicago/Turabian StylePark, Sohyeon, Yoonjin Park, Heejong Shin, Boyong Kim, and Seunggwan Lee. 2021. "Effect of Allium senescens Extract on Sorafenib Resistance in Hepatocarcinoma Cells" Applied Sciences 11, no. 8: 3696. https://0-doi-org.brum.beds.ac.uk/10.3390/app11083696
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.