
Salvianolic Acid C Protects against Cisplatin-Induced Acute Kidney Injury through Attenuation of Inflammation, Oxidative Stress and Apoptotic Effects and Activation of the CaMKK–AMPK–Sirt1-Associated Signaling Pathway in Mouse Models

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Animals
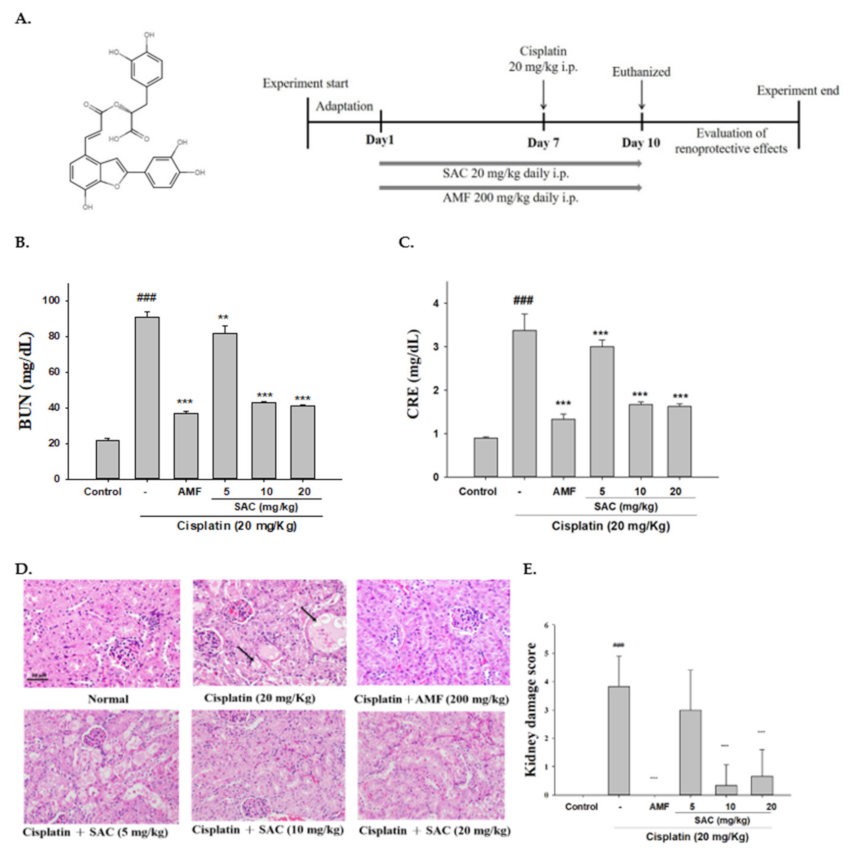
2.3. Research Design
2.4. Assess Kidney/Body Mass Index
2.5. Renal Function Tests
2.6. Histopathological Analysis
2.7. TUNEL Staining
2.8. Lipid Peroxidation Assays
2.9. Cytokine Assay
2.10. Nitrite Assay
2.11. Glutathione Estimation
2.12. Western Blot Analysis
2.13. Statistical Analysis
3. Results
3.1. SAC Inhibits Renal Damage and Improves Renal Function in Mice with Cisplatin-Induced Renal Injury
3.2. Changes in the Renal Index of SAC-Protected Mice Treated with Cisplatin
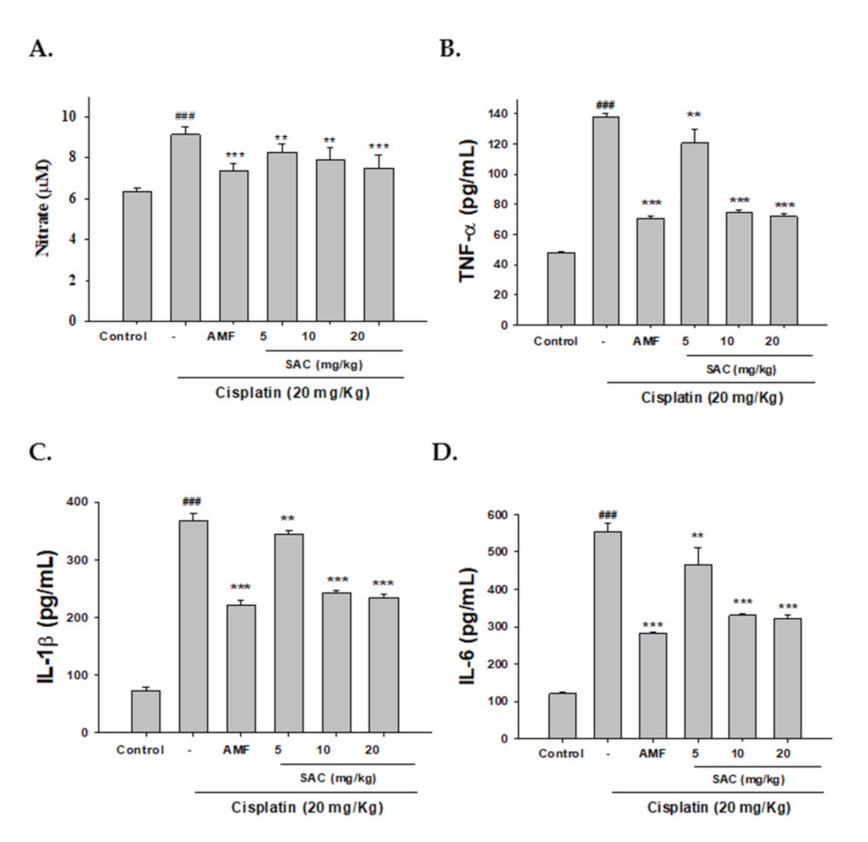
3.3. SAC Decreases NO and Pro-Inflammatory Cytokine Serum Levels in Cisplatin-Associated Nephrotoxicity
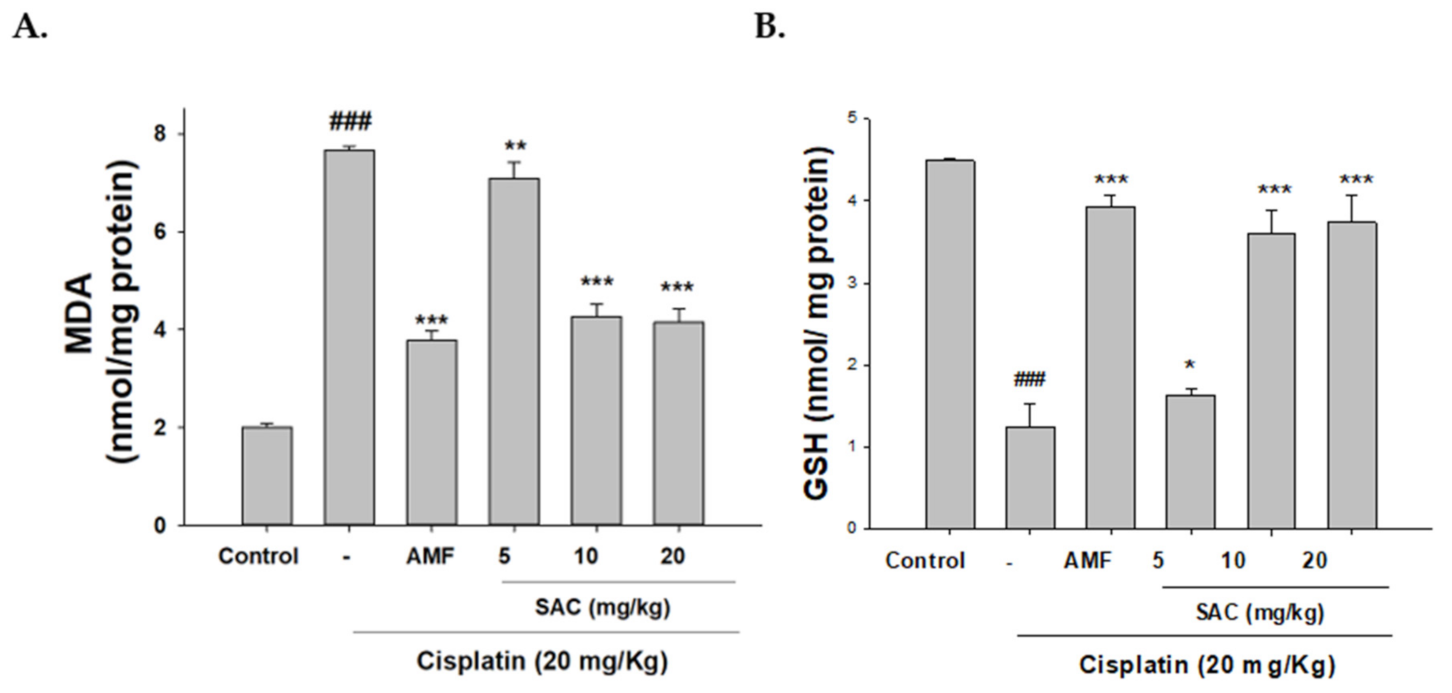
3.4. SAC Diminishes Oxidative Stress in Cisplatin-Associated Nephrotoxicity
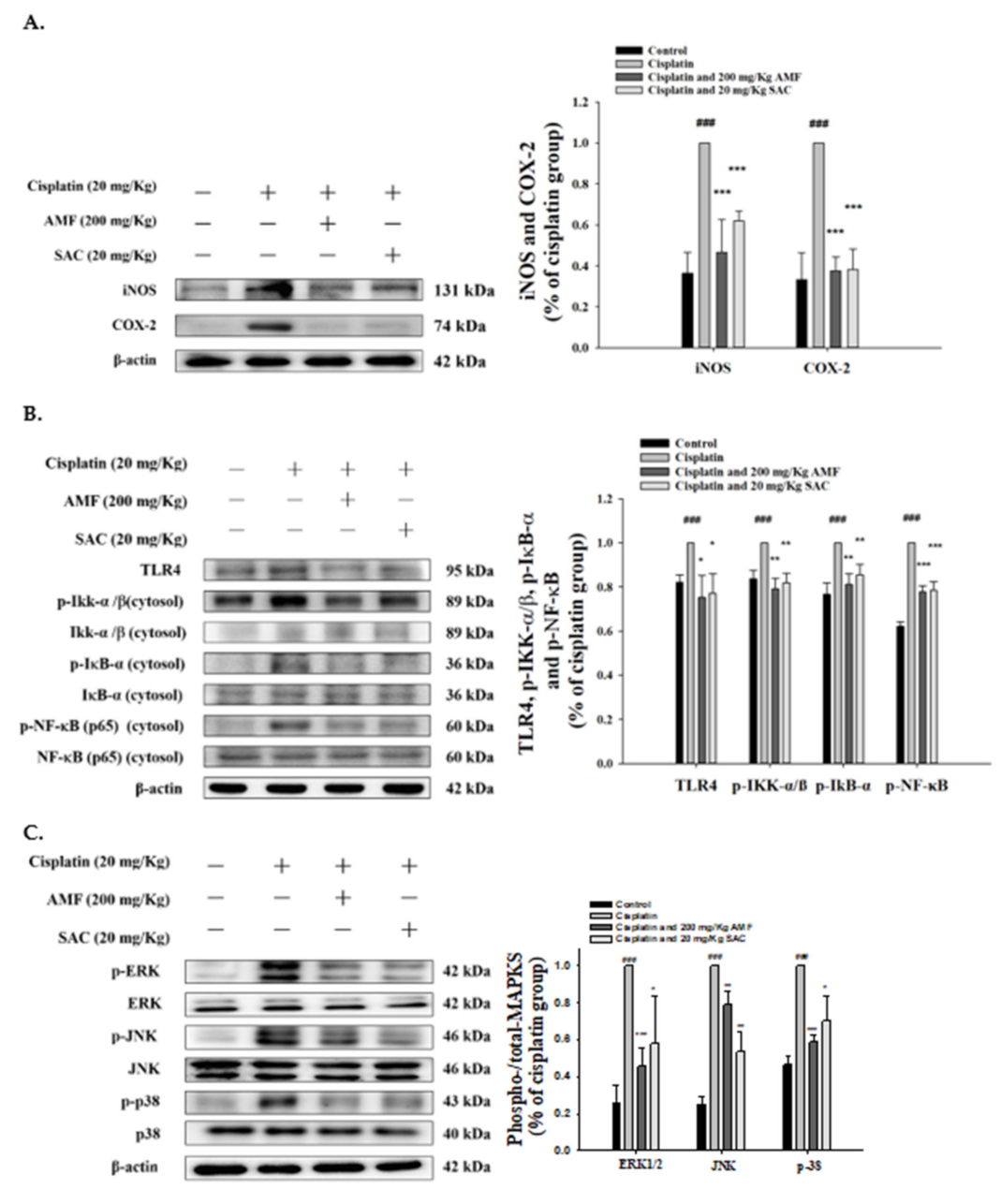
3.5. SAC Attenuated Cisplatin-Induced Inflammation in Renal Tissues
3.6. SAC-Inactivated Cisplatin Induces the MAPK Pathway in Kidneys
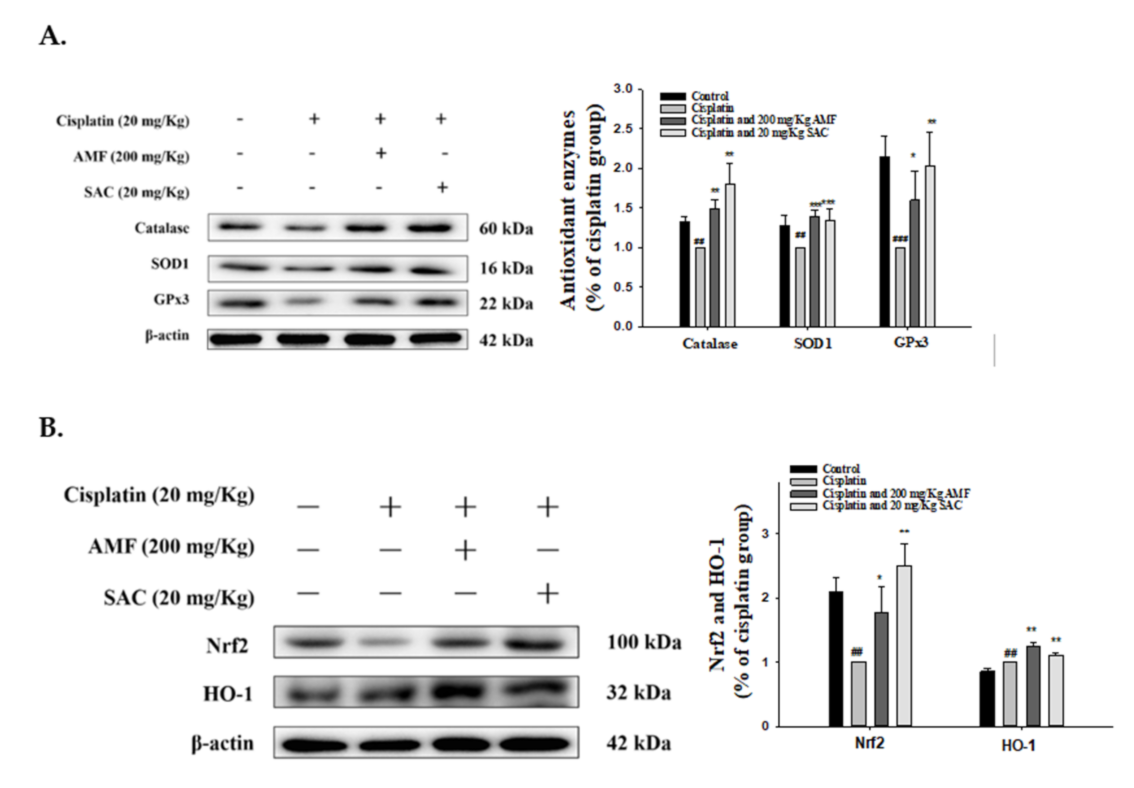
3.7. SAC Restores Renal Antioxidant Defense and the HO-1/Nrf2 Signaling Pathway in Cisplatin-Associated Nephrotoxicity
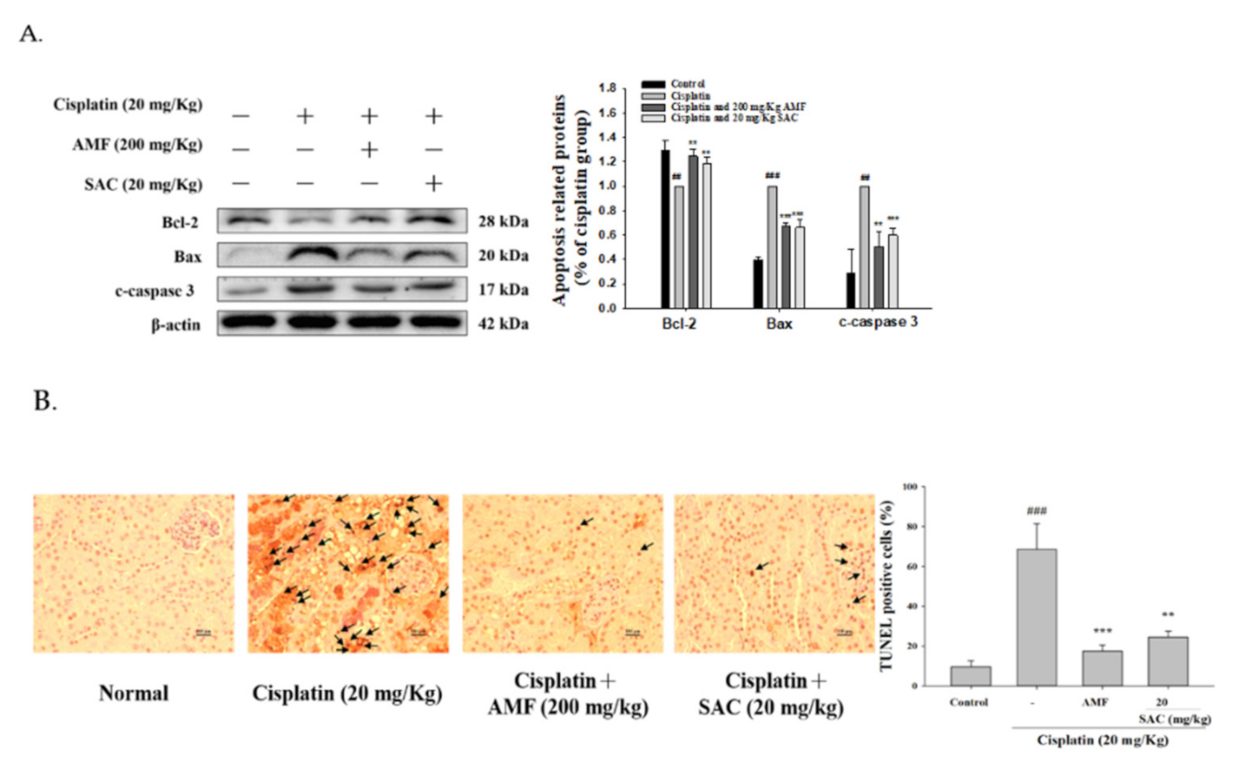
3.8. SAC Decreases the Cisplatin-Induced Apoptosis Signaling Pathway
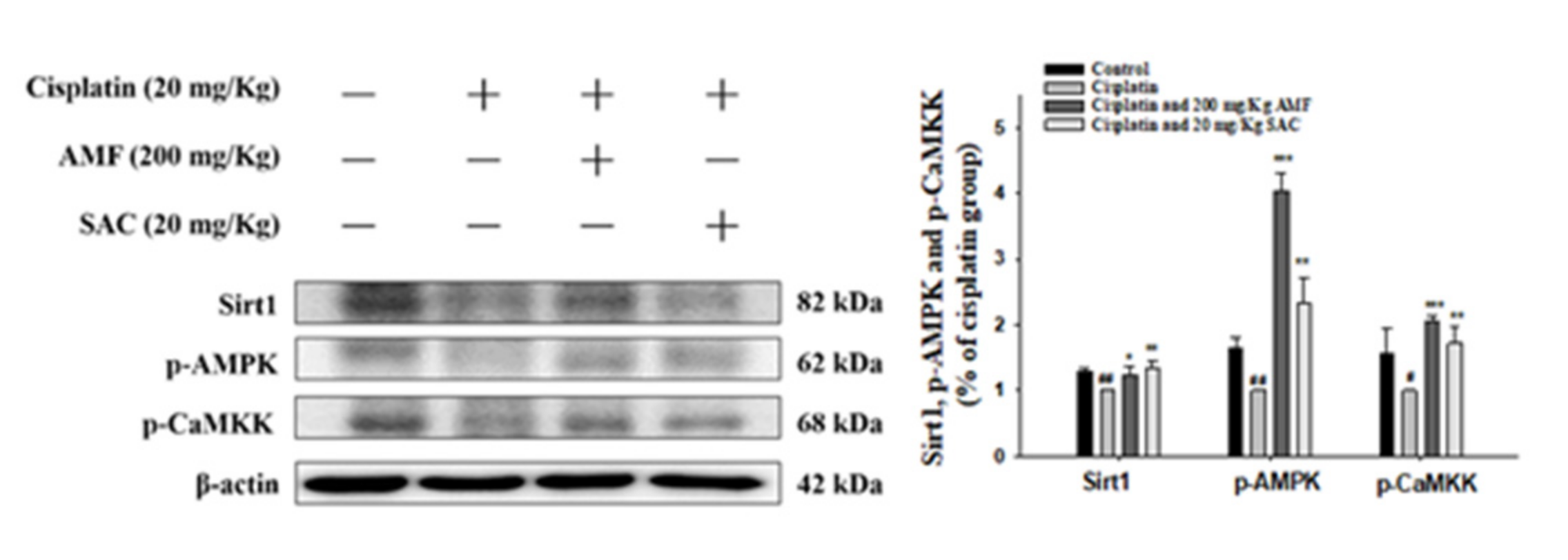
3.9. SAC Alleviates the Cisplatin-Induced CaMKK–AMPK–Sirt1 Axis
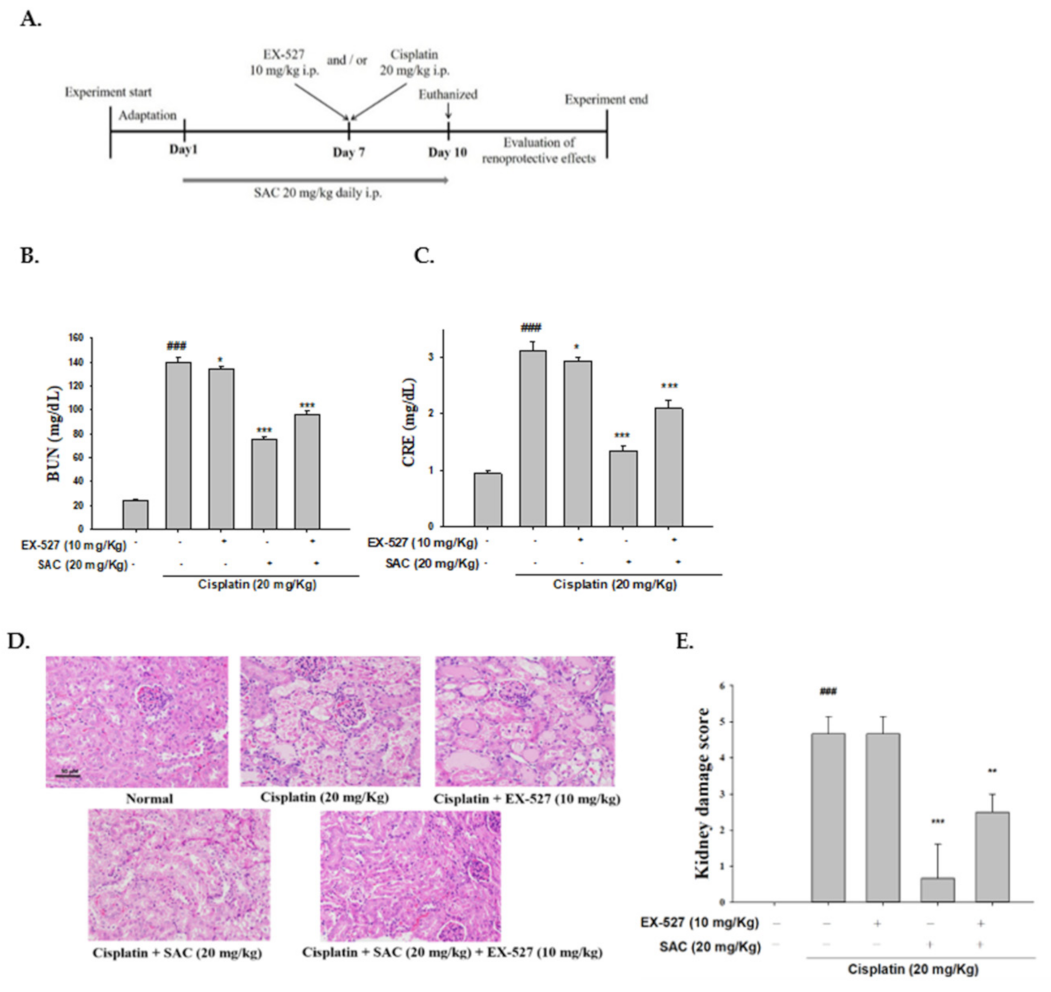
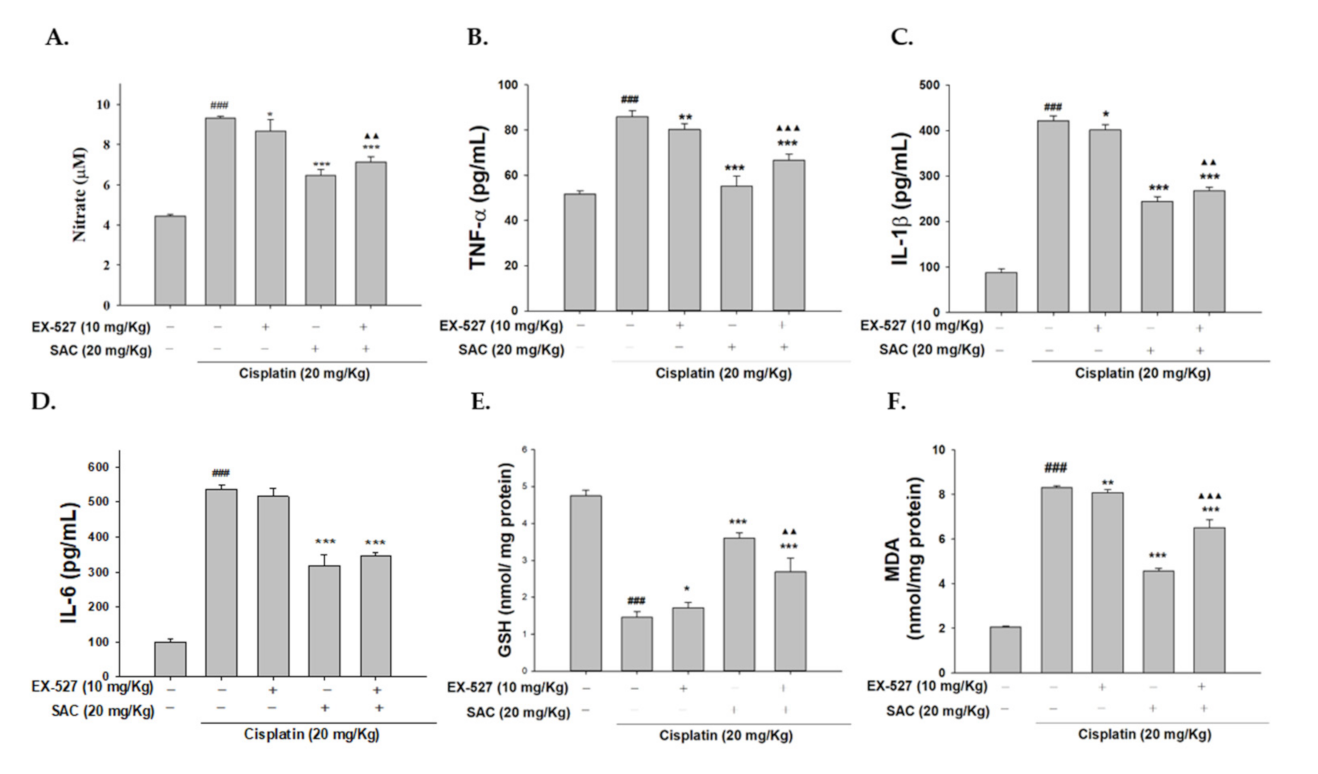
3.10. Blocking Sirt1 Synergy with EX-527 Increases Kidney Failure with Cisplatin-Induced Nephrotoxicity
3.11. SAC Demonstrate the Anti-Inflammatory Effect when Administering EX-527 as Sirt1 Blocker
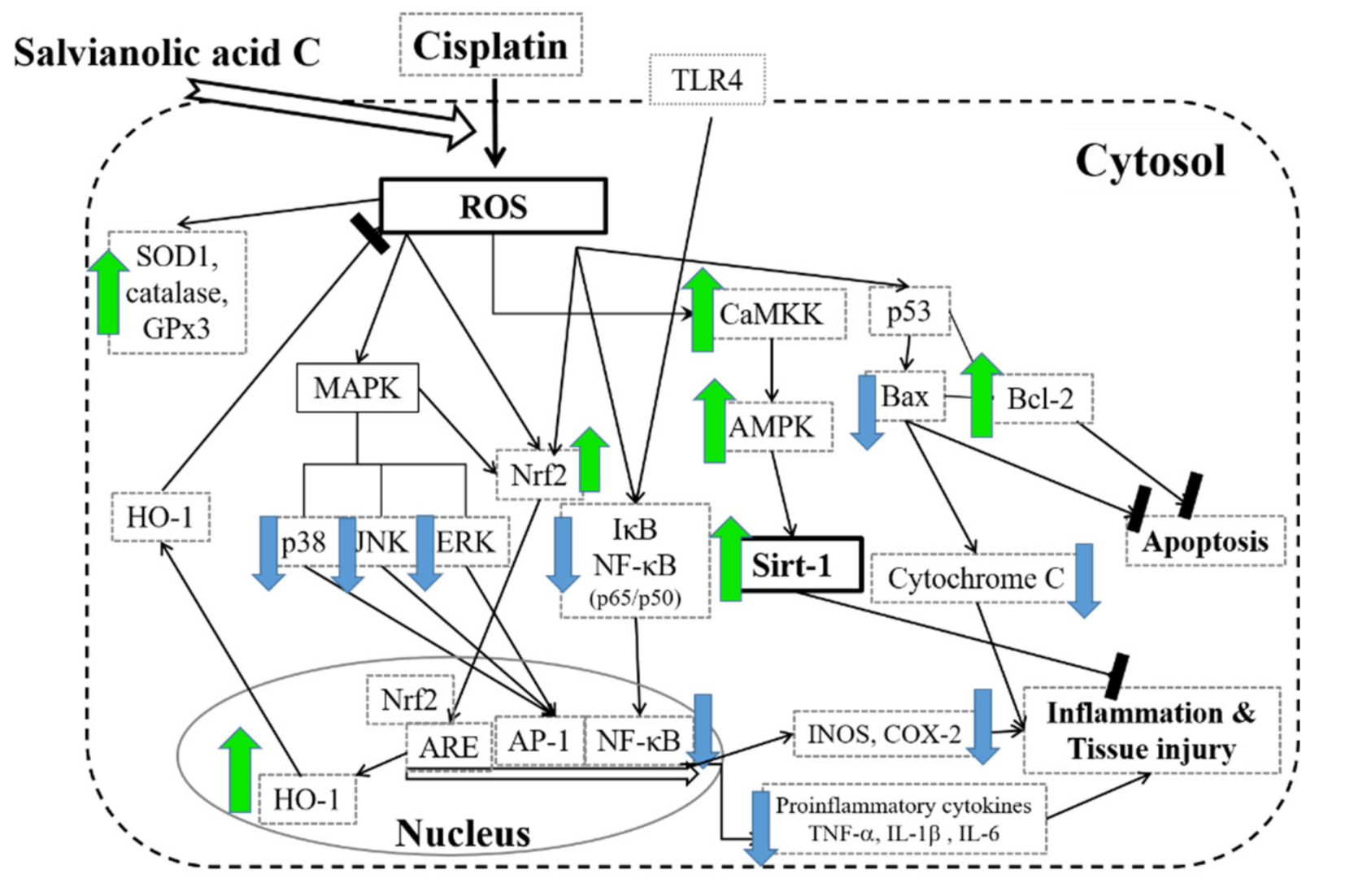
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Taber, A.; Christensen, E.; Lamy, P.; Nordentoft, I.; Prip, F.; Lindskrog, S.V.; Birkenkamp-Demtröder, K.; Okholm, T.L.H.; Knudsen, M.; Pedersen, J.S.; et al. Molecular correlates of cisplatin-based chemotherapy response in muscle invasive bladder cancer by integrated multi-omics analysis. Nat. Commun. 2020, 11, 4858. [Google Scholar] [CrossRef]
- Liu, Z.; Li, Z.; Chen, Z.; Li, C.; Lei, L.; Wu, X.; Li, Y. Numb ameliorates necrosis and inflammation in acute kidney injury induced by cisplatin. Chem. Biol. Interact. 2020, 330, 109251. [Google Scholar] [CrossRef] [PubMed]
- Din, Z.U.; Farooq, S.U.; Shahid, M.; Alghamdi, O.; Al-Hamoudi, N.; Vohra, F.; Abduljabbar, T. The flavonoid 6-hydroxyflavone prevention of cisplatin-induced nephrotoxicity. Histol. Histopathol. 2020, 35, 1197–1209. [Google Scholar] [CrossRef] [PubMed]
- Andrade-Silva, M.; Cenedeze, M.A.; Perandini, L.A.; Felizardo, R.J.F.; Watanabe, I.K.M.; Agudelo, J.S.H.; Castoldi, A.; Gonçalves, G.M.; Origassa, C.S.T.; Semedo, P.; et al. TLR2 and TLR4 play opposite role in autophagy associated with cisplatin-induced acute kidney injury. Clin. Sci. 2018, 132, 1725–1739. [Google Scholar] [CrossRef]
- Cenedeze, M.A.; Gonçalves, G.M.; Feitoza, C.Q.; Wang, P.M.; Damião, M.J.; Bertocchi, A.P.; Pacheco-Silva, A.; Câmara, N.O. The role of toll-like receptor 4 in cisplatin-induced renal injury. Transplant. Proc. 2007, 39, 409–411. [Google Scholar] [CrossRef] [PubMed]
- Yiu, W.H.; Lin, M.; Tang, S.C. Toll-like receptor activation: From renal inflammation to fibrosis. Kidney Int. Suppl. 2014, 4, 20–25. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.A.; Cozzi, M.; Bush, E.L.; Rabb, H. Distant Organ Dysfunction in Acute Kidney Injury: A Review. Am. J. Kidney Dis. 2018, 72, 846–856. [Google Scholar] [CrossRef]
- Zhang, J.; Bi, J.; Ren, Y.; Du, Z.; Li, T.; Wang, T.; Zhang, L.; Wang, M.; Wei, S.; Lv, Y.; et al. Involvement of GPX4 in irisin’s protection against ischemia reperfusion-induced acute kidney injury. J. Cell Physiol. 2021, 236, 931–945. [Google Scholar] [CrossRef]
- Kandemir, F.M.; Yildirim, S.; Caglayan, C.; Kucukler, S.; Eser, G. Protective effects of zingerone on cisplatin-induced nephrotoxicity in female rats. Environ. Sci. Pollut. Res. Int. 2019, 26, 22562–22574. [Google Scholar] [CrossRef]
- Vasaikar, N.; Mahajan, U.; Patil, K.R.; Suchal, K.; Patil, C.R.; Ojha, S.; Goyal, S.N. D-pinitol attenuates cisplatin-induced nephrotoxicity in rats: Impact on pro-inflammatory cytokines. Chem. Biol. Interact. 2018, 290, 6–11. [Google Scholar] [CrossRef]
- Deng, J.S.; Jiang, W.P.; Chen, C.C.; Lee, L.Y.; Li, P.Y.; Huang, W.C.; Liao, J.C.; Chen, H.Y.; Huang, S.S.; Huang, G.J. Cordyceps cicadae Mycelia Ameliorate Cisplatin-Induced Acute Kidney Injury by Suppressing the TLR4/NF-κB/MAPK and Activating the HO-1/Nrf2 and Sirt-1/AMPK Pathways in Mice. Oxid. Med. Cell. Longev. 2020, 2020, 7912763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azouz, A.A.; Abdel-Nassir Abdel-Razek, E.; Abo-Youssef, A.M. Amlodipine alleviates cisplatin-induced nephrotoxicity in rats through gamma-glutamyl transpeptidase (GGT) enzyme inhibition, associated with regulation of Nrf2/HO-1, MAPK/NF-κB, and Bax/Bcl-2 signaling. Saudi Pharm. J. 2020, 28, 1317–1325. [Google Scholar] [CrossRef]
- Glosse, P.; Föller, M. AMP-Activated Protein Kinase (AMPK)-Dependent Regulation of Renal Transport. Int. J. Mol. Sci. 2018, 19, 3481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farghali, H.; Kemelo, M.K.; Canová, N.K. SIRT1 Modulators in Experimentally Induced Liver Injury. Oxid. Med. Cell. Longev. 2019, 2019, 8765954. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.J.; Lee, Y.M.; Kang, S.G.; Lim, H.W.; Shin, K.O.; Jeong, S.K.; Huh, Y.H.; Choi, S.; Kor, M.; Seo, H.S.; et al. Aquatide Activation of SIRT1 Reduces Cellular Senescence through a SIRT1-FOXO1-Autophagy Axis. Biomol. Ther. 2017, 25, 511–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suetomi, T.; Miyamoto, S.; Brown, J.H. Inflammation in nonischemic heart disease: Initiation by cardiomyocyte CaMKII and NLRP3 inflammasome signaling. Am. J. Physiol. Heart Circ. Physiol. 2019, 317, H877–H890. [Google Scholar] [CrossRef] [PubMed]
- Abdou, R.M.; El-Maadawy, W.H.; Hassan, M.; El-Dine, R.S.; Aboushousha, T.; El-Tanbouly, N.D.; El-Sayed, A.M. Nephroprotective activity of Aframomum melegueta seeds extract against diclofenac-induced acute kidney injury: A mechanistic study. J. Ethnopharmacol. 2021, 273, 113939. [Google Scholar] [CrossRef]
- Ren, H.; Shao, Y.; Wu, C.; Ma, X.; Lv, C.; Wang, Q. Metformin alleviates oxidative stress and enhances autophagy in diabetic kidney disease via AMPK/SIRT1-FoxO1 pathway. Mol. Cell. Endocrinol. 2020, 500, 110628. [Google Scholar] [CrossRef] [PubMed]
- Jing, Z.; He, X.; Jia, Z.; Sa, Y.; Yang, B.; Liu, P. NCAPD2 inhibits autophagy by regulating Ca(2+)/CAMKK2/AMPK/mTORC1 pathway and PARP-1/SIRT1 axis to promote colorectal cancer. Cancer Lett. 2021, 520, 26–37. [Google Scholar] [CrossRef]
- Hao, X.; Pu, Z.; Cao, G.; You, D.; Zhou, Y.; Deng, C.; Shi, M.; Nile, S.H.; Wang, Y.; Zhou, W.; et al. Tanshinone and salvianolic acid biosynthesis are regulated by SmMYB98 in Salvia miltiorrhiza hairy roots. J. Adv. Res. 2020, 23, 1–12. [Google Scholar] [CrossRef]
- Lin, S.H.; Huang, K.J.; Weng, C.F.; Shiuan, D. Exploration of natural product ingredients as inhibitors of human HMG-CoA reductase through structure-based virtual screening. Drug Des. Dev. Ther. 2015, 9, 3313–3324. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.; Han, N.; Yu, H.; Wang, L.; Li, X.; Dong, Z.; Fu, W.; Yorinaka, H.; Cho, K.; Wu, W.; et al. Amelioration of salvianolic acid C on aortic structure in apolipoprotein E-deficient mice treated with angiotension II. Life Sci. 2016, 166, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Mercantepe, F.; Mercantepe, T.; Topcu, A.; Yılmaz, A.; Tumkaya, L. Protective effects of amifostine, curcumin, and melatonin against cisplatin-induced acute kidney injury. Naunyn Schmiedebergs Arch. Pharmacol. 2018, 391, 915–931. [Google Scholar] [CrossRef] [PubMed]
- Uzunoglu, S.; Karagol, H.; Ozpuyan, F.; Cosar, R.; Cicin, I.; Yurutcaloglu, V.; Denizli, B.; Tanriverdi, Ö.; Sut, N.; Kocak, Z. Protective effect of L-carnitine versus amifostine against cisplatin-induced nephrotoxicity in rats. Med. Oncol. 2011, 28 (Suppl. 1), S690–S696. [Google Scholar] [CrossRef]
- Ren, Q.; Guo, F.; Tao, S.; Huang, R.; Ma, L.; Fu, P. Flavonoid fisetin alleviates kidney inflammation and apoptosis via inhibiting Src-mediated NF-κB p65 and MAPK signaling pathways in septic AKI mice. Biomed. Pharmacother. 2020, 122, 109772. [Google Scholar] [CrossRef]
- Aguilar Diaz De Leon, J.; Borges, C.R. Evaluation of Oxidative Stress in Biological Samples Using the Thiobarbituric Acid Reactive Substances Assay. J. Vis. Exp. 2020, e61122. [Google Scholar] [CrossRef]
- Vargas-Maya, N.I.; Padilla-Vaca, F.; Romero-González, O.E.; Rosales-Castillo, E.A.S.; Rangel-Serrano, Á.; Arias-Negrete, S.; Franco, B. Refinement of the Griess method for measuring nitrite in biological samples. J. Microbiol. Methods 2021, 187, 106260. [Google Scholar] [CrossRef]
- Fakurazi, S.; Hairuszah, I.; Nanthini, U. Moringa oleifera Lam prevents acetaminophen induced liver injury through restoration of glutathione level. Food Chem. Toxicol. 2008, 46, 2611–2615. [Google Scholar] [CrossRef]
- Cao, W.Q.; Zhai, X.Q.; Ma, J.W.; Fu, X.Q.; Zhao, B.S.; Zhang, P.; Fu, X.Y. Natural borneol sensitizes human glioma cells to cisplatin-induced apoptosis by triggering ROS-mediated oxidative damage and regulation of MAPKs and PI3K/AKT pathway. Pharm. Biol. 2020, 58, 72–79. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Park, D.J.; Kim, J.H.; Jeong, E.Y.; Jung, M.H.; Kim, T.H.; Yang, J.I.; Lee, G.W.; Chung, H.J.; Chang, S.H. Glutamine protects against cisplatin-induced nephrotoxicity by decreasing cisplatin accumulation. J. Pharmacol. Sci. 2015, 127, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Nezu, M.; Suzuki, N. Roles of Nrf2 in Protecting the Kidney from Oxidative Damage. Int. J. Mol. Sci. 2020, 21, 2951. [Google Scholar] [CrossRef]
- Liu, X.Y.; Liu, F.C.; Deng, C.Y.; Zhang, M.Z.; Yang, M.; Xiao, D.Z.; Lin, Q.X.; Cai, S.T.; Kuang, S.J.; Chen, J.; et al. Left ventricular deformation associated with cardiomyocyte Ca(2+) transients delay in early stage of low-dose of STZ and high-fat diet induced type 2 diabetic rats. BMC Cardiovasc. Disord. 2016, 16, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozkok, A.; Edelstein, C.L. Pathophysiology of cisplatin-induced acute kidney injury. BioMed Res. Int. 2014, 2014, 967826. [Google Scholar] [CrossRef]
- Wu, C.F.; Bohnert, S.; Thines, E.; Efferth, T. Cytotoxicity of Salvia miltiorrhiza Against Multidrug-Resistant Cancer Cells. Am. J. Chin. Med. 2016, 44, 871–894. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.L.; Yan, L.; Zhu, L.; Jiao, D.M.; Chen, J.; Chen, Q.Y. Salvianolic acid A reverses cisplatin resistance in lung cancer A549 cells by targeting c-met and attenuating Akt/mTOR pathway. J. Pharmacol. Sci. 2017, 135, 1–7. [Google Scholar] [CrossRef]
- Lin, Z.; Bao, Y.; Hong, B.; Wang, Y.; Zhang, X.; Wu, Y. Salvianolic acid B attenuated cisplatin-induced cardiac injury and oxidative stress via modulating Nrf2 signal pathway. J. Toxicol. Sci. 2021, 46, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.M.; Ren, G.L.; Gao, L.; Yang, Q.; Li, H.D.; Wu, W.F.; Huang, C.; Zhang, L.; Lv, X.W.; Li, J. NADPH oxidase 4 promotes cisplatin-induced acute kidney injury via ROS-mediated programmed cell death and inflammation. Lab. Investig. 2018, 98, 63–78. [Google Scholar] [CrossRef] [Green Version]
- Perše, M.; Večerić-Haler, Ž. Cisplatin-Induced Rodent Model of Kidney Injury: Characteristics and Challenges. BioMed Res. Int. 2018, 2018, 1462802. [Google Scholar] [CrossRef]
- Chen, X.; Wei, W.; Li, Y.; Huang, J.; Ci, X. Hesperetin relieves cisplatin-induced acute kidney injury by mitigating oxidative stress, inflammation and apoptosis. Chem. Biol. Interact. 2019, 308, 269–278. [Google Scholar] [CrossRef]
- Sharp, C.N.; Siskind, L.J. Developing better mouse models to study cisplatin-induced kidney injury. Am. J. Physiol. Renal Physiol. 2017, 313, F835–F841. [Google Scholar] [CrossRef] [Green Version]
- Divya, M.K.; Lincy, L.; Raghavamenon, A.C.; Babu, T.D. Ameliorative effect of Apodytes dimidiata on cisplatin-induced nephrotoxicity in Wistar rats. Pharm. Biol. 2016, 54, 2149–2157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, I.; Umemura, M.; Mitsudo, K.; Kioi, M.; Nakashima, H.; Iwai, T.; Feng, X.; Oda, K.; Miyajima, A.; Makino, A.; et al. Hyperthermia generated with ferucarbotran (Resovist®) in an alternating magnetic field enhances cisplatin-induced apoptosis of cultured human oral cancer cells. J. Physiol. Sci. 2014, 64, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Gui, Y.; Ren, J.; Liu, X.; Feng, Y.; Zeng, Z.; He, W.; Yang, J.; Dai, C. Metformin Protects Against Cisplatin-Induced Tubular Cell Apoptosis and Acute Kidney Injury via AMPKα-regulated Autophagy Induction. Sci. Rep. 2016, 6, 23975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.; Dang, C.; Kang, H.; Dai, Z.; Lin, S.; Guan, H.; Liu, X.; Wang, X.; Hui, W. Saikosaponin-D reduces cisplatin-induced nephrotoxicity by repressing ROS-mediated activation of MAPK and NF-κB signalling pathways. Int. Immunopharmacol. 2015, 28, 399–408. [Google Scholar] [CrossRef]
- Tarang, S.; Sodhi, A.; Chauhan, P. Differential expression of Toll-like receptors in murine peritoneal macrophages in vitro on treatment with cisplatin. Int. Immunol. 2007, 19, 635–643. [Google Scholar] [CrossRef]
- Zhang, B.; Ramesh, G.; Uematsu, S.; Akira, S.; Reeves, W.B. TLR4 signaling mediates inflammation and tissue injury in nephrotoxicity. J. Am. Soc. Nephrol. 2008, 19, 923–932. [Google Scholar] [CrossRef]
- Ramesh, G.; Kimball, S.R.; Jefferson, L.S.; Reeves, W.B. Endotoxin and cisplatin synergistically stimulate TNF-alpha production by renal epithelial cells. Am. J. Physiol. Renal Physiol. 2007, 292, F812–F819. [Google Scholar] [CrossRef]
- Wang, S.; Tang, S.; Chen, X.; Li, X.; Jiang, S.; Li, H.P.; Jia, P.H.; Song, M.J.; Di, P.; Li, W. Pulchinenoside B4 exerts the protective effects against cisplatin-induced nephrotoxicity through NF-κB and MAPK mediated apoptosis signaling pathways in mice. Chem. Biol. Interact. 2020, 331, 109233. [Google Scholar] [CrossRef]
- Ma, X.; Yan, L.; Zhu, Q.; Shao, F. Puerarin attenuates cisplatin-induced rat nephrotoxicity: The involvement of TLR4/NF-κB signaling pathway. PLoS ONE 2017, 12, e0171612. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Yang, Y.; Li, Y.; Zhao, Y.; Jiang, H. Sirt5 Attenuates Cisplatin-Induced Acute Kidney Injury through Regulation of Nrf2/HO-1 and Bcl-2. Biomed. Res. Int. 2019, 2019, 4745132. [Google Scholar] [CrossRef]
- Zhu, L.; Yuan, Y.; Yuan, L.; Li, L.; Liu, F.; Liu, J.; Chen, Y.; Lu, Y.; Cheng, J. Activation of TFEB-mediated autophagy by trehalose attenuates mitochondrial dysfunction in cisplatin-induced acute kidney injury. Theranostics 2020, 10, 5829–5844. [Google Scholar] [CrossRef]
- Ogueji, E.; Nwani, C.; Mbah, C.; Iheanacho, S.; Nweke, F. Oxidative stress, biochemical, lipid peroxidation, and antioxidant responses in Clarias gariepinus exposed to acute concentrations of ivermectin. Environ. Sci. Pollut. Res. Int. 2020, 27, 16806–16815. [Google Scholar] [CrossRef]
- Zheng, Y.; Lu, H.; Huang, H. Desflurane Preconditioning Protects Against Renal Ischemia-Reperfusion Injury and Inhibits Inflammation and Oxidative Stress in Rats Through Regulating the Nrf2-Keap1-ARE Signaling Pathway. Drug Des. Dev. Ther. 2020, 14, 1351–1362. [Google Scholar] [CrossRef] [Green Version]
- Sun, P.; Bu, F.; Min, J.W.; Munshi, Y.; Howe, M.D.; Liu, L.; Koellhoffer, E.C.; Qi, L.; McCullough, L.D.; Li, J. Inhibition of calcium/calmodulin-dependent protein kinase kinase (CaMKK) exacerbates impairment of endothelial cell and blood-brain barrier after stroke. Eur. J. Neurosci. 2019, 49, 27–39. [Google Scholar] [CrossRef]
- Wang, S.Y.; Cai, G.Y.; Chen, X.M. Energy restriction in renal protection. Br. J. Nutr. 2018, 120, 1149–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Howell, G.M.; Guo, L.; Collage, R.D.; Loughran, P.A.; Zuckerbraun, B.S.; Rosengart, M.R. CaMKIV-dependent preservation of mTOR expression is required for autophagy during lipopolysaccharide-induced inflammation and acute kidney injury. J. Immunol. 2014, 193, 2405–2415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.Y.; Lim, J.H.; Youn, H.H.; Hong, Y.A.; Yang, K.S.; Park, H.S.; Chung, S.; Ko, S.H.; Shin, S.J.; Choi, B.S.; et al. Resveratrol prevents renal lipotoxicity and inhibits mesangial cell glucotoxicity in a manner dependent on the AMPK-SIRT1-PGC1α axis in db/db mice. Diabetologia 2013, 56, 204–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, S.M.; Chandel, N.S.; Hay, N. AMPK regulates NADPH homeostasis to promote tumour cell survival during energy stress. Nature 2012, 485, 661–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bairwa, S.C.; Parajuli, N.; Dyck, J.R. The role of AMPK in cardiomyocyte health and survival. Biochim. Biophys. Acta 2016, 1862, 2199–2210. [Google Scholar] [CrossRef]
- Jiang, Z.; Weng, P.; Xu, X.; Li, M.; Li, Y.; Lv, Y.; Chang, K.; Wang, S.; Lin, G.; Hu, C. IRF9 promotes apoptosis and innate immunity by inhibiting SIRT1-p53 axis in fish. Fish Shellfish Immunol. 2020, 103, 220–228. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Initial Body Weight (g) | Final Body Weight (g) | Kidney Index (mg/g) |
|---|---|---|---|
| Control | 29.8 ± 0.53 | 31.87 ± 0.45 | 1.34 ± 0.02 |
| Cisplatin (20 mg/kg) | 29.93 ± 0.65 | 25.72 ± 0.98 ### | 2.22 ± 0.08 ### |
| Cisplatin (20 mg/kg) + AMF (200 mg/kg) | 30.03 ± 0.21 | 30.27 ± 0.21 *** | 1.51 ± 0.02 *** |
| Cisplatin (20 mg/kg) + SAC (5 mg/kg) | 29.85 ± 0.39 | 26.62 ± 0.52 | 2.07 ± 0.07 ** |
| Cisplatin (20 mg/kg) + SAC (10 mg/kg) | 30.1 ± 0.28 | 29.1 ± 0.53 *** | 1.66 ± 0.04 *** |
| Cisplatin (20 mg/kg) + SAC (20 mg/kg) | 30.0 ± 0.37 | 29.45 ± 0.17 *** | 1.56 ± 0.02 *** |
| Groups | Initial Body (g) | Final Body (g) | Kidney Index (mg/g) |
|---|---|---|---|
| Control | 33.78 ± 0.66 | 37.39 ± 1.08 | 1.44 ± 0.04 |
| Cisplatin (20 mg/kg) | 33.39 ± 0.30 | 32.41 ± 0.62 ### | 2.46 ± 0.04 ### |
| Cisplatin (20 mg/kg) + SAC (20 mg/kg) | 33.65 ± 0.30 | 34.96 ± 0.51 *** | 1.55 ± 0.02 *** |
| Cisplatin (20 mg/kg) + EX-527 (10 mg/kg) | 33.43 ± 0.33 | 32.86 ± 0.22 | 2.39 ± 0.04 ** |
| Cisplatin (20 mg/kg) + SAC (20 mg/kg) + EX-527 (10 mg/kg) | 33.46 ± 0.19 | 33.9 ± 0.58 ** | 1.69 ± 0.03 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chien, L.-H.; Wu, C.-T.; Deng, J.-S.; Jiang, W.-P.; Huang, W.-C.; Huang, G.-J. Salvianolic Acid C Protects against Cisplatin-Induced Acute Kidney Injury through Attenuation of Inflammation, Oxidative Stress and Apoptotic Effects and Activation of the CaMKK–AMPK–Sirt1-Associated Signaling Pathway in Mouse Models. Antioxidants 2021, 10, 1620. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10101620
Chien L-H, Wu C-T, Deng J-S, Jiang W-P, Huang W-C, Huang G-J. Salvianolic Acid C Protects against Cisplatin-Induced Acute Kidney Injury through Attenuation of Inflammation, Oxidative Stress and Apoptotic Effects and Activation of the CaMKK–AMPK–Sirt1-Associated Signaling Pathway in Mouse Models. Antioxidants. 2021; 10(10):1620. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10101620
Chicago/Turabian StyleChien, Liang-Hsuan, Chien-Ta Wu, Jeng-Shyan Deng, Wen-Ping Jiang, Wen-Chin Huang, and Guan-Jhong Huang. 2021. "Salvianolic Acid C Protects against Cisplatin-Induced Acute Kidney Injury through Attenuation of Inflammation, Oxidative Stress and Apoptotic Effects and Activation of the CaMKK–AMPK–Sirt1-Associated Signaling Pathway in Mouse Models" Antioxidants 10, no. 10: 1620. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10101620