
Improved Electrical Stimulation-Based Exercise Model to Induce Mice Tibialis Anterior Muscle Hypertrophy and Function

 ,
,  ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Groups
2.2. Design of Electrical Stimulation Protocol
2.3. The Cross-Sectional Area of a Fiber (CSA)
2.4. Myonuclei Counting
2.5. In Vivo Muscle Function Analysis
2.6. Immunofluorescence Analysis
2.7. Western-Blot
2.8. Statistical Analysis
3. Results
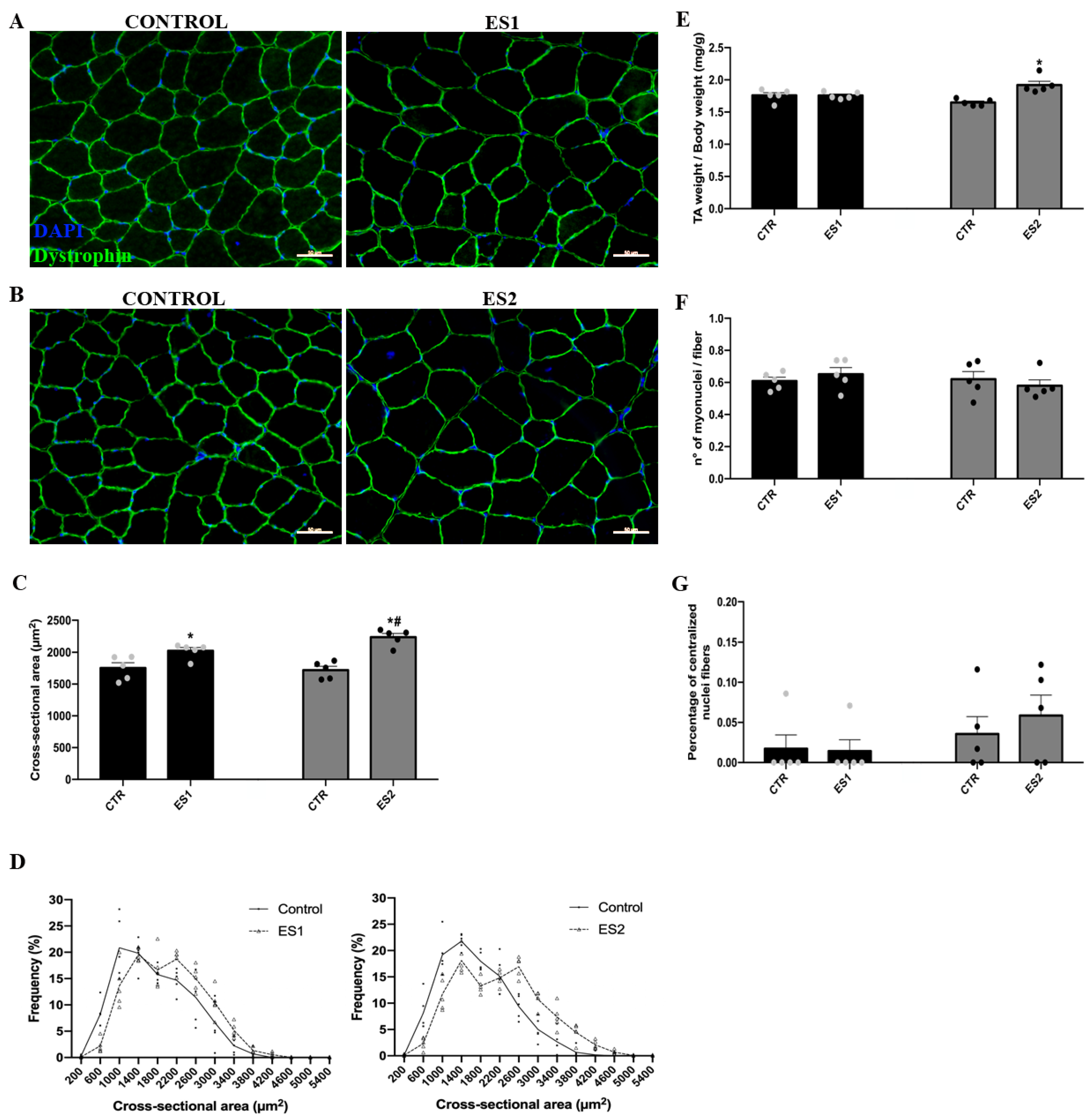
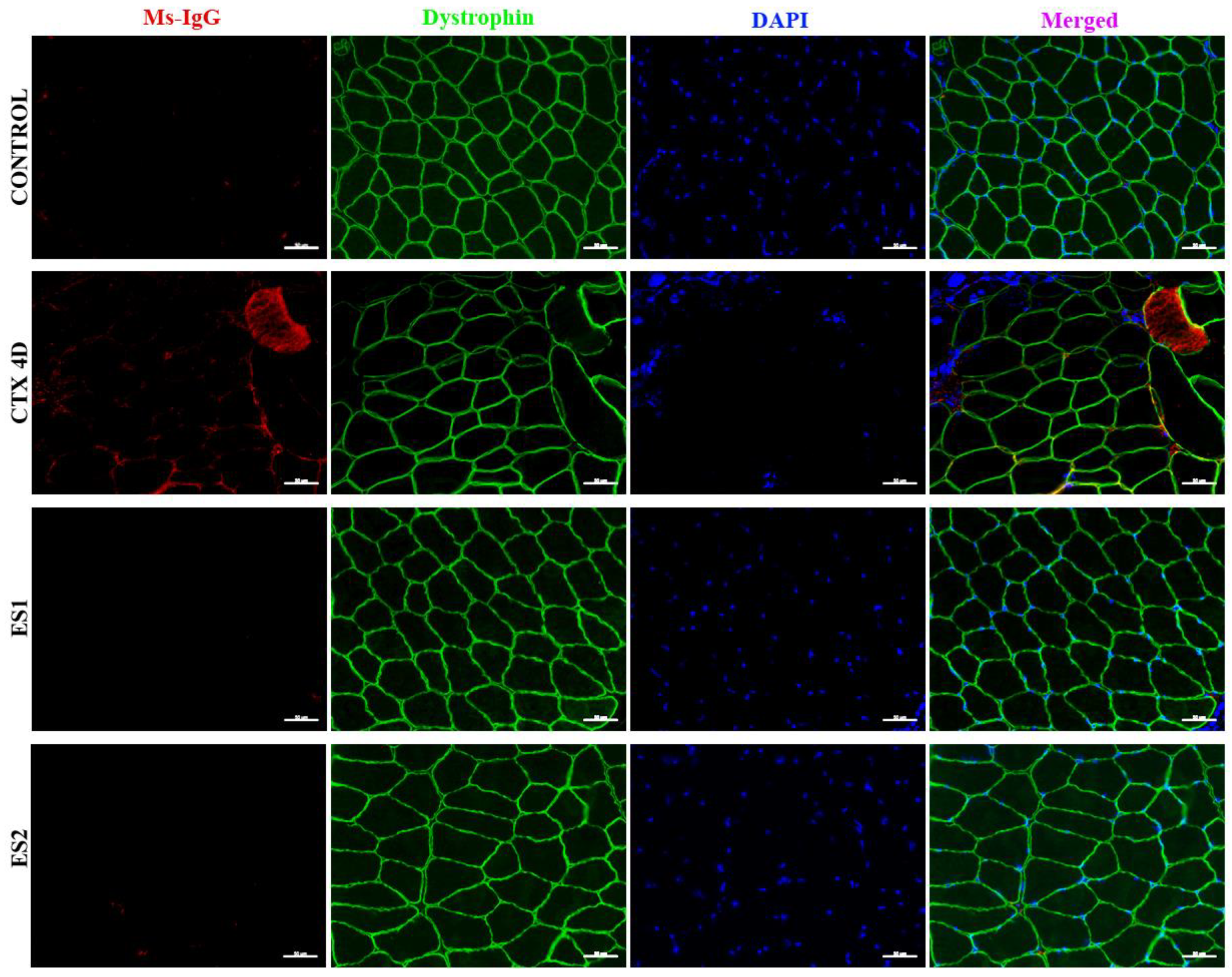
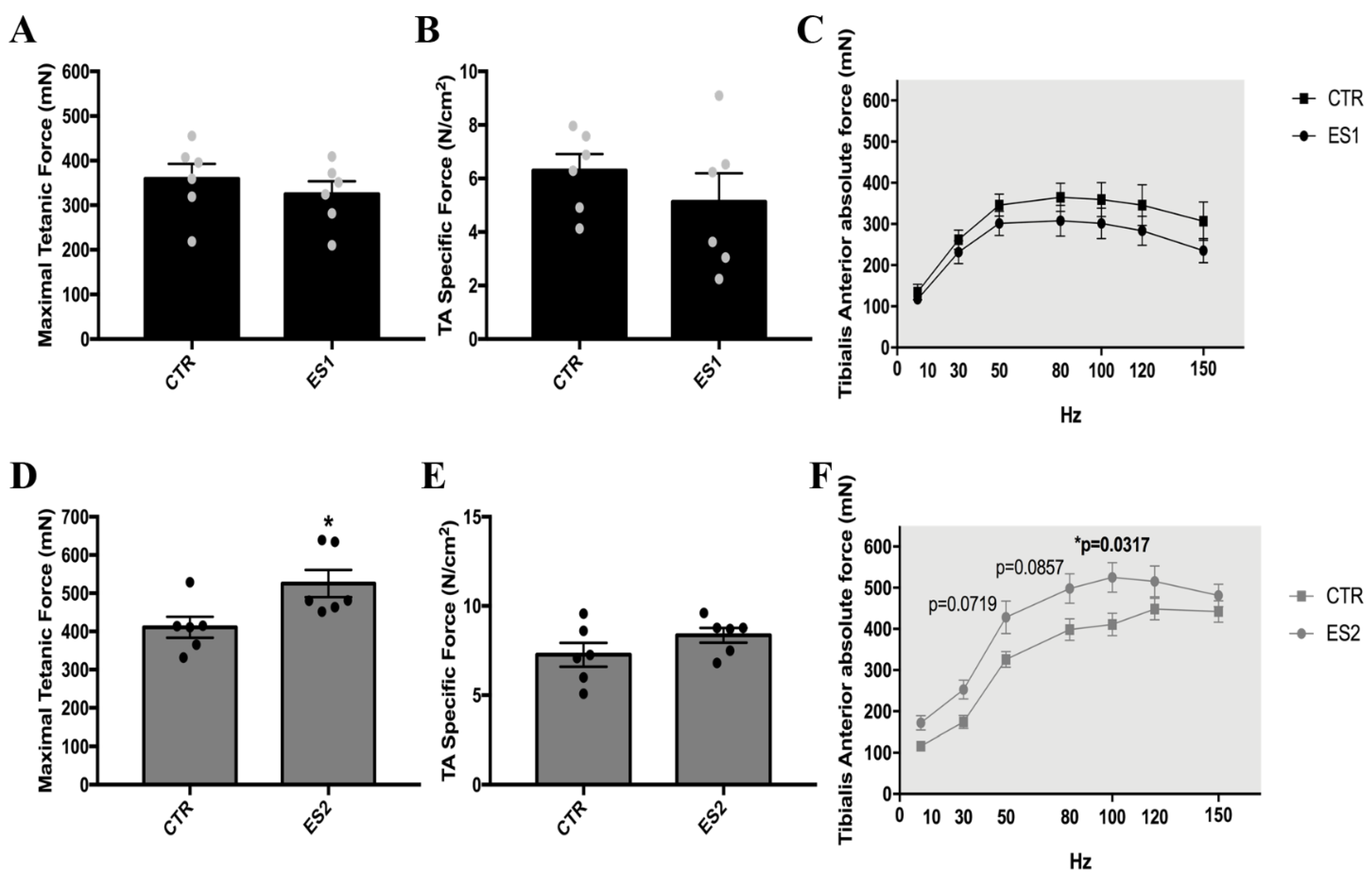
3.1. Electrical Stimulation Induced Skeletal Muscle Hypertrophy and Enhanced Tibialis Anterior Maximal Strength in Mice
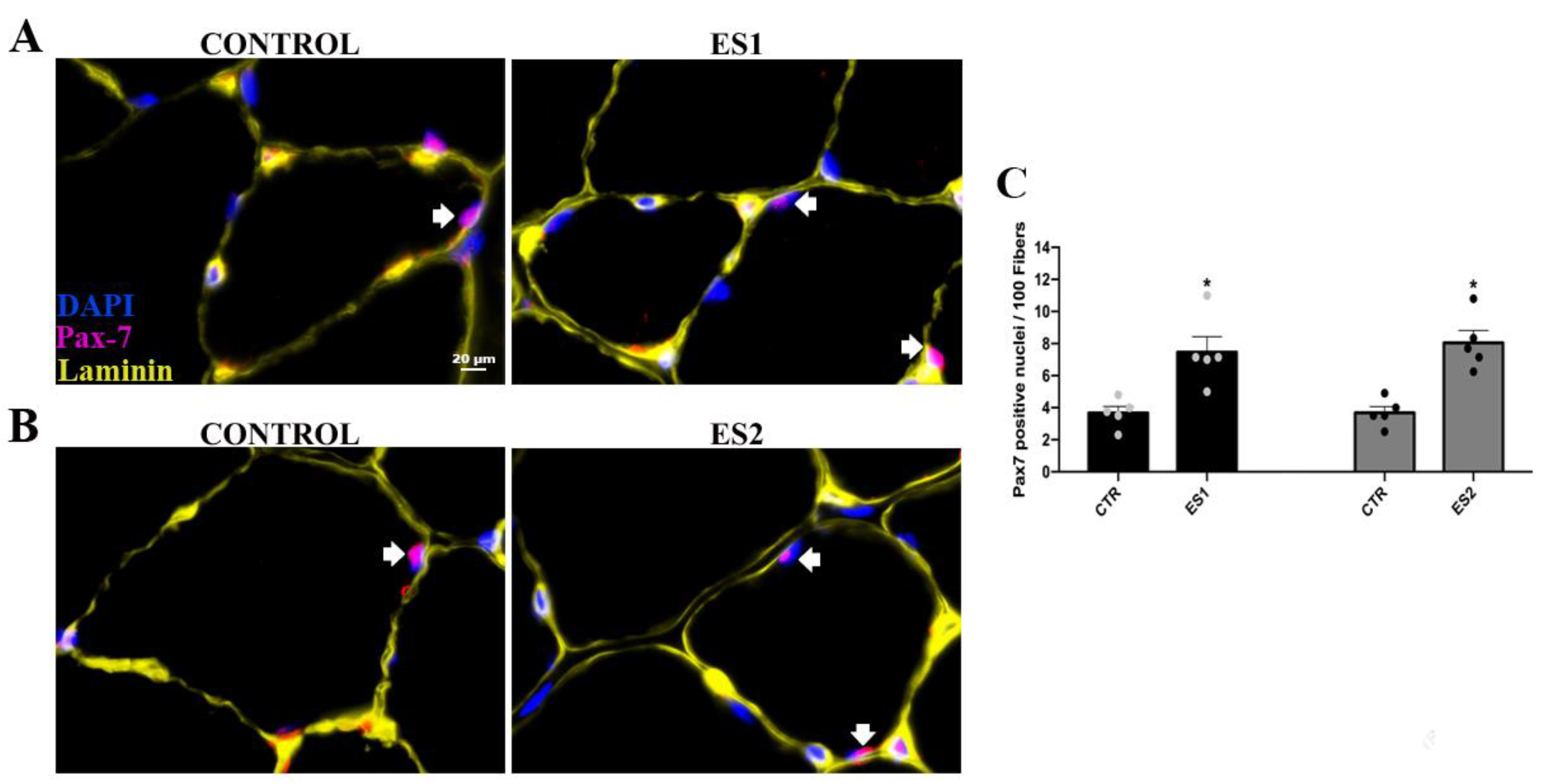
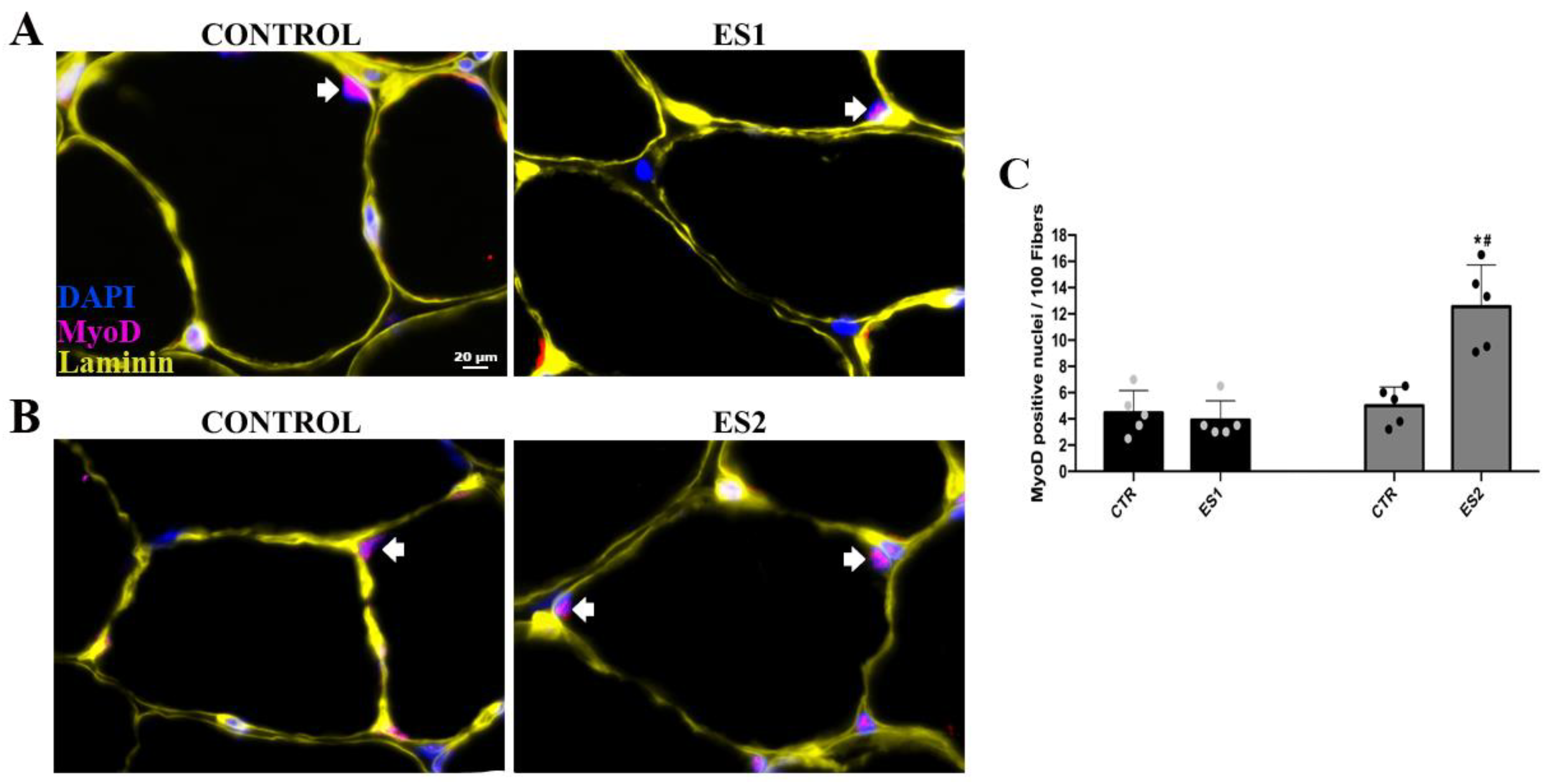
3.2. Electrical Stimulation Enhanced Pax-7 and MyoD Counts in Tibialis Anterior of Mice
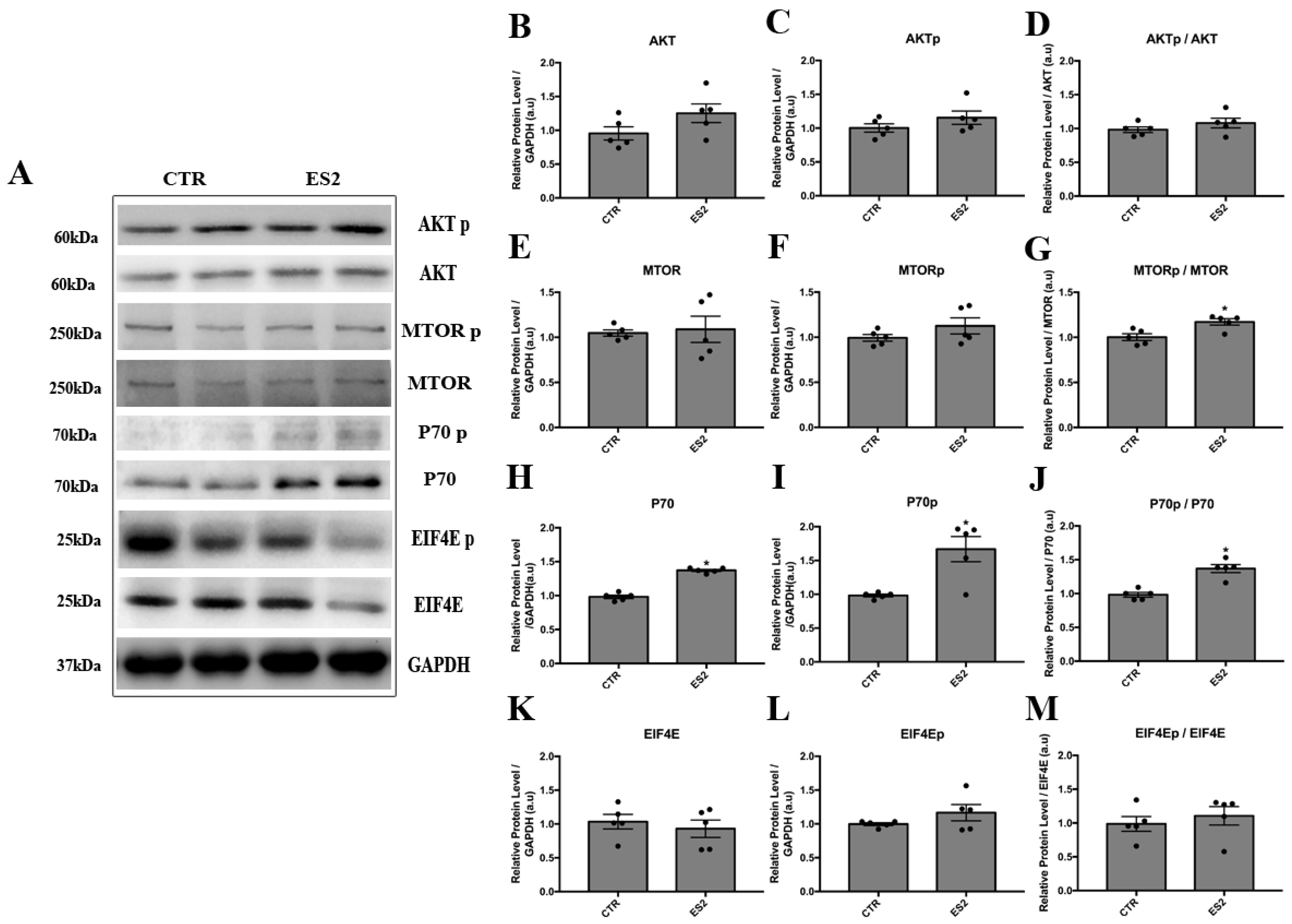
3.3. Electrical Stimulation Induced AKT/mTOR Signaling
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Harber, M.P.; Konopka, A.R.; Undem, M.K.; Hinkley, J.M.; Minchev, K.; Kaminsky, L.A.; Trappe, T.A.; Trappe, S. Aerobic Exercise Training Induces Skeletal Muscle Hypertrophy and Age-Dependent Adaptations in Myofiber Function in Young and Older Men. J. Appl. Physiol. 2012, 113, 1495–1504. [Google Scholar] [CrossRef] [Green Version]
- Hardee, J.P.; Mangum, J.E.; Gao, S.; Sato, S.; Hetzler, K.L.; Puppa, M.J.; Fix, D.K.; Carson, J.A. Eccentric Contraction-Induced Myofiber Growth in Tumor-Bearing Mice. J. Appl. Physiol. 2016, 120, 29–37. [Google Scholar] [CrossRef]
- Huang, Z.; Fang, Q.; Ma, W.; Zhang, Q.; Qiu, J.; Gu, X.; Yang, H.; Sun, H. Skeletal Muscle Atrophy Was Alleviated by Salidroside through Suppressing Oxidative Stress and Inflammation during Denervation. Front. Pharmacol. 2019, 10, 997. [Google Scholar] [CrossRef]
- Schoenfeld, B.J.; Contreras, B.; Krieger, J.; Grgic, J.; Delcastillo, K.; Belliard, R.; Alto, A. Resistance Training Volume Enhances Muscle Hypertrophy but Not Strength in Trained Men. Med. Sci. Sports Exerc. 2019, 51, 94–103. [Google Scholar] [CrossRef]
- Alves, P.K.N.; Cruz, A.; Silva, W.J.; Labeit, S.; Moriscot, A.S. Leucine Supplementation Decreases HDAC4 Expression and Nuclear Localization in Skeletal Muscle Fiber of Rats Submitted to Hindlimb Immobilization. Cells 2020, 9, 2582. [Google Scholar] [CrossRef]
- Adams, V.; Bowen, T.S.; Werner, S.; Barthel, P.; Amberger, C.; Konzer, A.; Graumann, J.; Sehr, P.; Lewis, J.; Provaznik, J.; et al. Small-Molecule-Mediated Chemical Knock-down of MuRF1/MuRF2 and Attenuation of Diaphragm Dysfunction in Chronic Heart Failure. J. Cachexia Sarcopenia Muscle 2019, 10, 1102–1115. [Google Scholar] [CrossRef]
- Debevec, T.; Ganse, B.; Mittag, U.; Eiken, O.; Mekjavic, I.B.; Rittweger, J. Hypoxia Aggravates Inactivity-Related Muscle Wasting. Front. Physiol. 2018, 9, 494. [Google Scholar] [CrossRef] [Green Version]
- Glass, D.J. Signalling Pathways That Mediate Skeletal Muscle Hypertrophy and Atrophy. Nat. Cell Biol. 2003, 5, 87–90. [Google Scholar] [CrossRef]
- Margolis, L.M.; Rivas, D.A. Implications of Exercise Training and Distribution of Protein Intake on Molecular Processes Regulating Skeletal Muscle Plasticity. Behav. Genet. 2015, 45, 211–221. [Google Scholar] [CrossRef]
- Tipton, K.D.; Wolfe, R.R. Exercise, Protein Metabolism, and Muscle Growth. Int. J. Sport Nutr. Exerc. Metab. 2001, 11, 109–132. [Google Scholar] [CrossRef] [Green Version]
- Ogasawara, R.; Fujita, S.; Hornberger, T.A.; Kitaoka, Y.; Makanae, Y.; Nakazato, K.; Naokata, I. The Role of MTOR Signalling in the Regulation of Skeletal Muscle Mass in a Rodent Model of Resistance Exercise. Sci. Rep. 2016, 6, 31142. [Google Scholar] [CrossRef]
- Takegaki, J.; Ogasawara, R.; Tamura, Y.; Takagi, R.; Arihara, Y.; Tsutaki, A.; Nakazato, K.; Ishii, N. Repeated Bouts of Resistance Exercise with Short Recovery Periods Activates MTOR Signaling, but Not Protein Synthesis, in Mouse Skeletal Muscle. Physiol. Rep. 2017, 5, e13515. [Google Scholar] [CrossRef] [PubMed]
- Bodine, S.C.; Baar, K. Analysis of Skeletal Muscle Hypertrophy in Models of Increased Loading. Methods Mol. Biol. 2012, 798, 213–229. [Google Scholar] [CrossRef] [PubMed]
- Communications, S. Progression Models in Resistance Training for Healthy Adults. Med. Sci. Sports Exerc. 2009, 41, 687–708. [Google Scholar] [CrossRef]
- Murach, K.A.; McCarthy, J.J.; Peterson, C.A.; Dungan, C.M. Making Mice Mighty: Recent Advances in Translational Models of Load-Induced Muscle Hypertrophy. J. Appl. Physiol. 2020, 129, 516–521. [Google Scholar] [CrossRef]
- Terena, S.M.L.; Fernandes, K.P.S.; Bussadori, S.K.; Deana, A.M.; Mesquita-Ferrari, R.A. Systematic Review of the Synergist Muscle Ablation Model for Compensatory Hypertrophy. Rev. Assoc. Med. Bras. 2017, 63, 164–172. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, R.B.; Marum, P.; Tullson, P.; Saubert IV, C.W. Acute Hypertrophic Response of Skeletal Muscle to Removal of Synergists. J. Appl. Physiol. 1979, 46, 835–842. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, K.M.; Cheadle, W.G.; Martinez, O.M.; Cooke, D.A. Effect of Functional Overload on Enzyme Levels in Different Types of Skeletal Muscle. J. Appl. Phys. Resp. Environ. Exerc. Physiol. 1977, 42, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.R.; Meadows, I.D.; Baldwin, K.M.; Edgerton, V.R. Functional Significance of Compensatory Overloaded Rat Fast Muscle. J. Appl. Physiol. 1982, 52, 473–478. [Google Scholar] [CrossRef]
- Tamaki, T.; Uchiyama, S.; Nakano, S. A Weight-Lifting Exercise Model for Inducing Hypertrophy in the Hindlimb Muscles of Rats. Med. Sci. Sports Exerc. 1992, 24, 881–886. [Google Scholar] [CrossRef]
- Barretti, D.L.M.; Melo, S.F.S.; Oliveira, E.M.; Barauna, V.G. Resistance Training Attenuates Salt Overload-Induced Cardiac Remodeling and Diastolic Dysfunction in Normotensive Rats. Braz. J. Med. Biol. Res. 2017, 50, e6146. [Google Scholar] [CrossRef] [Green Version]
- Mota, M.M.; da Silva, T.L.T.B.; Fontes, M.T.; Barreto, A.S.; dos Santos Araújo, J.E.; de Oliveira, A.C.C.; Wichi, R.B.; Santos, M.R.V. Resistance Exercise Restores Endothelial Function and Reduces Blood Pressure in Type 1 Diabetic Rats. Arq. Bras. Cardiol. 2014, 103, 25–32. [Google Scholar] [CrossRef]
- Alves, J.P.; Nunes, R.B.; da Cunha Ferreira, D.; Stefani, G.P.; Jaenisch, R.B.; Dal Lago, P. High-Intensity Resistance Training Alone or Combined with Aerobic Training Improves Strength, Heart Function and Collagen in Rats with Heart Failure. Am. J. Transl. Res. 2017, 9, 5432–5441. [Google Scholar] [PubMed]
- Aguiar, A.F.; Vechetti-Júnior, I.J.; Alves De Souza, R.W.; Castan, E.P.; Milanezi-Aguiar, R.C.; Padovani, C.R.; Carvalho, R.F.; Silva, M.D.P. Myogenin, MyoD and IGF-I Regulate Muscle Mass but Not Fiber-Type Conversion during Resistance Training in Rats. Int. J. Sports Med. 2013, 34, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Cutlip, R.G.; Baker, B.A.; Geronilla, K.B.; Mercer, R.R.; Kashon, M.L.; Miller, G.R.; Murlasits, Z.; Alway, S.E. Chronic Exposure to Stretch–Shortening Contractions Results in Skeletal Muscle Adaptation in Young Rats and Maladaptation in Old Rats. Appl. Physiol. Nutr. Metab. 2006, 31, 573–587. [Google Scholar] [CrossRef]
- Rader, E.P.; Layner, K.; Triscuit, A.M.; Chetlin, R.D.; Ensey, J.; Baker, B.A. Age-Dependent Muscle Adaptation after Chronic Stretch-Shortening Contractions in Rats. Aging Dis. 2016, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rader, E.P.; Naimo, M.A.; Ensey, J.; Baker, B.A. High-Intensity Stretch-Shortening Contraction Training Modifies Responsivity of Skeletal Muscle in Old Male Rats. Exp. Gerontol. 2018, 104, 118–126. [Google Scholar] [CrossRef]
- Duncan, N.D.; Williams, D.A.; Lynch, G.S. Adaptations in Rat Skeletal Muscle Following Long-Term Resistance Exercise Training. Eur. J. Appl. Physiol. 1998, 77, 372–378. [Google Scholar] [CrossRef]
- Yarasheski, K.E.; Lemon, P.W.R.; Gilloteaux, J. Effect of Heavy-Resistance Exercise Training on Muscle Fiber Composition in Young Rats. J. Appl. Physiol. 1990, 69, 434–437. [Google Scholar] [CrossRef]
- Hornberger, T.A.; Farrar, R.P. Physiological Hypertrophy of the FHL Muscle Following 8 Weeks of Progressive Resistance Exercise in the Rat. Can. J. Appl. Physiol. 2004, 29, 16–31. [Google Scholar] [CrossRef]
- Jung, S.; Ahn, N.; Kim, S.; Byun, J.; Joo, Y.; Kim, S.; Jung, Y.; Park, S.; Hwang, I.; Kim, K. The Effect of Ladder-Climbing Exercise on Atrophy/Hypertrophy-Related Myokine Expression in Middle-Aged Male Wistar Rats. J. Physiol. Sci. 2015, 65, 515–521. [Google Scholar] [CrossRef]
- Speretta, G.F.F.; Rosante, M.C.; Duarte, F.O.; Leite, R.D.; de Souza Lino, A.D.; Andre, R.A.; de Oliviera Silvestre, J.G.; de Araujo, H.S.S.; de Oliveira Duarte, A.C.G. The Effects of Exercise Modalities on Adiposity in Obese Rats. Clinics 2012, 67, 1469–1477. [Google Scholar] [CrossRef]
- Silvestre, J.G.; Speretta, G.F.F.; Fabrizzi, F.; Moraes, G.; De Oliveira Duarte, A.C.G. Acute Effects of Resistance Exercise Performed on Ladder on Energy Metabolism, Stress, and Muscle Damage in Rats. Mot. Rev. Educ. Fis. 2017, 23. [Google Scholar] [CrossRef]
- Wong, T.S.; Booth, F.W. Protein Metabolism in Rat Tibialis Anterior Muscle after Stimulated Chronic Eccentric Exercise. J. Appl. Physiol. 1990, 69, 1718–1724. [Google Scholar] [CrossRef]
- Wong, T.S.; Booth, F.W. Skeletal Muscle Enlargement with Weight-Lifting Exercise by Rats. J. Appl. Physiol. 1988, 65, 950–954. [Google Scholar] [CrossRef]
- Baar, K.; Esser, K. Phosphorylation of P70(S6k) Correlates with Increased Skeletal Muscle Mass Following Resistance Exercise. Am. J. Physiol. Cell Physiol. 1999, 276, 120–127. [Google Scholar] [CrossRef]
- Hardee, J.P.; Counts, B.R.; Gao, S.; VanderVeen, B.N.; Fix, D.K.; Koh, H.J.; Carson, J.A. Inflammatory Signalling Regulates Eccentric Contraction-Induced Protein Synthesis in Cachectic Skeletal Muscle. J. Cachexia Sarcopenia Muscle 2018, 9, 369–383. [Google Scholar] [CrossRef]
- Ogasawara, R.; Kobayashi, K.; Tsutaki, A.; Lee, K.; Abe, T.; Fujita, S.; Nakazato, K.; Ishii, N. MTOR Signaling Response to Resistance Exercise Is Altered by Chronic Resistance Training and Detraining in Skeletal Muscle. J. Appl. Physiol. 2013, 114, 934–940. [Google Scholar] [CrossRef] [Green Version]
- Ambrosio, F.; Kelley Fitzgerald, G.; Ferrari, R.; Distefano, G.; Carvell, G. A Murine Model of Muscle Training by Neuromuscular Electrical Stimulation. J. Vis. Exp. 2012, 63, e3914. [Google Scholar] [CrossRef] [Green Version]
- Brown, M.D.; Cotter, M.A.; Hudlická, O.; Vrbová, G. The Effects of Different Patterns of Muscle Activity on Capillary Density, Mechanical Properties and Structure of Slow and Fast Rabbit Muscles. Pflüg. Arch. Eur. J. Physiol. 1976, 361, 241–250. [Google Scholar] [CrossRef]
- Liu, F.; Fry, C.S.; Mula, J.; Jackson, J.R.; Lee, J.D.; Peterson, C.A.; Yang, L. Automated Fiber-Type-Specific Cross-Sectional Area Assessment and Myonuclei Counting in Skeletal Muscle. J. Appl. Physiol. 2013, 115, 1714–1724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, W.J.; Graça, F.A.; Cruz, A.; Silvestre, J.G.; Labeit, S.; Miyabara, E.H.; Yan, C.Y.I.; Wang, D.Z.; Moriscot, A.S. MiR-29c Improves Skeletal Muscle Mass and Function throughout Myocyte Proliferation and Differentiation and by Repressing Atrophy-Related Genes. Acta Physiol. 2019, 226, e13278. [Google Scholar] [CrossRef] [PubMed]
- Distefano, G.; Ferrari, R.J.; Weiss, C.; Deasy, B.M.; Boninger, M.L.; Fitzgerald, G.K.; Huard, J.; Ambrosio, F. Neuromuscular Electrical Stimulation as a Method to Maximize the Beneficial Effects of Muscle Stem Cells Transplanted into Dystrophic Skeletal Muscle. PLoS ONE 2013, 8, e54922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roche, J.A.; Tulapurkar, M.E.; Mueller, A.L.; van Rooijen, N.; Hasday, J.D.; Lovering, R.M.; Bloch, R.J. Myofiber Damage Precedes Macrophage Infiltration after in Vivo Injury in Dysferlin-Deficient A/J Mouse Skeletal Muscle. Am. J. Pathol. 2015, 185, 1686–1698. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alves, P.K.N.; Silvestre, J.G.; Sougey, W.W.D.; Cruz, A.; Moriscot, A.S. Improved Electrical Stimulation-Based Exercise Model to Induce Mice Tibialis Anterior Muscle Hypertrophy and Function. Appl. Sci. 2022, 12, 7673. https://0-doi-org.brum.beds.ac.uk/10.3390/app12157673
Alves PKN, Silvestre JG, Sougey WWD, Cruz A, Moriscot AS. Improved Electrical Stimulation-Based Exercise Model to Induce Mice Tibialis Anterior Muscle Hypertrophy and Function. Applied Sciences. 2022; 12(15):7673. https://0-doi-org.brum.beds.ac.uk/10.3390/app12157673
Chicago/Turabian StyleAlves, Paula Ketilly Nascimento, João G. Silvestre, Wenddy Wyllie Damascena Sougey, André Cruz, and Anselmo Sigari Moriscot. 2022. "Improved Electrical Stimulation-Based Exercise Model to Induce Mice Tibialis Anterior Muscle Hypertrophy and Function" Applied Sciences 12, no. 15: 7673. https://0-doi-org.brum.beds.ac.uk/10.3390/app12157673