
Members of the AP-1 Family of Transcription Factors Regulate the Expression of Gja1 in Mouse GC-1 Spermatogonial Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plasmids
2.3. Cell Culture and Transfections
2.4. RNA-Seq Analyses
2.5. Chromatin Immunoprecipitation Assays
2.6. Western Blot Assays
2.7. Statistical Analyses
3. Results
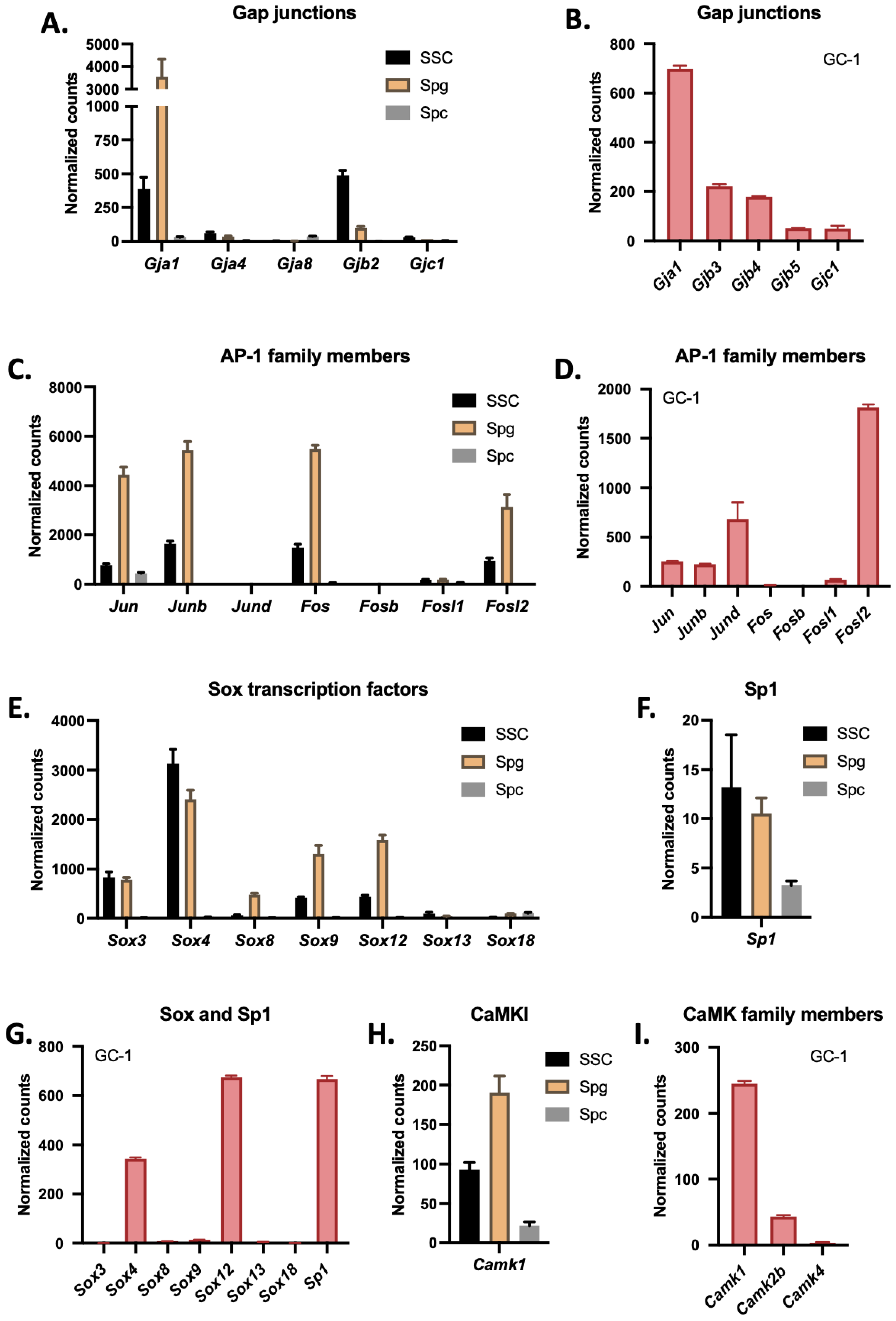
3.1. Gja1 Is the Most Highly Expressed Connexin in Spermatogonia and GC-1 Cells
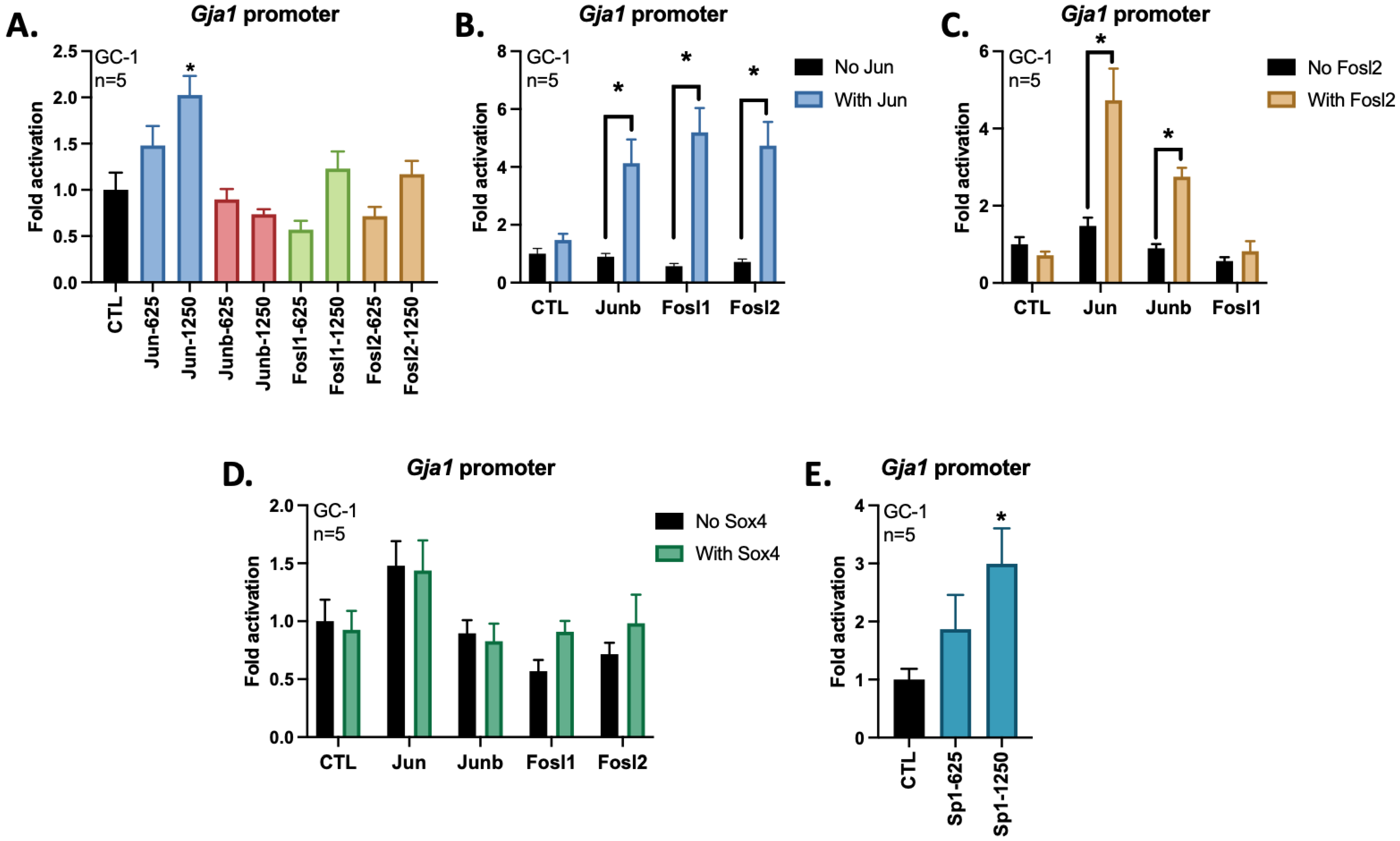
3.2. AP-1 Members Jun and Fosl2 Cooperate to Activate the Gja1 Promoter in GC-1 Cells
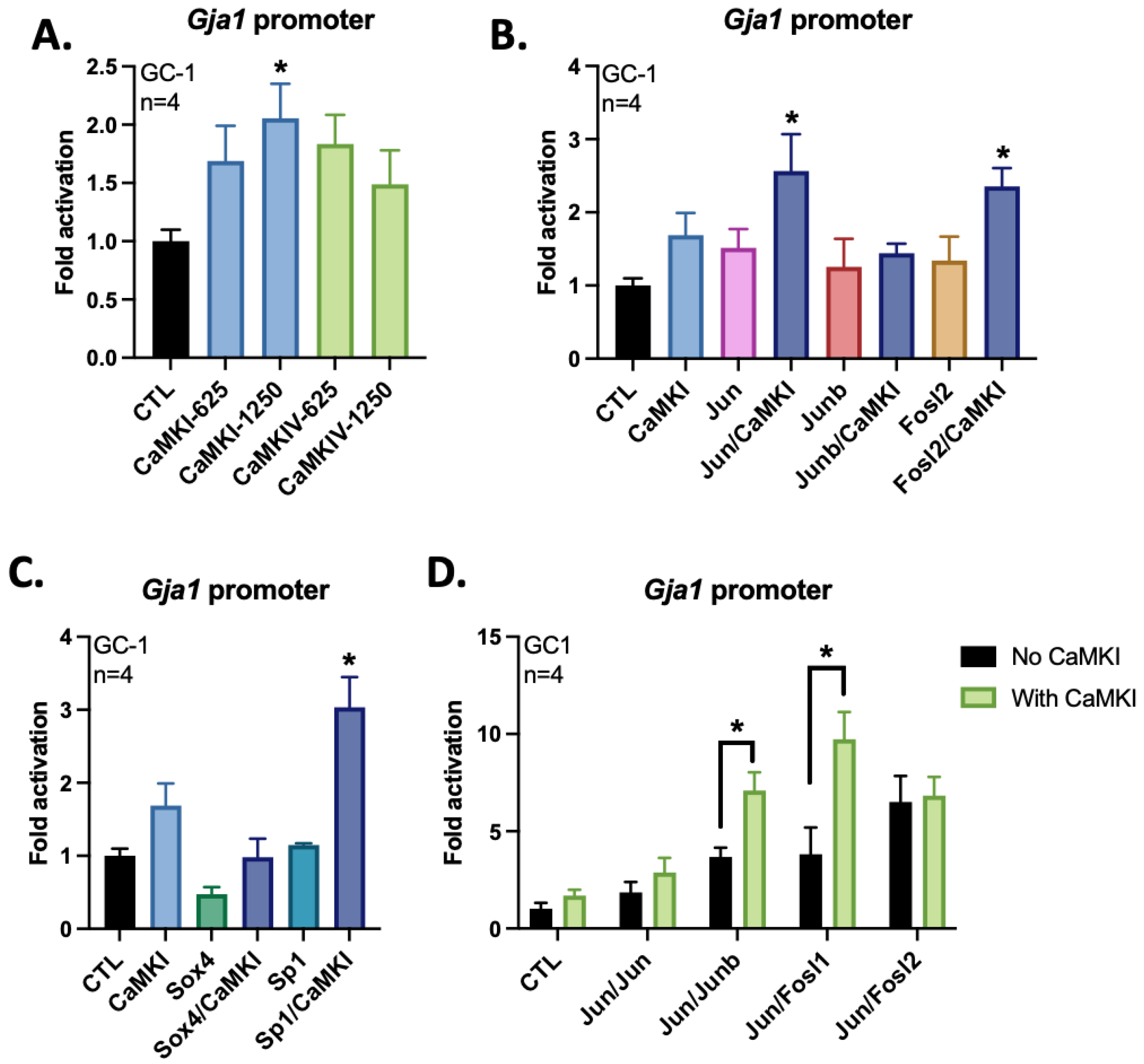
3.3. CaMKI Activates the Gja1 Promoter in GC-1 Cells
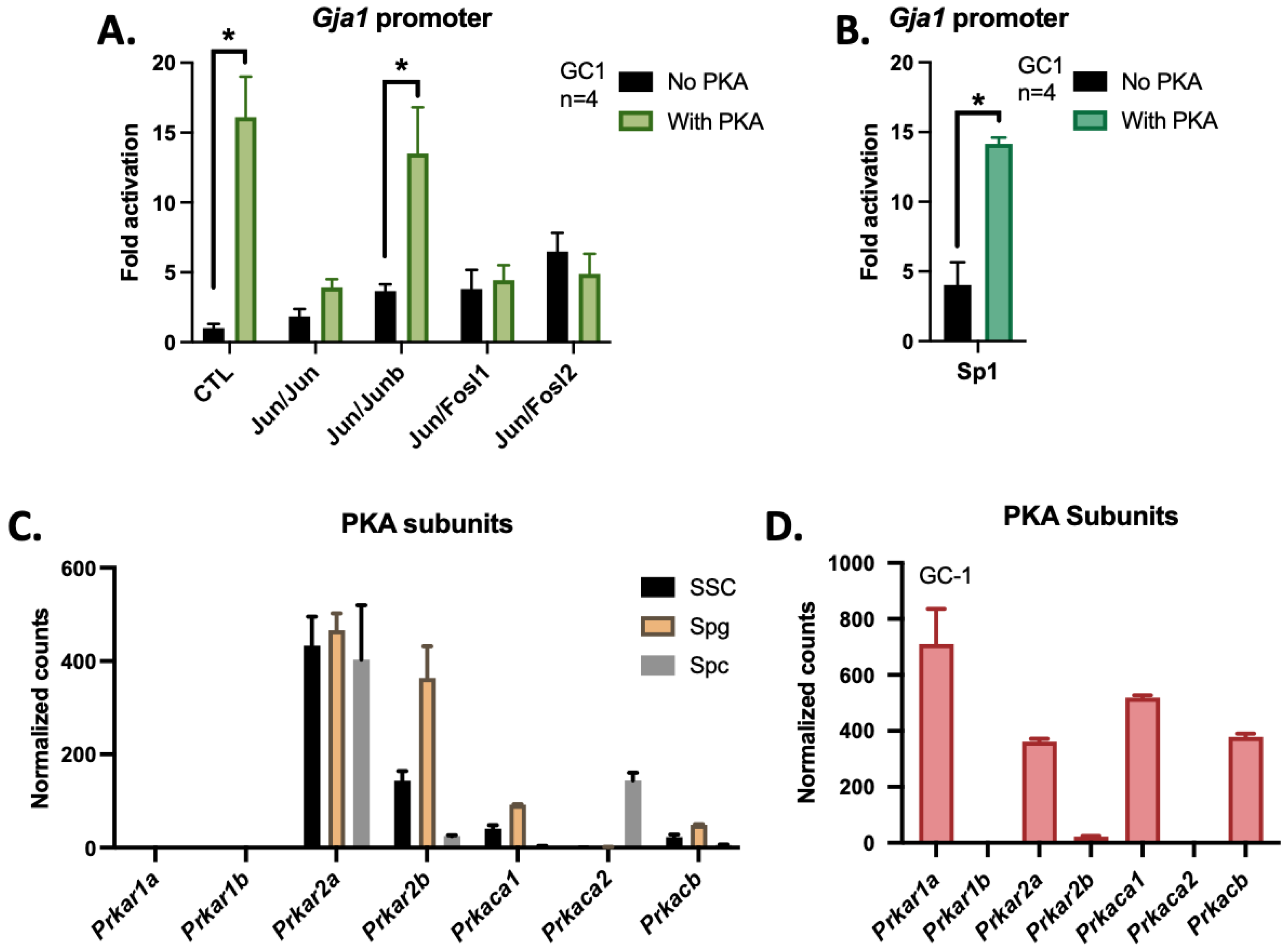
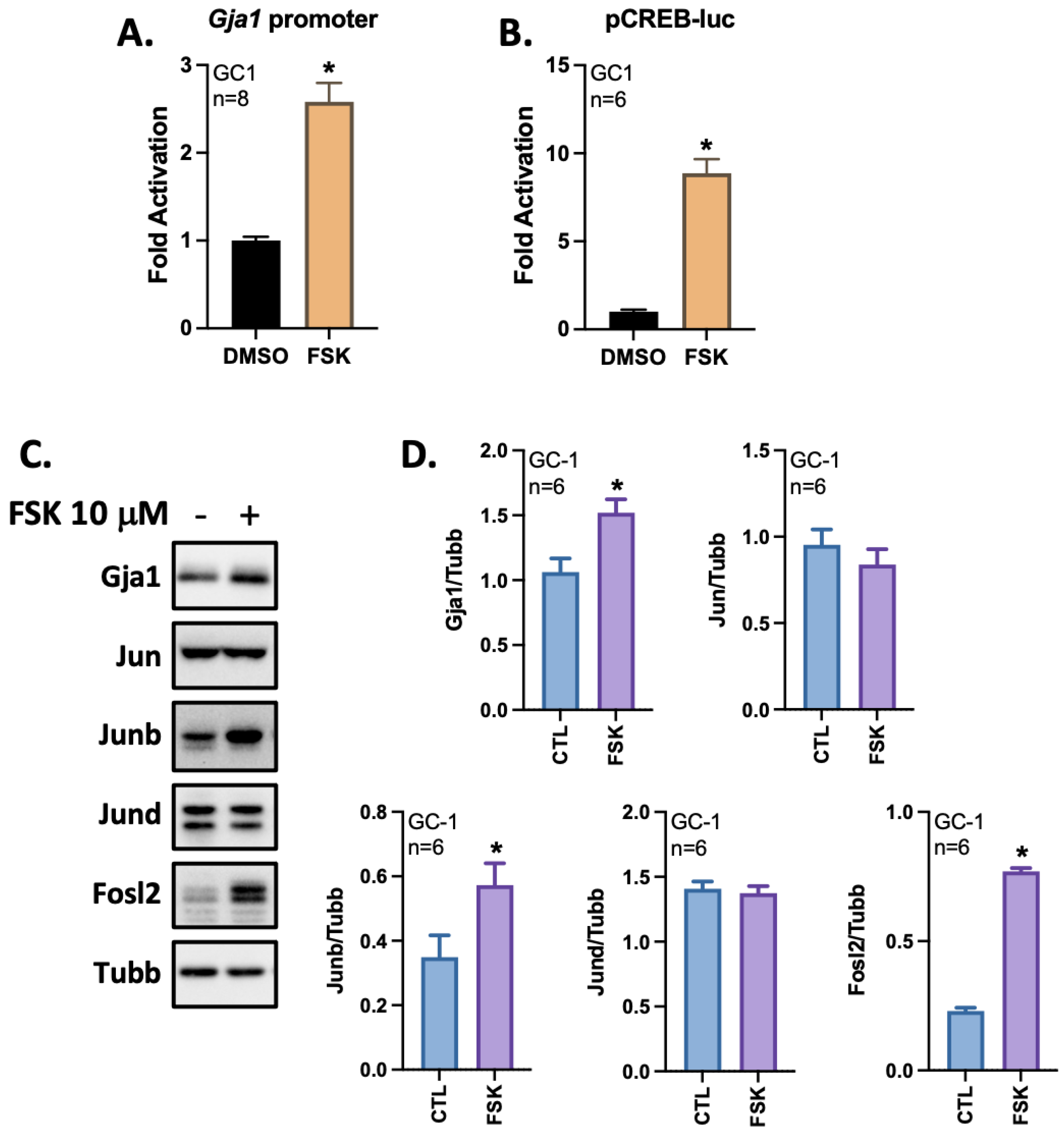
3.4. The cAMP/PKA Pathway Activates the Gja1 Promoter in GC-1 Cells
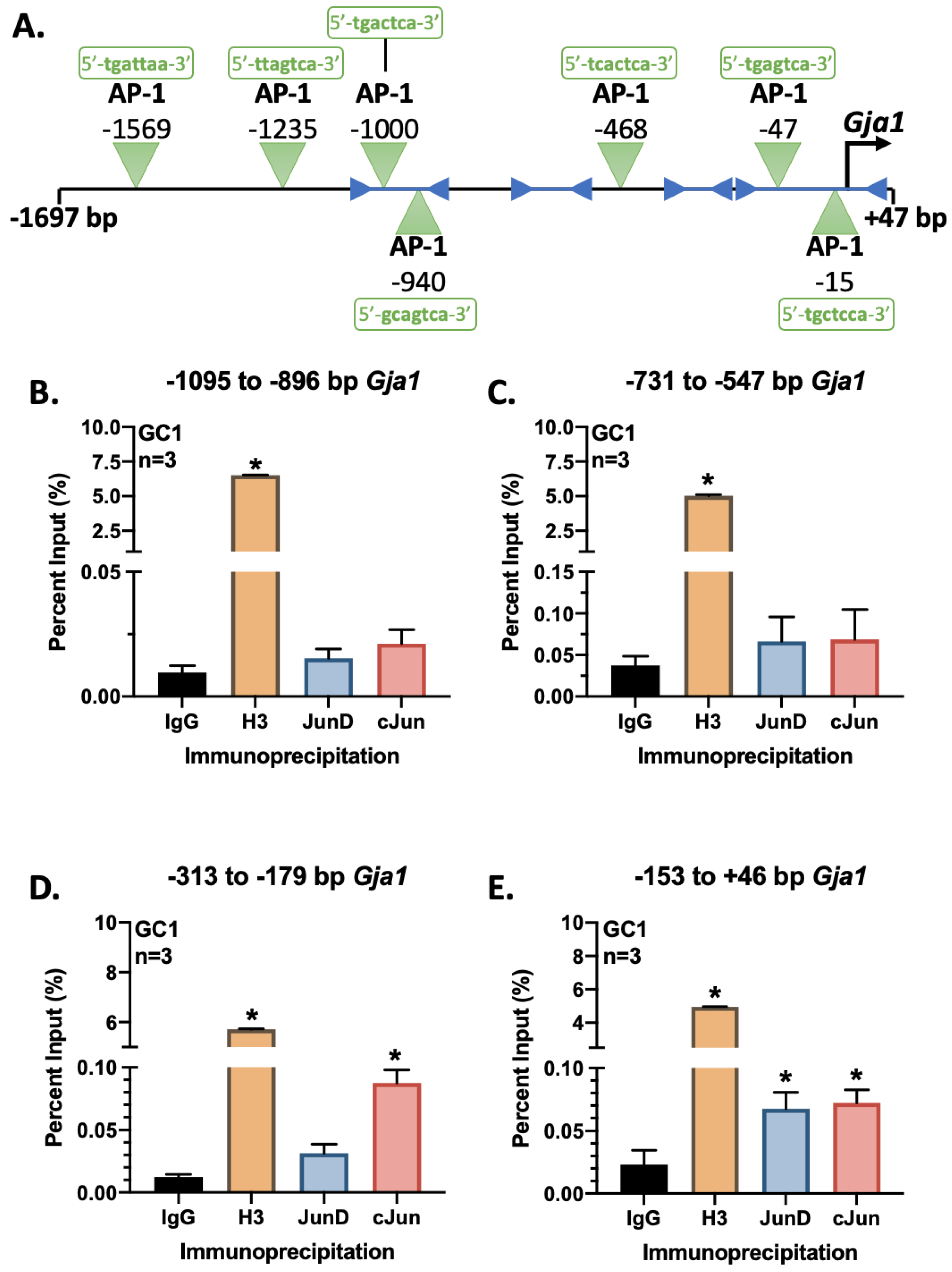
3.5. Jun Is Recruited to the Proximal Region of the Gja1 Promoter
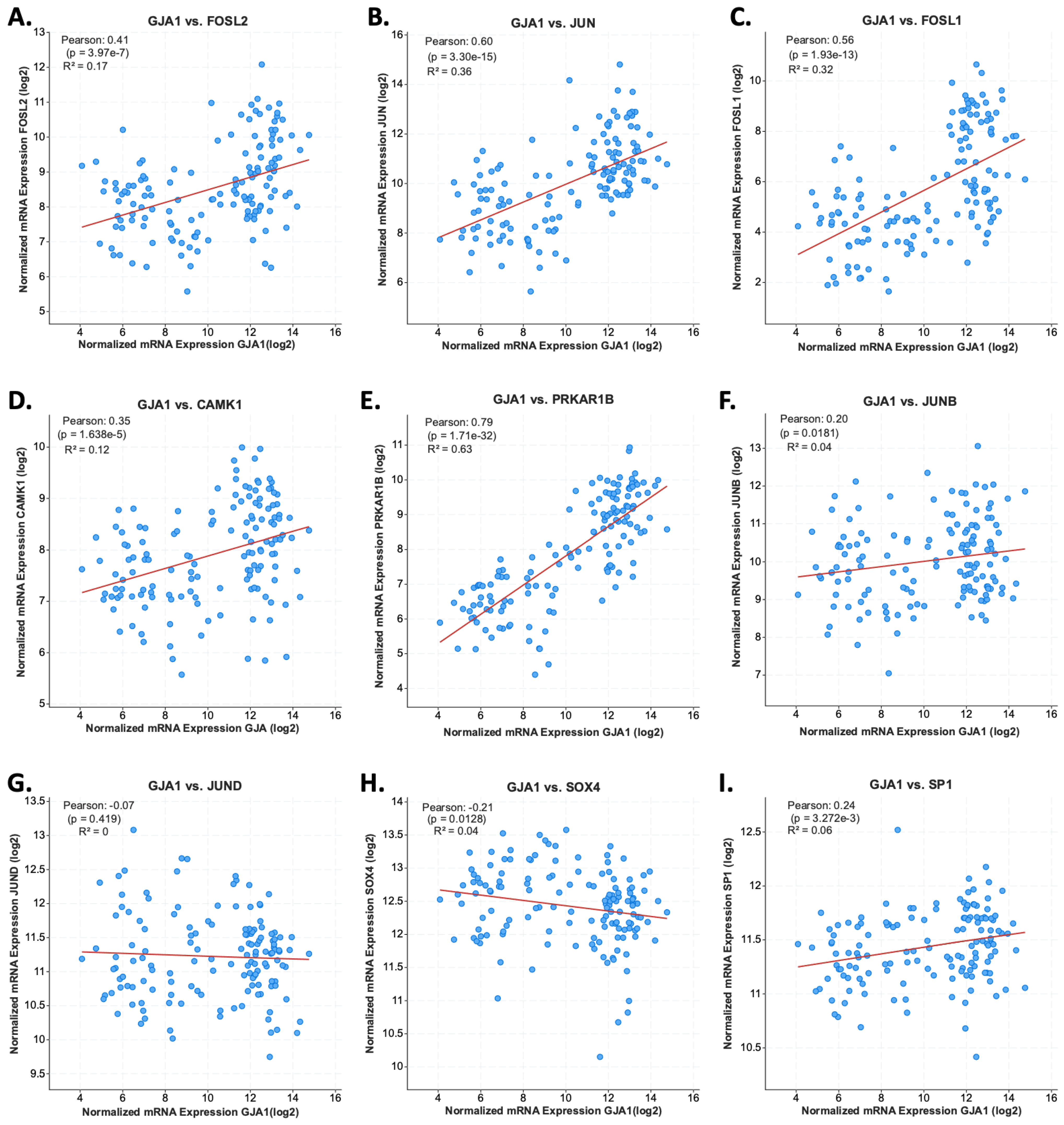
3.6. JUN and FOSL2 Are Correlated with GJA1 Expression in Testicular Germ Cells Tumors
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Risley, M.S.; Tan, I.P.; Roy, C.; Sáez, J.C. Cell-, Age- and Stage-Dependent Distribution of Connexin43 Gap Junctions in Testes. J. Cell. Sci. 1992, 103, 81–96. [Google Scholar] [CrossRef] [PubMed]
- Batias, C.; Defamie, N.; Lablack, A.; Thepot, D.; Fenichel, P.; Segretain, D.; Pointis, G. Modified Expression of Testicular Gap-Junction Connexin 43 during Normal Spermatogenic Cycle and in Altered Spermatogenesis. Cell Tissue Res. 1999, 298, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Juneja, S.C.; Barr, K.J.; Enders, G.C.; Kidder, G.M. Defects in the Germ Line and Gonads of Mice Lacking Connexin43. Biol. Reprod. 1999, 60, 1263–1270. [Google Scholar] [CrossRef] [PubMed]
- Brehm, R.; Zeiler, M.; Rüttinger, C.; Herde, K.; Kibschull, M.; Winterhager, E.; Willecke, K.; Guillou, F.; Lécureuil, C.; Steger, K.; et al. A Sertoli Cell-Specific Knockout of Connexin43 Prevents Initiation of Spermatogenesis. Am. J. Pathol. 2007, 171, 19–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sridharan, S.; Simon, L.; Meling, D.D.; Cyr, D.G.; Gutstein, D.E.; Fishman, G.I.; Guillou, F.; Cooke, P.S. Proliferation of Adult Sertoli Cells Following Conditional Knockout of the Gap Junctional Protein GJA1 (Connexin 43) in Mice. Biol. Reprod. 2007, 76, 804–812. [Google Scholar] [CrossRef] [Green Version]
- Carette, D.; Weider, K.; Gilleron, J.; Giese, S.; Dompierre, J.; Bergmann, M.; Brehm, R.; Denizot, J.-P.; Segretain, D.; Pointis, G. Major Involvement of Connexin 43 in Seminiferous Epithelial Junction Dynamics and Male Fertility. Dev. Biol. 2010, 346, 54–67. [Google Scholar] [CrossRef]
- Noelke, J.; Wistuba, J.; Damm, O.S.; Fietz, D.; Gerber, J.; Gaehle, M.; Brehm, R. A Sertoli Cell-Specific Connexin43 Knockout Leads to Altered Interstitial Connexin Expression and Increased Leydig Cell Numbers. Cell Tissue Res. 2015, 361, 633–644. [Google Scholar] [CrossRef]
- Günther, S.; Fietz, D.; Weider, K.; Bergmann, M.; Brehm, R. Effects of a Murine Germ Cell-Specific Knockout of Connexin 43 on Connexin Expression in Testis and Fertility. Transgenic Res. 2013, 22, 631–641. [Google Scholar] [CrossRef]
- Saez, J.C.; Berthoud, V.M.; Branes, M.C.; Martinez, A.D.; Beyer, E.C. Plasma Membrane Channels Formed by Connexins: Their Regulation and Functions. Physiol. Rev. 2003, 83, 1359–1400. [Google Scholar] [CrossRef] [Green Version]
- Roger, C.; Mograbi, B.; Chevallier, D.; Michiels, J.F.; Tanaka, H.; Segretain, D.; Pointis, G.; Fenichel, P. Disrupted Traffic of Connexin 43 in Human Testicular Seminoma Cells: Overexpression of Cx43 Induces Membrane Location and Cell Proliferation Decrease. J. Pathol. 2004, 202, 241–246. [Google Scholar] [CrossRef]
- Defamie, N.; Berthaut, I.; Mograbi, B.; Chevallier, D.; Dadoune, J.-P.; Fénichel, P.; Segretain, D.; Pointis, G. Impaired Gap Junction Connexin43 in Sertoli Cells of Patients with Secretory Azoospermia: A Marker of Undifferentiated Sertoli Cells. Lab. Investig. 2003, 83, 449–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brehm, R.; Rüttinger, C.; Fischer, P.; Gashaw, I.; Winterhager, E.; Kliesch, S.; Bohle, R.M.; Steger, K.; Bergmann, M. Transition from Preinvasive Carcinoma in Situ to Seminoma Is Accompanied by a Reduction of Connexin 43 Expression in Sertoli Cells and Germ Cells. Neoplasia 2006, 8, 499–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghouili, F.; Martin, L.J. Cooperative Regulation of Gja1 Expression by Members of the AP-1 Family CJun and CFos in TM3 Leydig and TM4 Sertoli Cells. Gene 2017, 635, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Ghouili, F.; Roumaud, P.; Martin, L.J. Gja1 Expression Is Regulated by Cooperation between SOX8/SOX9 and CJUN Transcription Factors in TM4 and 15P-1 Sertoli Cell Lines. Mol. Reprod. Dev. 2018, 85, 875–886. [Google Scholar] [CrossRef]
- Tratner, I.; Ofir, R.; Verma, I.M. Alteration of a Cyclic AMP-Dependent Protein Kinase Phosphorylation Site in the c-Fos Protein Augments Its Transforming Potential. Mol. Cell. Biol. 1992, 12, 998–1006. [Google Scholar] [CrossRef] [Green Version]
- Teyssier, C.; Belguise, K.; Galtier, F.; Chalbos, D. Characterization of the Physical Interaction between Estrogen Receptor Alpha and JUN Proteins. J. Biol. Chem. 2001, 276, 36361–36369. [Google Scholar] [CrossRef] [Green Version]
- Wayman, G.A.; Kaech, S.; Grant, W.F.; Davare, M.; Impey, S.; Tokumitsu, H.; Nozaki, N.; Banker, G.; Soderling, T.R. Regulation of Axonal Extension and Growth Cone Motility by Calmodulin-Dependent Protein Kinase I. J. Neurosci. 2004, 24, 3786–3794. [Google Scholar] [CrossRef]
- Tremblay, J.J.; Hamel, F.; Viger, R.S. Protein Kinase A-Dependent Cooperation between GATA and CCAAT/Enhancer-Binding Protein Transcription Factors Regulates Steroidogenic Acute Regulatory Protein Promoter Activity. Endocrinology 2002, 143, 3935–3945. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, M.C.; Narisawa, S.; Hess, R.A.; Millán, J.L. Immortalization of Germ Cells and Somatic Testicular Cells Using the SV40 Large T Antigen. Exp. Cell Res. 1992, 201, 417–435. [Google Scholar] [CrossRef]
- Boussif, O.; Lezoualc’h, F.; Zanta, M.A.; Mergny, M.D.; Scherman, D.; Demeneix, B.; Behr, J.P. A Versatile Vector for Gene and Oligonucleotide Transfer into Cells in Culture and in Vivo: Polyethylenimine. Proc. Natl. Acad. Sci. USA 1995, 92, 7297–7301. [Google Scholar] [CrossRef] [Green Version]
- Zagore, L.L.; Sweet, T.J.; Hannigan, M.M.; Weyn-Vanhentenryck, S.M.; Jobava, R.; Hatzoglou, M.; Zhang, C.; Licatalosi, D.D. DAZL Regulates Germ Cell Survival through a Network of PolyA-Proximal MRNA Interactions. Cell Rep. 2018, 25, 1225–1240.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie Enables Improved Reconstruction of a Transcriptome from RNA-Seq Reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Gao, C.; Lin, X.; Ning, Y.; He, W.; Zheng, C.; Zhang, D.; Yan, L.; Jiang, B.; Zhao, Y.; et al. The MicroRNA MiR-202 Prevents Precocious Spermatogonial Differentiation and Meiotic Initiation during Mouse Spermatogenesis. Development 2021, 148, dev199799. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A Fast Spliced Aligner with Low Memory Requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Echetebu, C.O.; Ali, M.; Izban, M.G.; MacKay, L.; Garfield, R.E. Localization of Regulatory Protein Binding Sites in the Proximal Region of Human Myometrial Connexin 43 Gene. Mol. Hum. Reprod. 1999, 5, 757–766. [Google Scholar] [CrossRef] [Green Version]
- Geimonen, E.; Boylston, E.; Royek, A.; Andersen, J. Elevated Connexin-43 Expression in Term Human Myometrium Correlates with Elevated c-Jun Expression and Is Independent of Myometrial Estrogen Receptors. J. Clin. Endocrinol. Metab. 1998, 83, 1177–1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Groot, R.P.; Sassone-Corsi, P. Activation of Jun/AP-1 by Protein Kinase A. Oncogene 1992, 7, 2281–2286. [Google Scholar] [PubMed]
- Agustin, J.T.; Wilkerson, C.G.; Witman, G.B. The Unique Catalytic Subunit of Sperm CAMP-Dependent Protein Kinase Is the Product of an Alternative Calpha MRNA Expressed Specifically in Spermatogenic Cells. Mol. Biol. Cell 2000, 11, 3031–3044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The CBio Cancer Genomics Portal: An Open Platform for Exploring Multidimensional Cancer Genomics Data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative Analysis of Complex Cancer Genomics and Clinical Profiles Using the CBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [Green Version]
- Alcivar, A.A.; Hake, L.E.; Hardy, M.P.; Hecht, N.B. Increased Levels of JunB and C-Jun MRNAs in Male Germ Cells Following Testicular Cell Dissociation. Maximal Stimulation in Prepuberal Animals. J. Biol. Chem. 1990, 265, 20160–20165. [Google Scholar] [CrossRef]
- Geimonen, E.; Jiang, W.; Ali, M.; Fishman, G.I.; Garfield, R.E.; Andersen, J. Activation of Protein Kinase C in Human Uterine Smooth Muscle Induces Connexin-43 Gene Transcription through an AP-1 Site in the Promoter Sequence. J. Biol. Chem. 1996, 271, 23667–23674. [Google Scholar] [CrossRef] [Green Version]
- Hoadley, K.A.; Yau, C.; Hinoue, T.; Wolf, D.M.; Lazar, A.J.; Drill, E.; Shen, R.; Taylor, A.M.; Cherniack, A.D.; Thorsson, V.; et al. Cell-of-Origin Patterns Dominate the Molecular Classification of 10,000 Tumors from 33 Types of Cancer. Cell 2018, 173, 291–304.e6. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Xu, H.; Lin, S.; Deng, W.; Zhou, J.; Zhang, Y.; Shi, Y.; Peng, D.; Xue, Y. GPS 5.0: An Update on the Prediction of Kinase-Specific Phosphorylation Sites in Proteins. Genom. Proteom. Bioinform. 2020, 18, 72–80. [Google Scholar] [CrossRef]
- Sheng, M.; Thompson, M.A.; Greenberg, M.E. CREB: A Ca2+-Regulated Transcription Factor Phosphorylated by Calmodulin-Dependent Kinases. Science 1991, 252, 1427–1430. [Google Scholar] [CrossRef]
- Fang, W.-L.; Lai, S.-Y.; Lai, W.-A.; Lee, M.-T.; Liao, C.-F.; Ke, F.-C.; Hwang, J.-J. CRTC2 and Nedd4 Ligase Involvement in FSH and TGFβ1 Upregulation of Connexin43 Gap Junction. J. Mol. Endocrinol. 2015, 55, 263–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamph, W.W.; Dwarki, V.J.; Ofir, R.; Montminy, M.; Verma, I.M. Negative and Positive Regulation by Transcription Factor CAMP Response Element-Binding Protein Is Modulated by Phosphorylation. Proc. Natl. Acad. Sci. USA 1990, 87, 4320–4324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitt, J.M.; Wayman, G.A.; Nozaki, N.; Soderling, T.R. Calcium Activation of ERK Mediated by Calmodulin Kinase I. J. Biol. Chem. 2004, 279, 24064–24072. [Google Scholar] [CrossRef] [Green Version]
- Monje, P.; Hernández-Losa, J.; Lyons, R.J.; Castellone, M.D.; Gutkind, J.S. Regulation of the Transcriptional Activity of C-Fos by ERK. A Novel Role for the Prolyl Isomerase PIN1. J. Biol. Chem. 2005, 280, 35081–35084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, I.; Hughes, R.T.; Mayer, B.J.; Yee, K.; Woodgett, J.R.; Avruch, J.; Kyriakis, J.M.; Zon, L.I. Role of SAPK/ERK Kinase-1 in the Stress-Activated Pathway Regulating Transcription Factor c-Jun. Nature 1994, 372, 794–798. [Google Scholar] [CrossRef]
- Chalmers, C.J.; Gilley, R.; March, H.N.; Balmanno, K.; Cook, S.J. The Duration of ERK1/2 Activity Determines the Activation of c-Fos and Fra-1 and the Composition and Quantitative Transcriptional Output of AP-1. Cell Signal. 2007, 19, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Karin, M. The Regulation of AP-1 Activity by Mitogen-Activated Protein Kinases. J. Biol. Chem. 1995, 270, 16483–16486. [Google Scholar] [CrossRef] [Green Version]
- Yanagibashi, K.; Papadopoulos, V.; Masaki, E.; Iwaki, T.; Kawamura, M.; Hall, P.F. Forskolin Activates Voltage-Dependent Ca2+ Channels in Bovine but Not in Rat Fasciculata Cells. Endocrinology 1989, 124, 2383–2391. [Google Scholar] [CrossRef]
- Morita, K.; Dohi, T.; Kitayama, S.; Koyama, Y.; Tsujimoto, A. Stimulation-Evoked Ca2+ Fluxes in Cultured Bovine Adrenal Chromaffin Cells Are Enhanced by Forskolin. J. Neurochem. 1987, 48, 248–252. [Google Scholar] [CrossRef]
- Carystinos, G.D.; Kandouz, M.; Alaoui-Jamali, M.A.; Batist, G. Unexpected Induction of the Human Connexin 43 Promoter by the Ras Signaling Pathway Is Mediated by a Novel Putative Promoter Sequence. Mol. Pharmacol. 2003, 63, 821–831. [Google Scholar] [CrossRef] [Green Version]
- Winterhager, E.; Pielensticker, N.; Freyer, J.; Ghanem, A.; Schrickel, J.W.; Kim, J.-S.; Behr, R.; Grümmer, R.; Maass, K.; Urschel, S.; et al. Replacement of Connexin43 by Connexin26 in Transgenic Mice Leads to Dysfunctional Reproductive Organs and Slowed Ventricular Conduction in the Heart. BMC Dev. Biol. 2007, 7, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Risley, M.S. Connexin Gene Expression in Seminiferous Tubules of the Sprague-Dawley Rat. Biol. Reprod. 2000, 62, 748–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldberg, G.S.; Valiunas, V.; Brink, P.R. Selective Permeability of Gap Junction Channels. Biochim. Biophys. Acta 2004, 1662, 96–101. [Google Scholar] [CrossRef] [Green Version]
- Decrouy, X.; Gasc, J.-M.; Pointis, G.; Segretain, D. Functional Characterization of Cx43 Based Gap Junctions during Spermatogenesis. J. Cell. Physiol. 2004, 200, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Risley, M.S.; Tan, I.P.; Farrell, J. Gap Junctions with Varied Permeability Properties Establish Cell-Type Specific Communication Pathways in the Rat Seminiferous Epithelium. Biol. Reprod. 2002, 67, 945–952. [Google Scholar] [CrossRef] [Green Version]
- Rode, K.; Langeheine, M.; Seeger, B.; Brehm, R. Connexin43 in Germ Cells Seems to Be Dispensable for Murine Spermatogenesis. Int. J. Mol. Sci. 2021, 22, 7924. [Google Scholar] [CrossRef]
- Kidder, G.M.; Cyr, D.G. Roles of Connexins in Testis Development and Spermatogenesis. Semin. Cell Dev. Biol. 2016, 50, 22–30. [Google Scholar] [CrossRef]
- Chevallier, D.; Carette, D.; Segretain, D.; Gilleron, J.; Pointis, G. Connexin 43 a Check-Point Component of Cell Proliferation Implicated in a Wide Range of Human Testis Diseases. Cell. Mol. Life Sci. 2013, 70, 1207–1220. [Google Scholar] [CrossRef]
- Yamasaki, H.; Omori, Y.; Krutovskikh, V.; Zhu, W.; Mironov, N.; Yamakage, K.; Mesnil, M. Connexins in Tumour Suppression and Cancer Therapy. Novartis Found. Symp. 1999, 219, 241–254; discussion 254–260. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Najih, M.; Basque, A.; Nguyen, H.T.; Diawara, M.; Martin, L.J. Members of the AP-1 Family of Transcription Factors Regulate the Expression of Gja1 in Mouse GC-1 Spermatogonial Cells. Appl. Sci. 2022, 12, 1408. https://0-doi-org.brum.beds.ac.uk/10.3390/app12031408
Najih M, Basque A, Nguyen HT, Diawara M, Martin LJ. Members of the AP-1 Family of Transcription Factors Regulate the Expression of Gja1 in Mouse GC-1 Spermatogonial Cells. Applied Sciences. 2022; 12(3):1408. https://0-doi-org.brum.beds.ac.uk/10.3390/app12031408
Chicago/Turabian StyleNajih, Mustapha, Audrey Basque, Ha Tuyen Nguyen, Mariama Diawara, and Luc J. Martin. 2022. "Members of the AP-1 Family of Transcription Factors Regulate the Expression of Gja1 in Mouse GC-1 Spermatogonial Cells" Applied Sciences 12, no. 3: 1408. https://0-doi-org.brum.beds.ac.uk/10.3390/app12031408