Comparison of Petroleum Hydrocarbons Degradation by Klebsiella pneumoniae and Pseudomonas aeruginosa

 , , , ,
, , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Chemicals and Culture Mediums
2.3. Petroleum Hydrocarbons for Degradation
2.4. Surface Tension Test
2.5. Bacterial Biomass and PHs Degradation Efficacy Test
2.6. Hydrophobicity and Emulsification Test
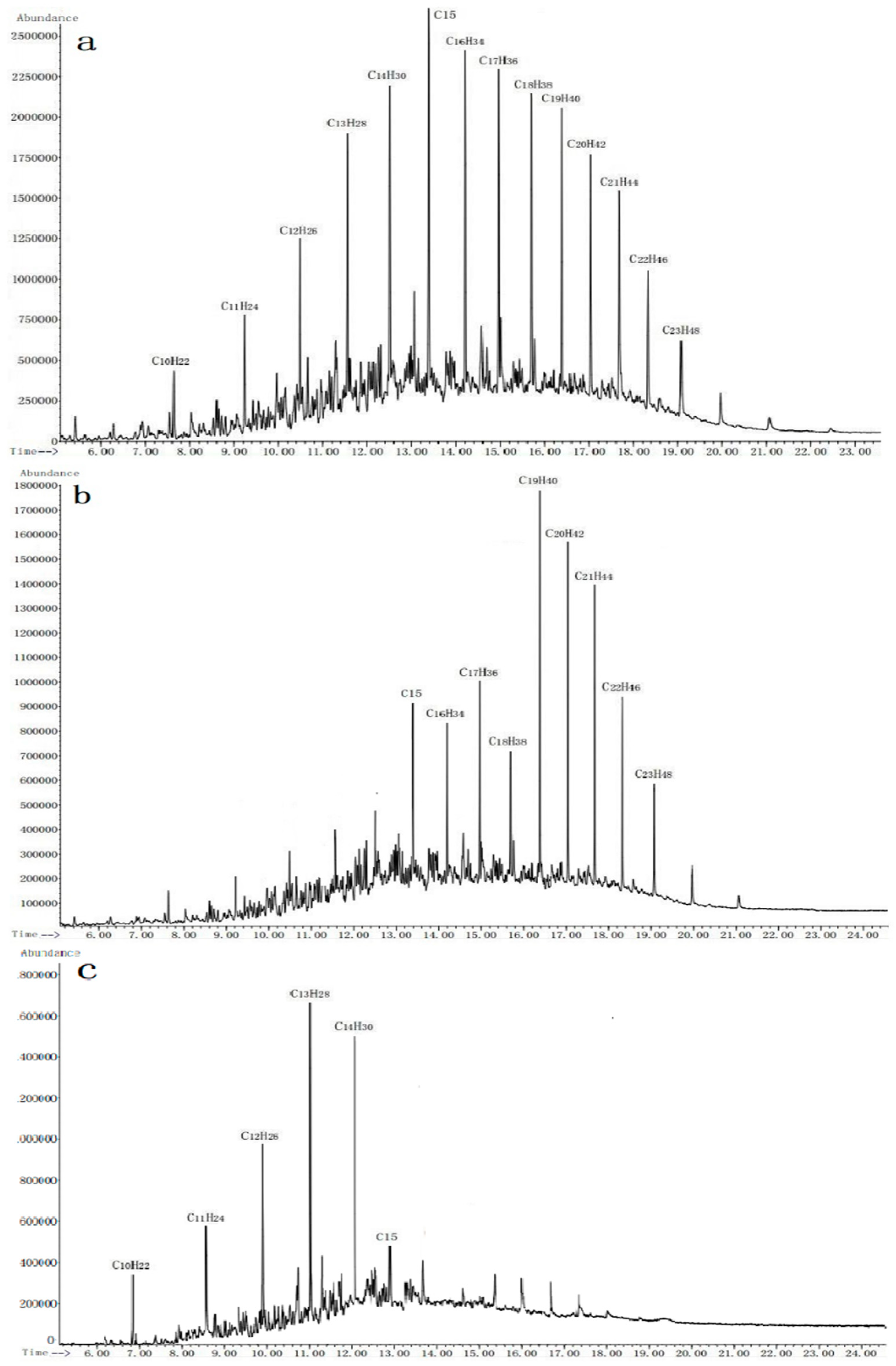
2.7. GC-MS Analysis
2.8. FTIR and HPLC-MS Analyses
2.9. Statistics
3. Results
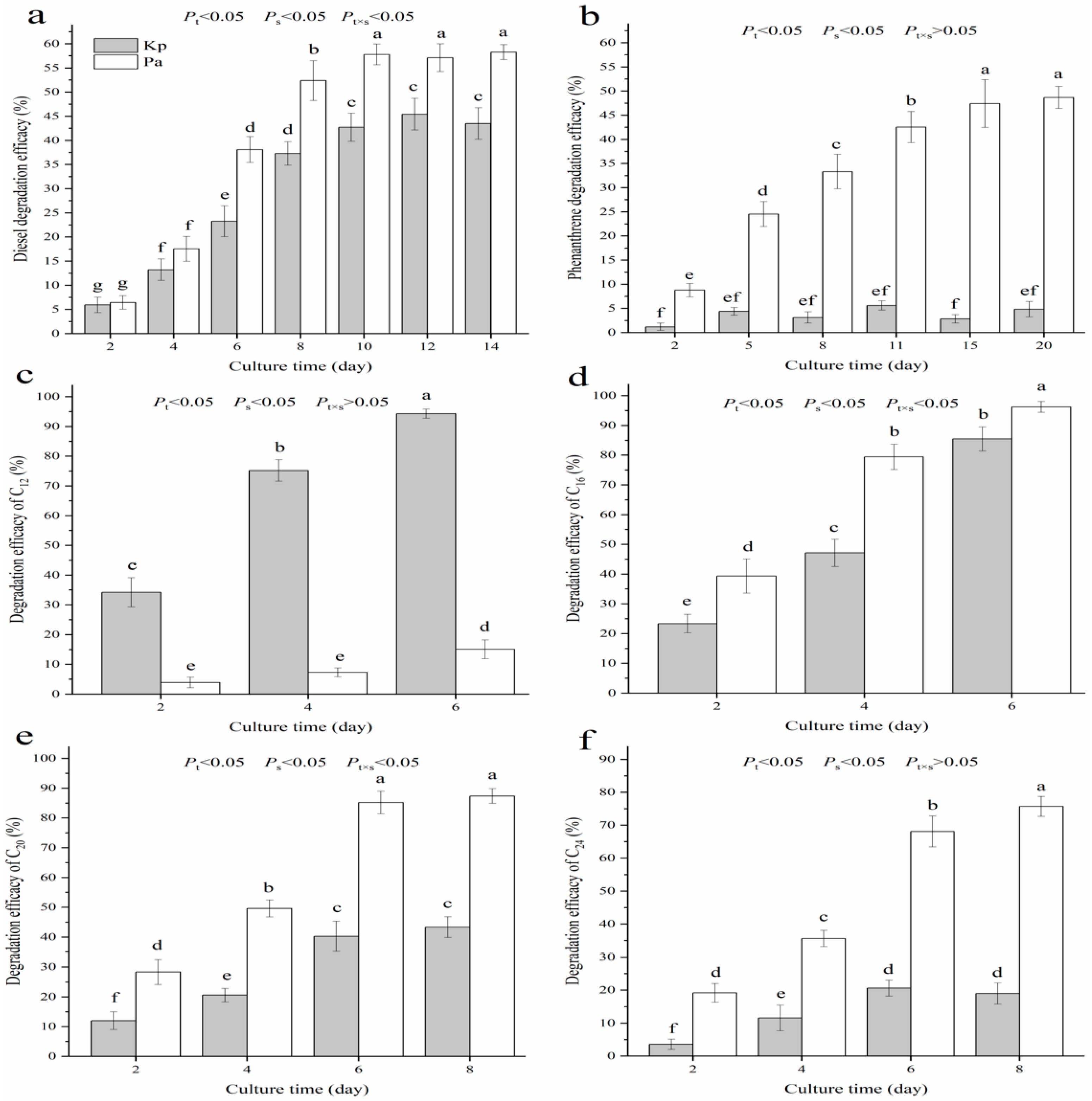
3.1. Biodegradability of Diesel and Its Main Constituents
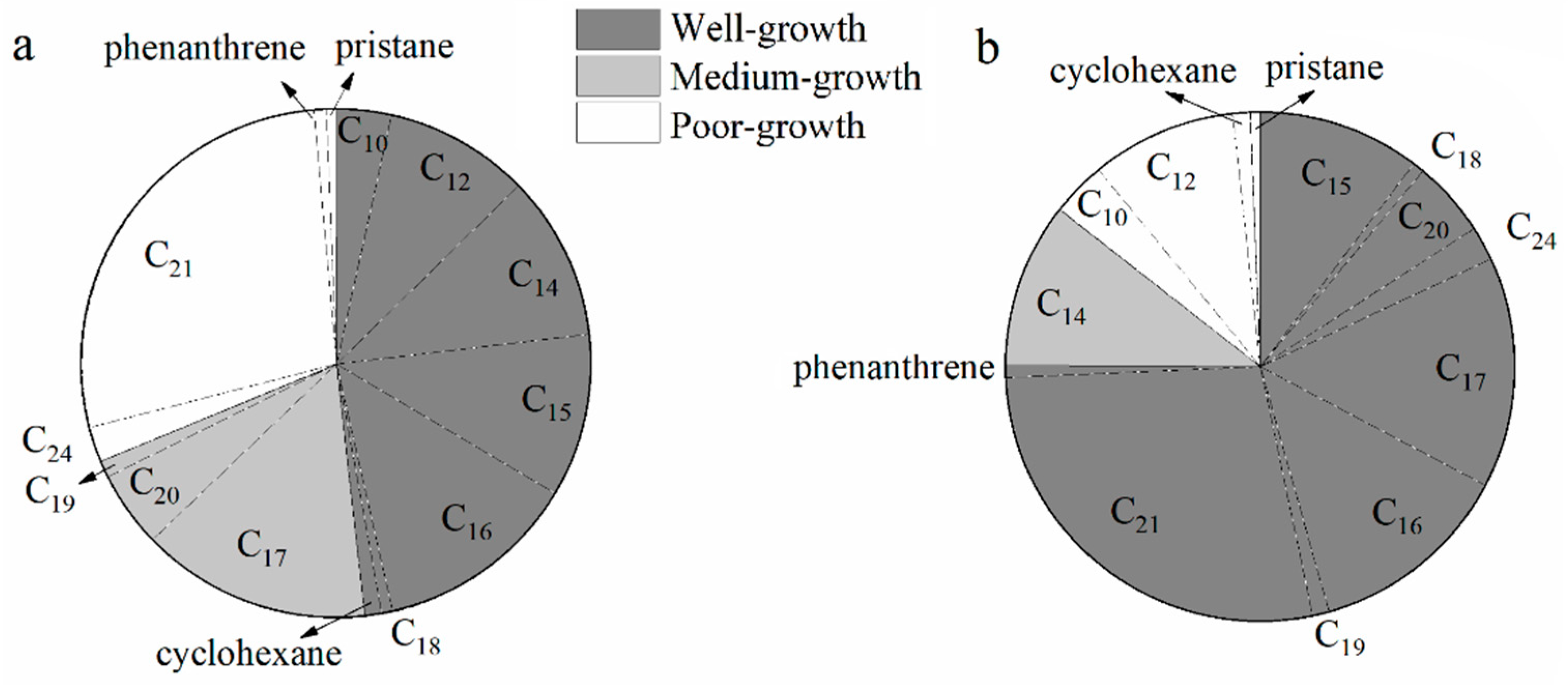
3.2. Biodegrading Range of Diesel Constituents
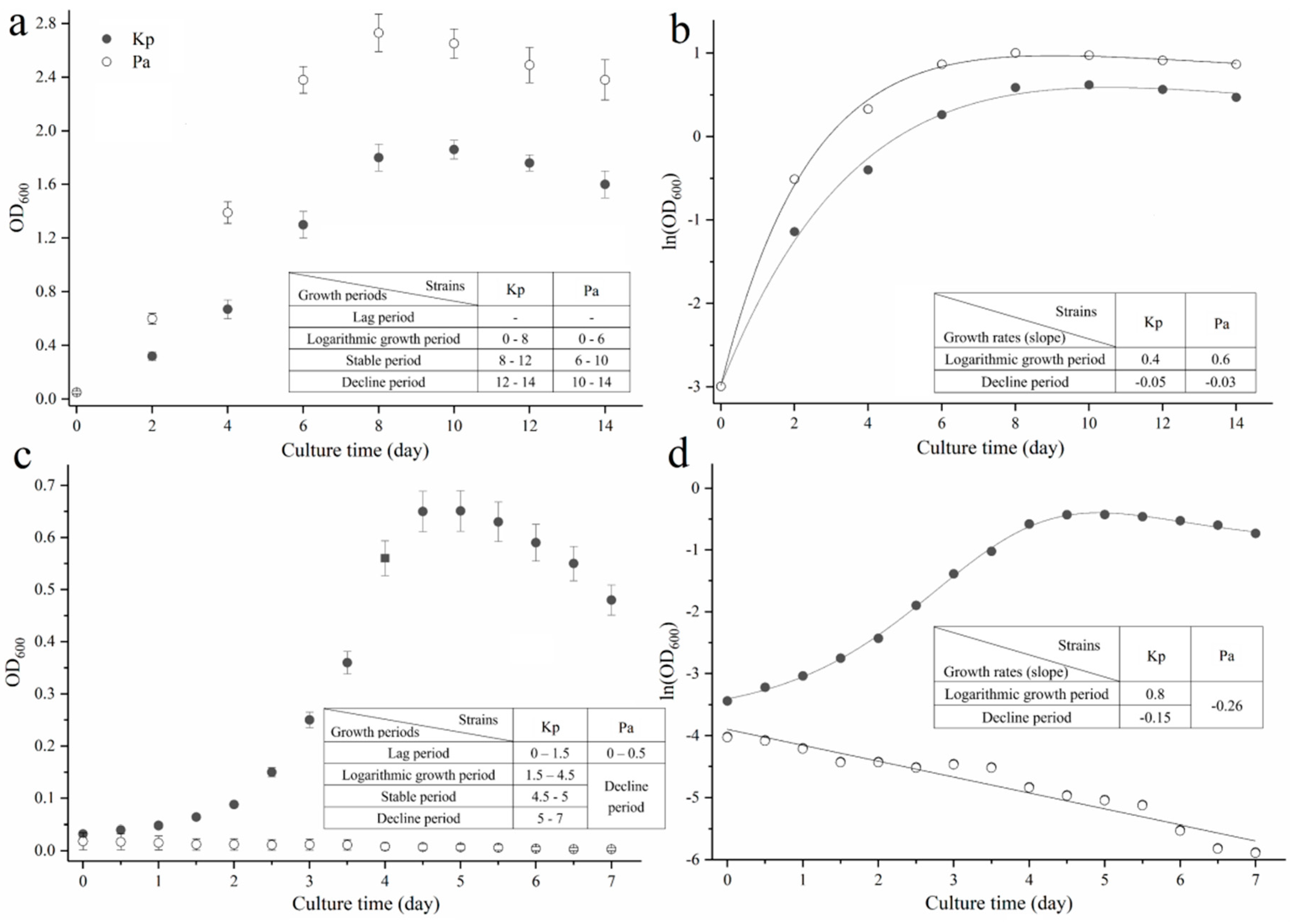
3.3. Growth Characteristics of Kp and Pa in Media with Diesel or Its Constituents as Sole Carbon Source
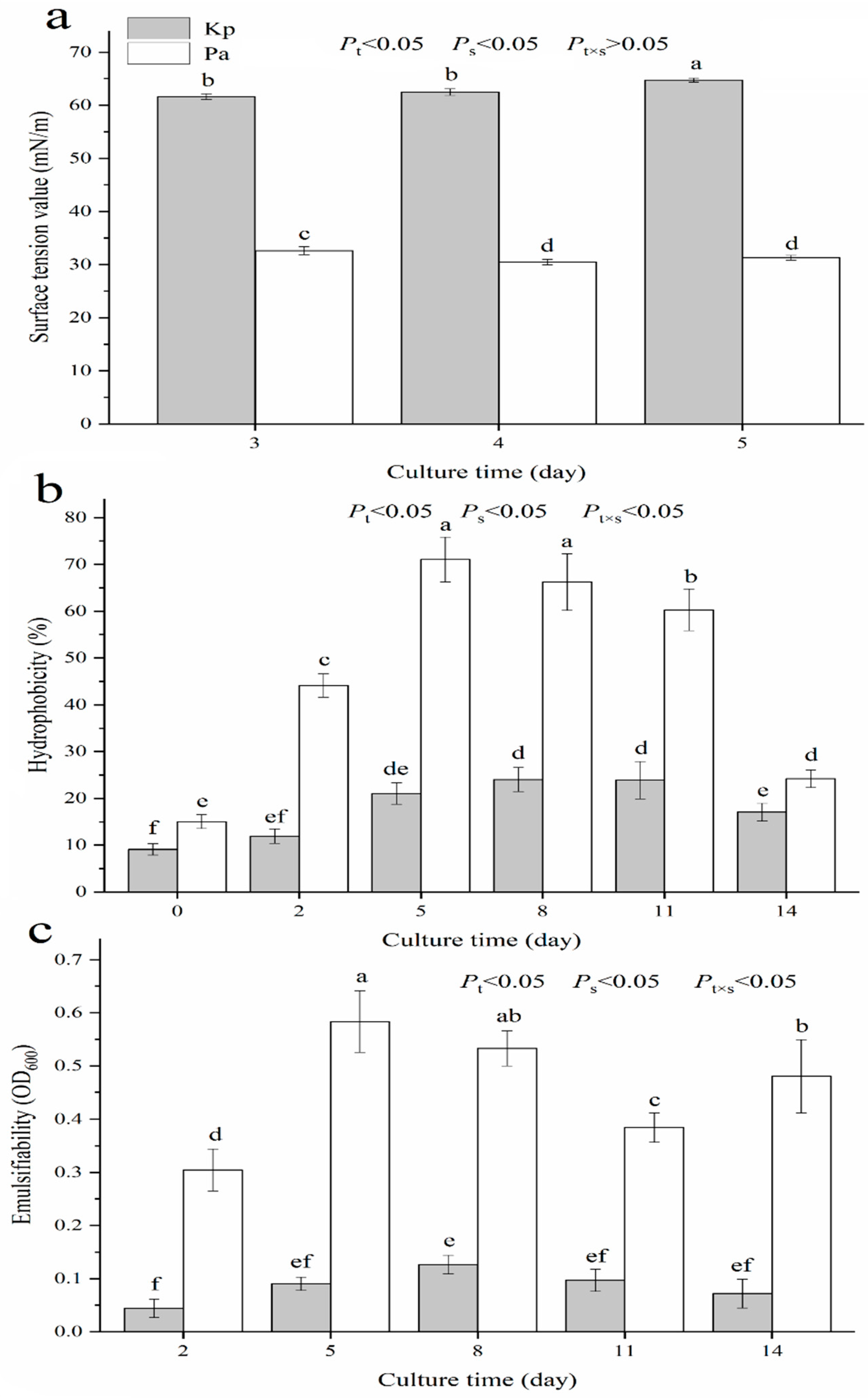
3.4. Surface Tension, Hydrophobicity and Emulsifiability
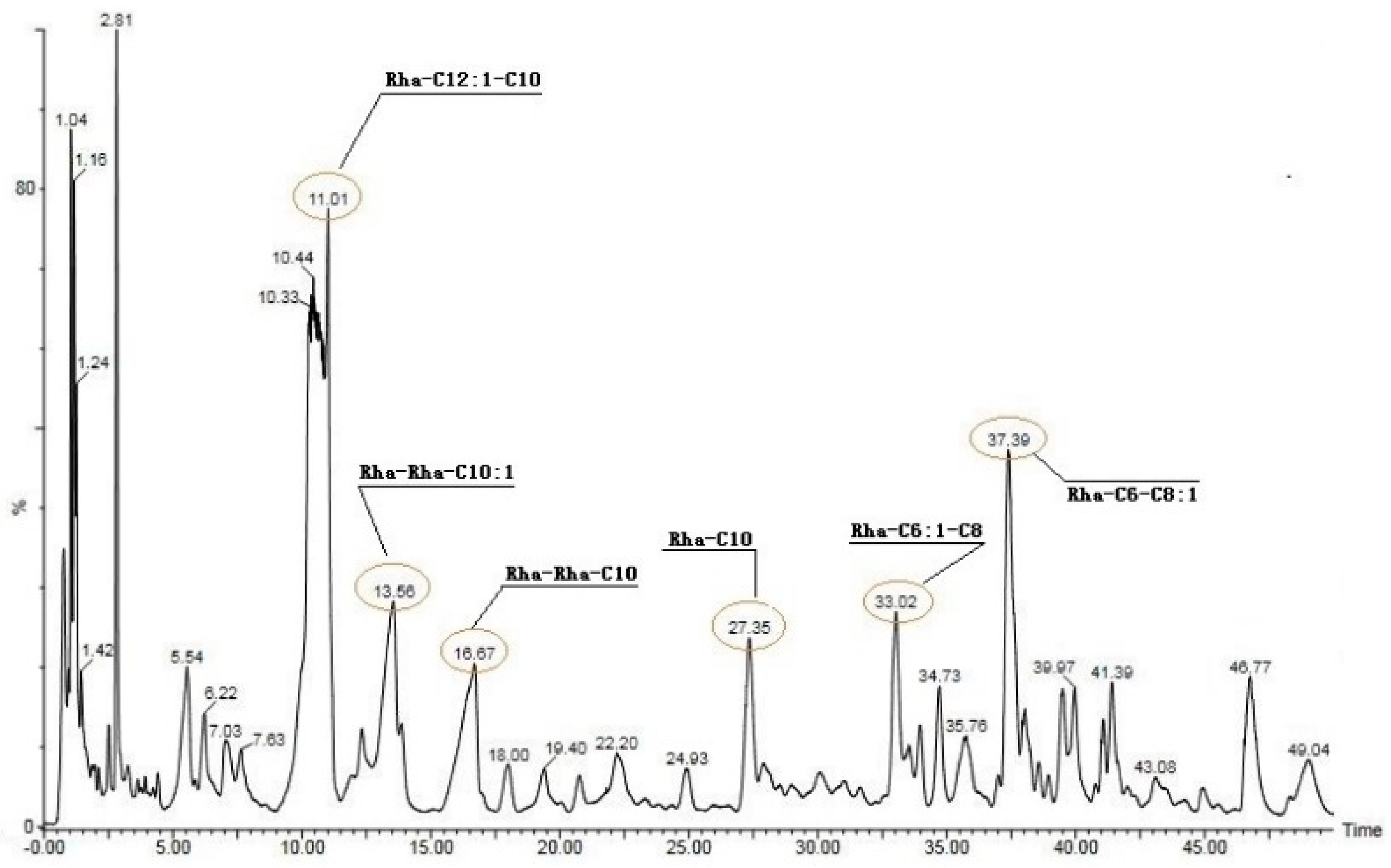
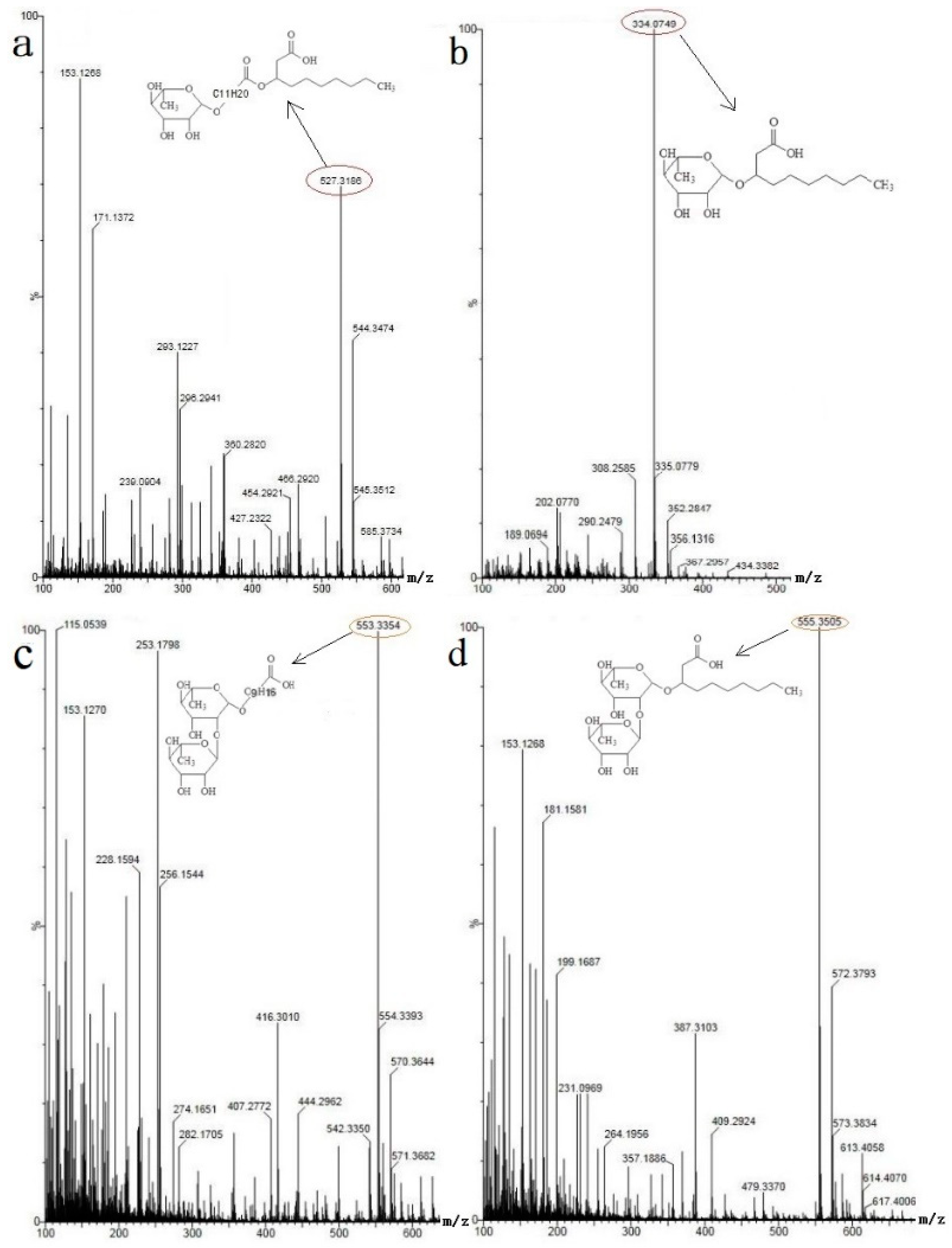
3.5. Group Characterization of Bacterial Secretions
3.6. Potential Factors Related to Degradation Efficacy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
References
- Nakazawa, M.M.; Gavazza, S.; Kato, M.T.; Florencio, L. Evaluation of rhamnolipid addition on the natural attenuation of estuarine sediments contaminated with diesel oil. Environ. Sci. Pollut. R. 2017, 24, 25522–25533. [Google Scholar] [CrossRef]
- Shpiner, R.; Liu, G.; Stuckey, D. Treatment of oilfield produced water by waste stabilization ponds: Biodegradation of petroleum-derived materials. Bioresour. Technol. 2009, 100, 6229–6235. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; Pi, Y.; Bao, M.; Xu, N.; Li, Y.; Lu, J. Biodegradation of different petroleum hydrocarbons by free and immobilized microbial consortia. Environ. Sci. Proc. Impact. 2015, 17, 2022–2033. [Google Scholar] [CrossRef] [PubMed]
- Atlas, R.M.; Hazen, T.C. Oil Biodegradation and Bioremediation: A Tale of the Two Worst Spills in US History. Environ. Sci. Technol. 2011, 45, 6709–6715. [Google Scholar] [CrossRef] [PubMed]
- Pinedo, J.; Ibanez, R.; Lijzen, J.P.A.; Irabien, A. Assessment of soil pollution based on total petroleum hydrocarbons and individual oil substances. J. Environ. Manag. 2013, 130, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.W.; Zheng, X.L.; Li, L.L.; Sun, J. Adsorption of aqueous oil on sands and its studies of effective factors. Environ. Sci. 2006, 27, 2019. [Google Scholar]
- Wang, Z.Q.; Wu, Q.; Zou, Z.G.; Chen, H.; Yang, X.C.; Zhao, J.C. Study on the groundwater petroleum contaminant remediation by air sparging. Environ. Sci. 2007, 28, 754. [Google Scholar]
- Ugochukwu, U.C.; Ochonogor, A. Groundwater contamination by polycyclic aromatic hydrocarbon due to diesel spill from a telecom base station in a Nigerian City: Assessment of human health risk exposure. Environ. Monit. Assess. 2018, 190, 249. [Google Scholar] [CrossRef]
- Cheng, Q.; Ge, W.; Chai, C.; Wu, J.; Ma, D.; Zeng, L.; Zhu, X.; Chen, Q.; Li, J. Polycyclic Aromatic Hydrocarbons in Soil Around Coal-Fired Power Plants in Shandong, China. Pol. J. Environ. Stud. 2019, 28, 53–65. [Google Scholar] [CrossRef]
- Wojtera-Kwiczor, J.; Zukowska, W.; Graj, W.; Malecka, A.; Piechalak, A.; Ciszewska, L.; Chrzanowski, L.; Lisiecki, P.; Komorowicz, I.; Baralkiewicz, D.; et al. Rhizoremediation of diesel-contaminated soil with two rapeseed varieties and petroleum degraders reveals different responses of the plant defense mechanisms. Int. J. Phytoremediat. 2014, 16, 770–789. [Google Scholar] [CrossRef]
- Polyak, Y.M.; Bakina, L.G.; Chugunova, M.V.; Mayachkina, N.V.; Gerasimov, A.O.; Bure, V.M. Effect of remediation strategies on biological activity of oil-contaminated soil—A field study. Int. Biodeterior. Biodegrad. 2018, 126, 57–68. [Google Scholar] [CrossRef]
- Safdari, M.S.; Kariminia, H.R.; Nejad, Z.G.; Fletcher, T.H. Study Potential of Indigenous Pseudomonas aeruginosa and Bacillus subtilis in Bioremediation of Diesel-Contaminated Water. Water Air Soil Poll. 2017, 228, 1–7. [Google Scholar] [CrossRef]
- Vazquez, S.; Monien, P.; Minetti, R.P.; Jurgens, J.; Curtosi, A.; Primitz, J.V.; Frickenhaus, S.; Abele, D.; Mac Cormack, W.; Helmke, E. Bacterial communities and chemical parameters in soils and coastal sediments in response to diesel spills at Carlini Station, Antarctica. Sci. Total. Environ. 2017, 605, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Nie, M.Q.; Wan, Y.; Tian, X.T.; Nie, H.Y.; Zi, J.; Ma, X. Functional characterization of two alkane hydroxylases in a versatile Pseudomonas aeruginosa strain NY3. Ann. Microbiol. 2017, 67, 459–468. [Google Scholar] [CrossRef]
- Rashidi, H.R.; Sulaiman, N.M.N.; Hashim, N.A.; Hassan, C.R.C.; Emami, S.D. Simulated textile (batik) wastewater pre-treatment through application of a baffle separation tank. Desalin. Water. Treat. 2016, 57, 151–160. [Google Scholar] [CrossRef]
- Sang, Y.M.; Gu, Q.B.; Sun, T.C.; Li, F.S. Color and organic compounds removal from secondary effluent of landfill leachate with a novel inorganic polymer coagulant. Water. Sci. Technol. 2008, 58, 1423–1432. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, T.V.C.; Cardoso, S.L.; Vieira, R.B.; Vieira, P.A.; Cardoso, V.L. Biodegradation of effluent contaminated with diesel oil and gasoline using chitosan as a natural coagulant in a continuous process. Braz. J. Chem. Eng. 2016, 4, 863. [Google Scholar] [CrossRef]
- Mailler, R.; Gasperi, J.; Rocher, V.; Gilbert-Pawlik, S.; Geara-Matta, D.; Moilleron, R.; Chebbo, G. Biofiltration vs conventional activated sludge plants: what about priority and emerging pollutants removal? Environ. Sci. Pollut. Res. 2014, 21, 5379–5390. [Google Scholar] [CrossRef]
- Gill, R.T.; Harbottle, M.J.; Smith, J.W.; Thornton, S.F. Electrokinetic-enhanced bioremediation of organic contaminants: A review of processes and environmental applications. Chemosphere 2014, 107, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Kadri, T.; Rouissi, T.; Kaur Brar, S.; Cledon, M.; Sarma, S.; Verma, M. Biodegradation of polycyclic aromatic hydrocarbons (PAHs) by fungal enzymes: A review. J. Environ. Sci. China 2017, 51, 52–74. [Google Scholar] [CrossRef]
- Kachieng‘a, L.; Momba, M.N.B. Kinetics of petroleum oil biodegradation by a consortium of three protozoan isolates (Aspidisca sp., Trachelophyllum sp. and Peranema sp.). Biotechnol. Rep. (Amst.) 2017, 15, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Tso, S.F.; Taghon, G.L. Protozoan grazing increases mineralization of naphthalene in marine sediment. Microb. Ecol. 2006, 51, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, N.; Shahi, S.K. Degradation of polycyclic aromatic hydrocarbon (pyrene) using novel fungal strain Coriolopsis byrsina strain APC5. Int. Biodeter. Biodegr. 2017, 122, 69–81. [Google Scholar] [CrossRef]
- Moubasher, A.H.; Abdel-Sater, M.A.; Soliman, Z.S.M. Yeasts and filamentous fungi associated with some dairy products in Egypt. J. Mycol. Med. 2018, 28, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Acevedo-Sandoval, O.; Gutierrez-Alcantara, E.J.; Perez-Balan, R.; Rodriguez-Vazquez, G.; Zamorategui-Molina, A.; Tirado-Torres, D. Degradation Of Polycyclic Aromatic Hydrocarbons Using Bacterial Isolate From the Contaminated Soil And White Rot Fungus Pleurotus Ostreatus. Appl. Ecol. Environ. Res. 2018, 16, 3815–3829. [Google Scholar] [CrossRef]
- Shaeyan, M.; Tirandaz, H.; Ghanbarpour, S.; Seyedipour, N.; Shavandi, M.; Dastgheib, S.M.M. Bioremediation of a drilling waste-contaminated soil; biotreatability assessment and microcosm optimization for developing a field-scale remediation process. Iran. J. Biotechnol. 2018, 16, 193–199. [Google Scholar] [CrossRef]
- Al-Hawash, A.B.; Li, S.; Alhujaily, A.; Zhang, X.; Ma, F.; Dragh, M.A.; Abbood, H.A. Principles of microbial degradation of petroleum hydrocarbons in the environment. Egypt. J. Aquat. Res. 2018, 44, 71–76. [Google Scholar] [CrossRef]
- Zhang, Z.Z.; Gai, L.X.; Hou, Z.W.; Yang, C.Y.; Ma, C.Q.; Wang, Z.G.; Sun, B.P.; He, X.F.; Tang, H.Z.; Xu, P. Characterization and biotechnological potential of petroleum-degrading bacteria isolated from oil-contaminated soils. Bioresour. Technol. 2010, 101, 8452–8456. [Google Scholar] [CrossRef]
- You-Qing, L.; Hong-Fang, L.; Zhen-Le, T.; Li-Hua, Z.; Ying-Hui, W.; He-Qing, T. Diesel pollution biodegradation: Synergetic effect of Mycobacterium and filamentous fungi. Biomed. Environ. Sci. 2008, 21, 181–187. [Google Scholar]
- Xue, J.; Yu, Y.; Bai, Y.; Wang, L.; Wu, Y. Marine Oil-Degrading Microorganisms and Biodegradation Process of Petroleum Hydrocarbon in Marine Environments: A Review. Curr. Microbiol. 2015, 71, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Wei, G.U.O.; Mengchang, H.E.; Zhifeng, Y. A Review of Studies on the Degradation of Petroleum Hydrocarbon in Soils and Sediments by Microorganism. Bull. Mineral. Petrol. Geochem. 2007, 26, 276–283. [Google Scholar]
- Montagnolli, R.N.; Lopes, P.R.; Bidoia, E.D. Assessing Bacillus subtilis biosurfactant effects on the biodegradation of petroleum products. Environ. Monit. Assess. 2015, 187, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.X.; Li, Z.; Ji, Q.N.; Wang, S.W.; Su, D.G.; Lin, Y.S. Effect of thermal oxidation of activated carbon surface on its adsorption towards dibenzothiophene. Chem. Eng. J. 2009, 148, 242–247. [Google Scholar] [CrossRef]
- Adam, G.; Duncan, H.J. Effect of diesel fuel on growth of selected plant species. Environ. Geochem. Health 1999, 21, 353–357. [Google Scholar] [CrossRef]
- Sugiura, K.; Ishihara, M.; Toshitsugu Shimauchi, A.; Harayama, S. Physicochemical Properties and Biodegradability of Crude Oil. Environ. Sci. Technol. 1997, 31, 45–51. [Google Scholar] [CrossRef]
- Takei, D.; Washio, K.; Morikawa, M. Identification of alkane hydroxylase genes in Rhodococcus sp. strain TMP2 that degrades a branched alkane. Biotechnol. Lett. 2008, 30, 1447. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tang, J.; Wang, L.; Liu, J.; Gurav, R.G.; Sun, K. A novel bioremediation strategy for petroleum hydrocarbon pollutants using salt tolerant Corynebacterium variabile HRJ4 and biochar. J. Environ. Sci. China 2016, 47, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Ulrici, W. Contaminant Soil Areas, Different Countries and Contaminant Monitoring of Contaminants, Biotechnology: Second, Completely Revised Edition; Wiley Online Library: Hoboken, NJ, USA, 2008; pp. 11–12. [Google Scholar]
- Ron, E.Z.; Rosenberg, E. Biosurfactants and oil bioremediation. Curr. Opin. Biotechnol. 2002, 13, 249–252. [Google Scholar] [CrossRef]
- Morán, A.C.; Olivera, N.; Commendatore, M.; Esteves, J.L.; Siñeriz, F. Enhancement of hydrocarbon wastebiodegradation by addition of a biosurfactantfrom Bacillus subtilis O9. Biodegradation 2000, 11, 65–71. [Google Scholar] [CrossRef]
- Tsomides, H.J.; Hughes, J.B.; Thomas, J.M.; Ward, C.H. Effect of surfactant addition on phenanthrene biodegradation in sediments. Environ. Toxicol. Chem. 1995, 14, 953–959. [Google Scholar] [CrossRef] [Green Version]
- Chrzanowski, Ł.; Czaczyk, K. Why do microorganisms produce rhamnolipids? World J. Microbiol. Biotechnol. 2012, 28, 401–419. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Leon, V.; Materano, A.S.; Ilzins, O.A. Enhancement of oil degradation by co-culture of hydrocarbon degrading and biosurfactant producing bacteria. Pol. J. Microbiol. 2006, 55, 139. [Google Scholar] [PubMed]
- Lu, J.R.; Zhao, X.B.; Yaseen, M. Biomimetic amphiphiles: Biosurfactants. Curr. Opin. Colloid In. 2007, 12, 60–67. [Google Scholar] [CrossRef]
- Lindman, B.; Wennerström, H. Miceles. Amphiphile aggregation in aqueous solution. Top. Curr. Chem. 1980, 87, 1–83. [Google Scholar] [PubMed]
- Owsianiak, M.; Szulc, A.; Chrzanowski, Ł.; Cyplik, P.; Bogacki, M.; Olejnikschmidt, A.K.; Heipieper, H.J. Biodegradation and surfactant-mediated biodegradation of diesel fuel by 218 microbial consortia are not correlated to cell surface hydrophobicity. Appl. Microbiol. Biotechnol. 2009, 84, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Yao, J.; Liu, R.; Zhu, M.; Wang, F.; Wu, X.; Liu, H. Effect of natural and synthetic surfactants on crude oil biodegradation by indigenous strains. Ecotoxicol. Environ. Saf. 2016, 129, 171. [Google Scholar] [CrossRef] [PubMed]
- Muthukamalam, S.; Sivagangavathi, S.; Dhrishya, D.; Sudha, S.R. Characterization of dioxygenases and biosurfactants produced by crude oil degrading soil bacteria. Braz. J. Microbiol. 2017, 48, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Reiling, H.E.; Thanei-Wyss, U.; Guerra-Santos, L.H.; Hirt, R.; Käppeli, O.; Fiechter, A. Pilot plant production of rhamnolipid biosurfactant by Pseudomonas aeruginosa. Appl. Environ. Microbiol. 1986, 51, 985–989. [Google Scholar]
- Xu, J.; Deng, H.; Huang, T.; Song, S. Enhanced biodegradation of crude oil in contaminated soil by inoculation of hydrocarbon-degraders. Desalin. Water Treat. 2014, 52, 5126–5135. [Google Scholar] [CrossRef]
- Van Hamme, J.D.; Singh, A.; Ward, O.P. Recent advances in petroleum microbiology. Microbiol. Mol. Biol. Rev. 2003, 67, 503–549. [Google Scholar] [CrossRef]
- Pi, Y.; Meng, L.; Bao, M.; Sun, P.; Lu, J. Degradation of crude oil and relationship with bacteria and enzymatic activities in laboratory testing. Int. Biodeterior. Biodegrad. 2016, 106, 106–116. [Google Scholar] [CrossRef]
- Fuchs, G.; Boll, M.; Heider, J. Microbial degradation of aromatic compounds—From one strategy to four. Nat. Rev. Microbiol. 2011, 9, 803–816. [Google Scholar] [CrossRef] [PubMed]
- Kronenberg, M.; Trably, E.; Bernet, N.; Patureau, D. Biodegradation of polycyclic aromatic hydrocarbons: Using microbial bioelectrochemical systems to overcome an impasse. Environ. Pollut. 2017, 231, 509–523. [Google Scholar] [CrossRef] [PubMed]
- Ostendorf, D.W.; Schoenberg, T.H.; Hinlein, E.S.; Long, S.C. Monod kinetics for aerobic biodegradation of petroleum hydrocarbons in unsaturated soil microcosms. Environ. Sci. Technol. 2007, 41, 2343–2349. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-Y.; Xu, Y.; Wang, H.-Y.; Zhao, J.; Gao, D.-M.; Li, F.-M.; Xing, B. Biodegradation of Crude Oil in Contaminated Soils by Free and Immobilized Microorganisms. Pedosphere 2012, 22, 717–725. [Google Scholar] [CrossRef]
- Xu, X.; You, Z.; Zhang, S.; Tang, Y.; Li, Z.; Xu, H. Synergistic mechanisms of degradation of diesel oil by bacterial consortium. Environ. Sci. Technol. 2018, 41, 47–55. [Google Scholar]
- Patowary, K.; Saikia, R.R.; Kalita, M.C.; Deka, S. Degradation of polyaromatic hydrocarbons employing biosurfactant-producing Bacillus pumilus KS2. Ann. Microbiol. 2015, 65, 225–234. [Google Scholar] [CrossRef]
- Wang, L.; Tang, Y.; Wang, S.; Liu, R.L.; Liu, M.Z.; Zhang, Y.; Liang, F.L.; Feng, L. Isolation and characterization of a novel thermophilic Bacillus strain degrading long-chain n-alkanes. Extremophiles 2006, 10, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Bouchez-Naitali, M.; Rakatozafy, H.; Marchal, R.; Leveau, J.Y.; Vandecasteele, J.P. Diversity of bacterial strains degrading hexadecane in relation to the mode of substrate uptake. J. Appl. Microbiol. 1999, 86, 421–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzintzun-Camacho, O.; Loera, O.; Ramirez-Saad, H.C.; Gutierrez-Rojas, M. Comparison of mechanisms of hexadecane uptake among pure and mixed cultures derived from a bacterial consortium. Int. Biodeterior. Biodegrad. 2012, 70, 1–7. [Google Scholar] [CrossRef]
- Mariaamalraj, S.K.; Pasumarthi, R.; Achary, A.; Mutnuri, S. Effect of rhamnolipid on biodegradation of hydrocarbons in non-aqueous-phase liquid (NAPL). Bioremed. J. 2016, 20, 183–193. [Google Scholar] [CrossRef]
- Coreteam, R. R: A language and environment for statistical computing. Computing 2015, 1, 12–21. [Google Scholar]
- Perni, S.; Andrew, P.W.; Shama, G. Estimating the maximum growth rate from microbial growth curves: definition is everything. Food Microbiol. 2005, 22, 491–495. [Google Scholar] [CrossRef] [Green Version]
- Wei, S. Electrospray ionization mass spectrometric detection of rhamnolipids and their acid precursors in Pseudomonas sp. BS-03 cultures. Prog. Biotechnol. 2005, 25, 83–87. [Google Scholar]
- Habib, S.; Ahmad, S.A.; Johari, W.L.W.; Abd Shukor, M.Y.; Alias, S.A.; Khalil, K.A.; Yasid, N.A. Evaluation of conventional and response surface level optimisation of n-dodecane (n-C12) mineralisation by psychrotolerant strains isolated from pristine soil at Southern Victoria Island, Antarctica. Microb. Cell Fact. 2018, 17, 44–65. [Google Scholar] [CrossRef]
- Almailem, D.M.; Sorkhoh, N.A.; Alawadhi, H.; Eliyas, M.; Radwan, S.S. Biodegradation of crude oil and pure hydrocarbons by extreme halophilic archaea from hypersaline coasts of the Arabian Gulf. Extremophiles 2010, 14, 321–328. [Google Scholar] [CrossRef]
- Ebadi, A.; Olamaee, M.; Sima, N.A.K.; Nasrabadi, R.G.; Hashemi, M. Isolation and Characterization of Biosurfactant Producing and Crude Oil Degrading Bacteria from Oil Contaminated Soils. Iran. J. Sci. Technol. A 2018, 42, 1149–1156. [Google Scholar] [CrossRef]
- Wu, T.; Xu, J.; Xie, W.; Yao, Z.; Yang, H.; Sun, C.; Li, X. Pseudomonas aeruginosa L10: A Hydrocarbon-Degrading, Biosurfactant-Producing, and Plant-Growth-Promoting Endophytic Bacterium Isolated From a Reed (Phragmites australis). Front. Microbiol. 2018, 9, 1087–1099. [Google Scholar] [CrossRef]
- Sharma, A.; Rehman, M.B. Laboratory scale bioremediation of diesel hydrocarbon in soil by indigenous bacterial consortium. Indian J. Exp. Biol. 2009, 47, 766–769. [Google Scholar]
- Wang, X.B.; Chi, C.Q.; Nie, Y.; Tang, Y.Q.; Tan, Y.; Wu, G.; Wu, X.L. Degradation of petroleum hydrocarbons (C6-C40) and crude oil by a novel Dietzia strain. Bioresour. Technol. 2011, 102, 7755–7761. [Google Scholar] [CrossRef]
- Gojgic-Cvijovic, G.D.; Milic, J.S.; Solevic, T.M.; Beskoski, V.P.; Ilic, M.V.; Djokic, L.S.; Narancic, T.M.; Vrvic, M.M. Biodegradation of petroleum sludge and petroleum polluted soil by a bacterial consortium: A laboratory study. Biodegradation 2012, 23, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lingyi, M. Screening, Purification and Analysis of a Strain Producted Glycolipid Surfactant. Ph.D. Thesis, Northeast Normal University, Changchun, China, 2015. [Google Scholar]
- Reddy, P.G.; Singh, H.D.; Pathak, M.G.; Bhagat, S.D.; Baruah, J.N. Isolation And Functional-Characterization Of Hydrocarbon Emulsifying And Solubilizing Factors Produced by a Pseudomonas Species. Biotechnol Bioeng. 1983, 25, 387–401. [Google Scholar] [CrossRef] [PubMed]
- Qiao, N.; Shao, Z. Isolation and characterization of a novel biosurfactant produced by hydrocarbon-degrading bacterium Alcanivorax dieselolei B-5. J. Appl. Microbiol. 2010, 108, 1207–1216. [Google Scholar] [CrossRef] [Green Version]
- Mani, P.; Dineshkumar, G.; Jayaseelan, T.; Deepalakshmi, K.; Kumar, C.G.; Balan, S.S. Antimicrobial activities of a promising glycolipid biosurfactant from a novel marine Staphylococcussaprophyticus SBPS 15. 3Biotech 2016, 6, 163–172. [Google Scholar]
- Batista, S.B.; Mounteer, A.H.; Amorim, F.R.; Tótola, M.R. Isolation and characterization of biosurfactant/bioemulsifier-producing bacteria from petroleum contaminated sites. Bioresour. Technol. 2006, 97, 868–875. [Google Scholar] [CrossRef] [PubMed]
- Chrzanowski, L.; Wick, L.Y.; Meulenkamp, R.; Kaestner, M.; Heipieper, H.J. Rhamnolipid biosurfactants decrease the toxicity of chlorinated phenols to Pseudomonas putida DOT-T1E. Lett. Appl. Microbiol. 2010, 48, 756–762. [Google Scholar]
- Chrzanowski, L.; Owsianiak, M.; Szulc, A.; Marecik, R.; Piotrowska-Cyplik, A.; Olejnik-Schmidt, A.K.; Staniewski, J.; Lisiecki, P.; Ciesielczyk, F.; Jesionowski, T. Interactions between rhamnolipid biosurfactants and toxic chlorinated phenols enhance biodegradation of a model hydrocarbon-rich effluent. Int. Biodeterior. Biodegrad. 2011, 65, 605–611. [Google Scholar] [CrossRef]
- Kaczorek, E.; Chrzanowski, L.; Pijanowska, A.; Olszanowski, A. Yeast and bacteria cell hydrophobicity and hydrocarbon biodegradation in the presence of natural surfactants: Rhamnolipides and saponins. Bioresour. Technol. 2008, 99, 4285–4291. [Google Scholar] [CrossRef] [PubMed]
- Łukasz, C.; Dziadas, M.; Łukasz, Ł.; Cyplik, P.; Białas, W.; Szulc, A.; Lisiecki, P.; Jeleń, H. Biodegradation of rhamnolipids in liquid cultures: Effect of biosurfactant dissipation on diesel fuel/B20 blend biodegradation efficiency and bacterial community composition. Bioresour. Technol. 2012, 111, 328–335. [Google Scholar]
- Sydow, M.; Owsianiak, M.; Szczepaniak, Z.; Framski, G.; Smets, B.F.; Łukasz, Ł.; Lisiecki, P.; Szulc, A.; Cyplik, P.; Łukasz, C. Evaluating robustness of a diesel-degrading bacterial consortium isolated from contaminated soil. New Biotechnol. 2016, 33, 852–859. [Google Scholar] [CrossRef] [Green Version]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
You, Z.; Xu, H.; Zhang, S.; Kim, H.; Chiang, P.-C.; Yun, W.; Zhang, L.; He, M. Comparison of Petroleum Hydrocarbons Degradation by Klebsiella pneumoniae and Pseudomonas aeruginosa. Appl. Sci. 2018, 8, 2551. https://0-doi-org.brum.beds.ac.uk/10.3390/app8122551
You Z, Xu H, Zhang S, Kim H, Chiang P-C, Yun W, Zhang L, He M. Comparison of Petroleum Hydrocarbons Degradation by Klebsiella pneumoniae and Pseudomonas aeruginosa. Applied Sciences. 2018; 8(12):2551. https://0-doi-org.brum.beds.ac.uk/10.3390/app8122551
Chicago/Turabian StyleYou, Zhaoyang, Haiyang Xu, Shujuan Zhang, Hyunook Kim, Pen-Chi Chiang, Wenfei Yun, Li Zhang, and Miao He. 2018. "Comparison of Petroleum Hydrocarbons Degradation by Klebsiella pneumoniae and Pseudomonas aeruginosa" Applied Sciences 8, no. 12: 2551. https://0-doi-org.brum.beds.ac.uk/10.3390/app8122551