
Protective Effect of Anthocyanin-Enriched Polyphenols from Hibiscus syriacus L. (Malvaceae) against Ultraviolet B-Induced Damage


 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of AHs
2.2. Reagents and Antibodies
2.3. Cell Culture and Cell Viability
2.4. Analysis of Cell Viability and Population of Dead Cells
2.5. UVB Irradiation
2.6. Annexin V Staining for Apoptosis
2.7. Analysis of Caspase-3/7 Activity
2.8. Analysis of Intracellular Reactive Oxygen Species (ROS)
2.9. Mitochondrial Membranes Potential and Mitochondrial ROS (mtROS)
2.10. Western Blotting
2.11. Maintenance of Zebrafish
2.12. Evaluation of Survival Rate and Hatchability in UVB-Irradiated Zebrafish Larvae and Embryos
2.13. Detection of Total ROS and mitoROS in Zebrafish Larvae
2.14. Statistical Analysis
3. Results
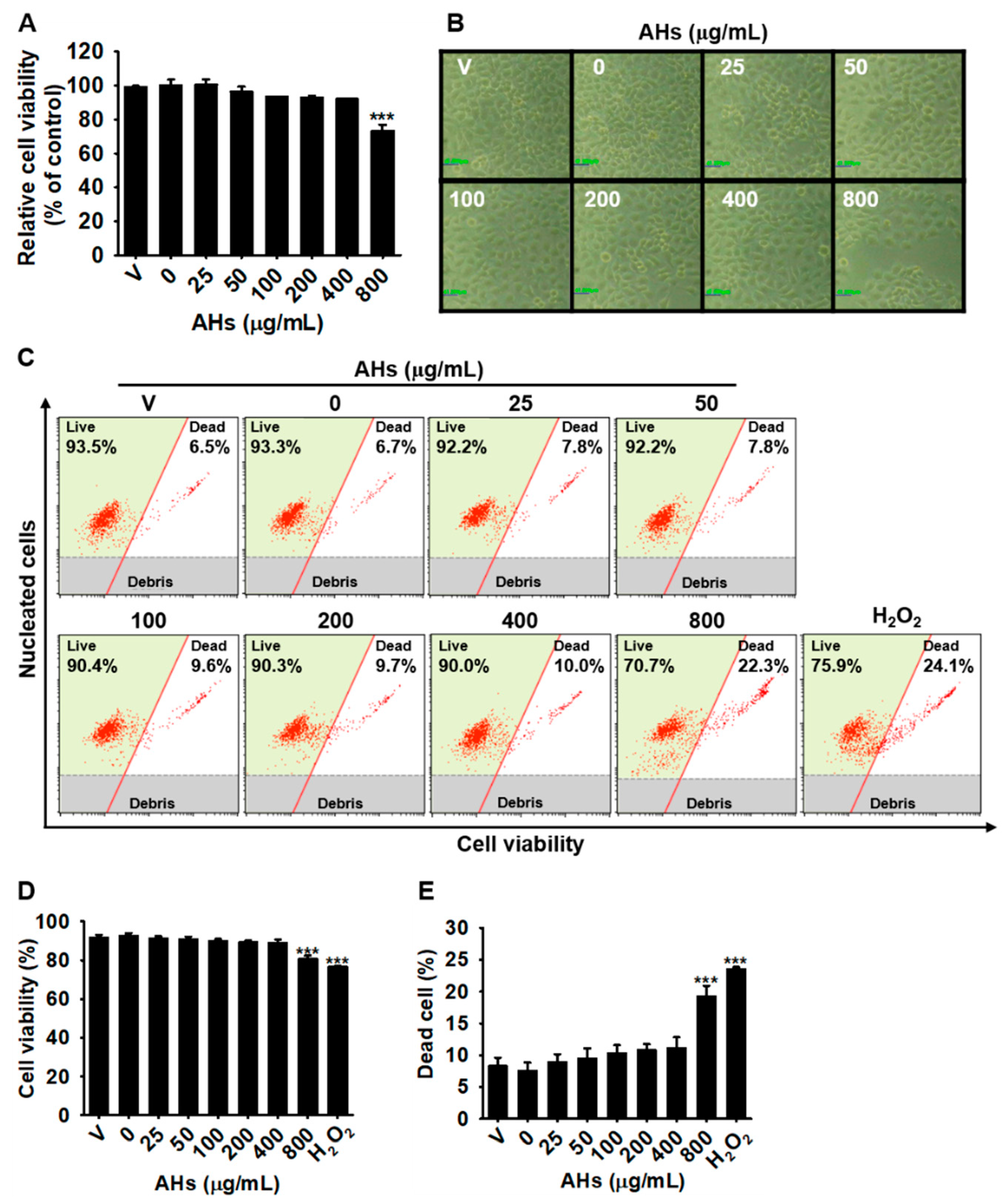
3.1. Low Concentrations of AHs Are Not Cytotoxic in HaCaT Keratinocytes
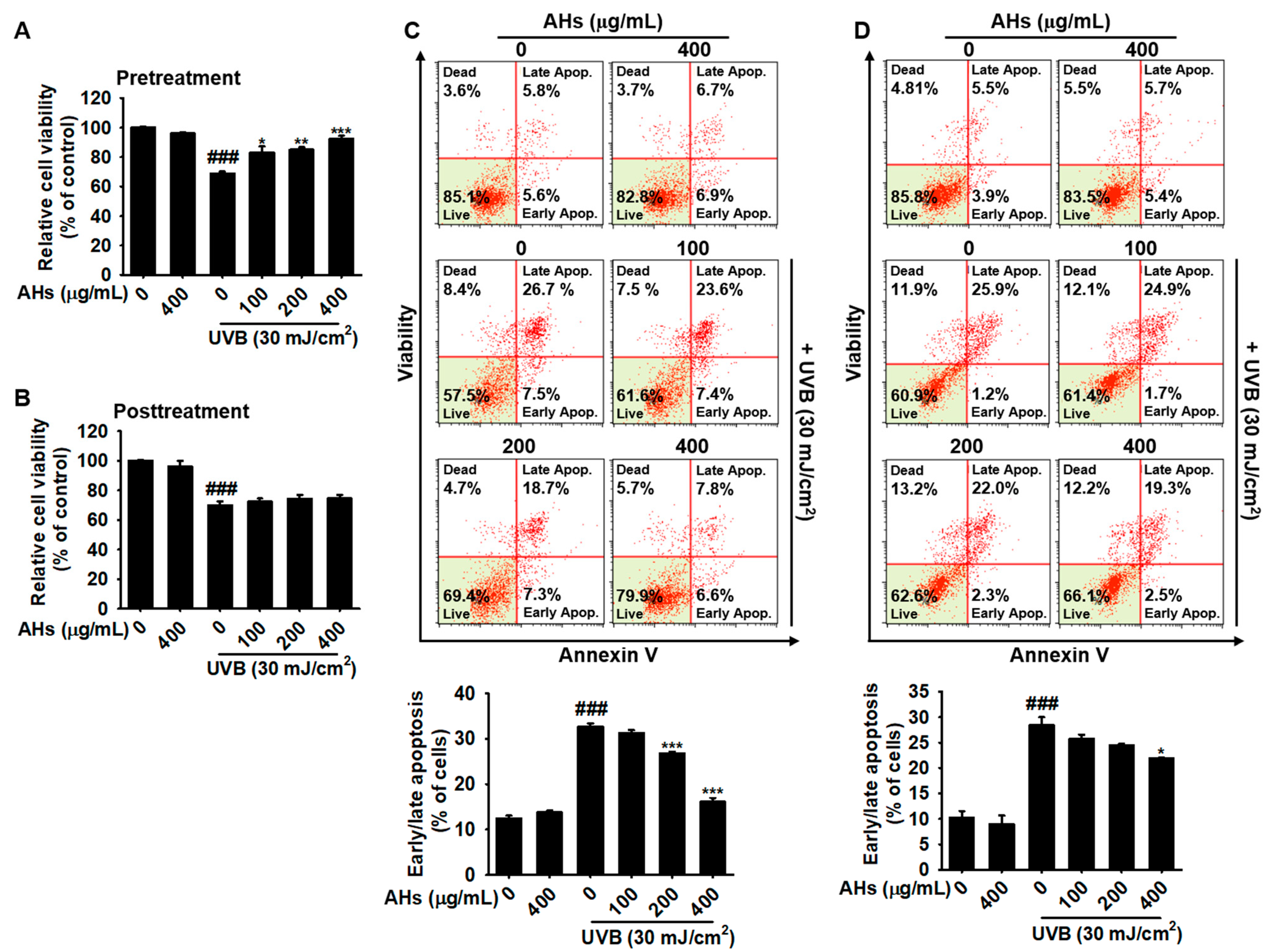
3.2. Pretreatment, but Not Posttreatment with AHs Protects HaCaT Keratinocytes from UVB-Induced Apoptosis
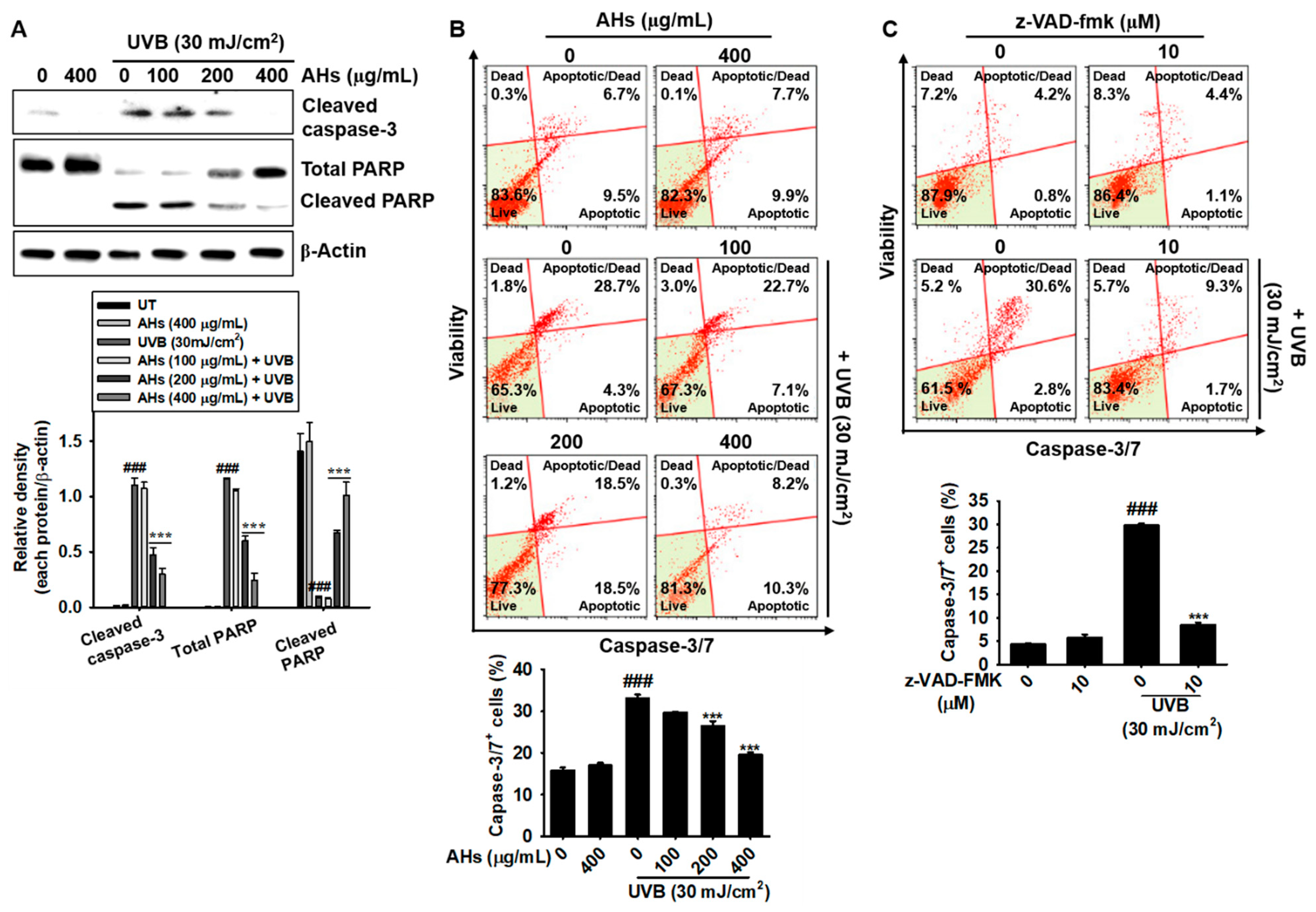
3.3. AHs Prevents UVB-Induced Apoptosis by Inhibiting the Caspase-Mediated Signaling Pathway
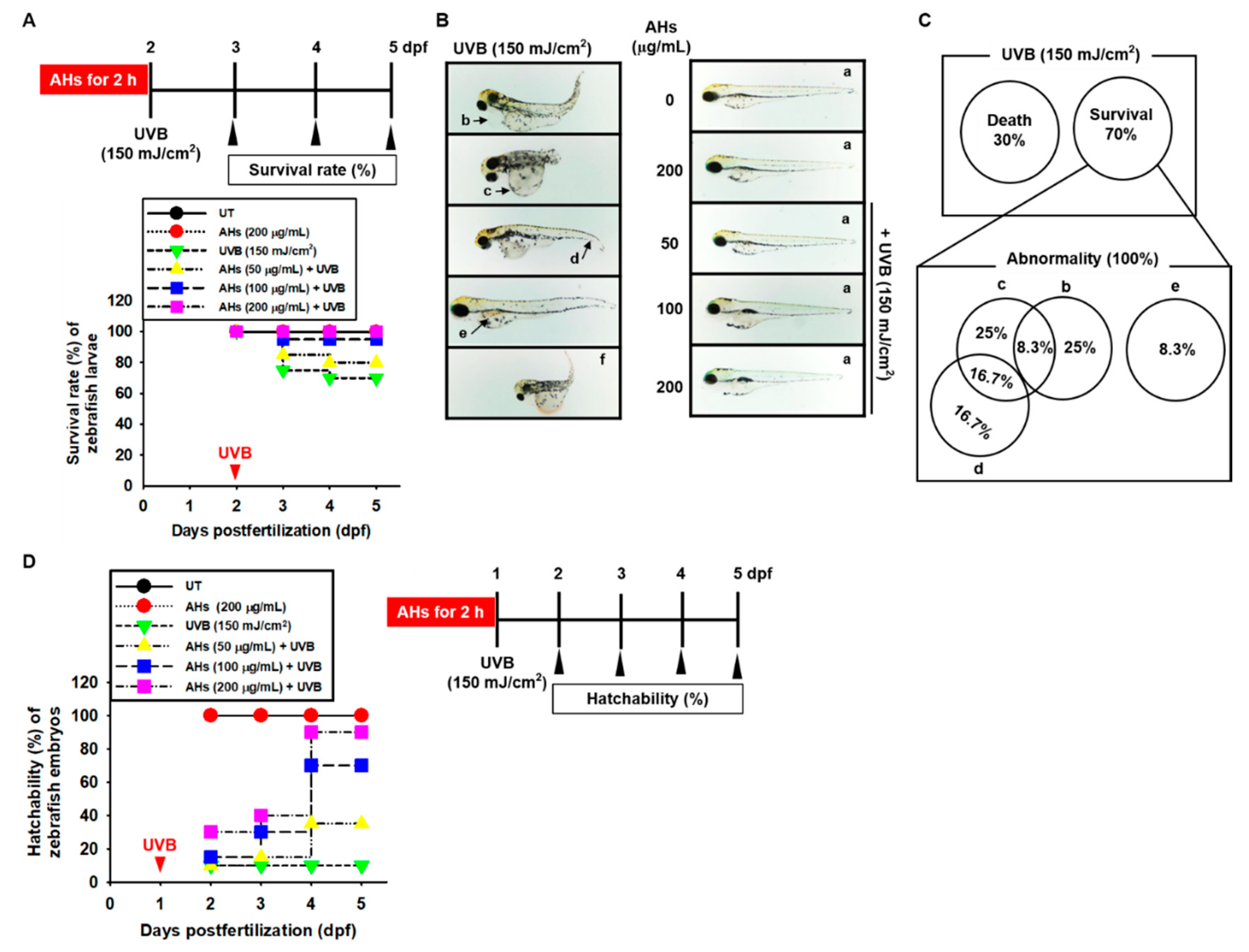
3.4. AHs Decreases Mortality and Morphological Abnormalities in UVB-Irradiated Zebrafish Larvae and Increases Hatchability in UVB-Irradiated Zebrafish Embryos
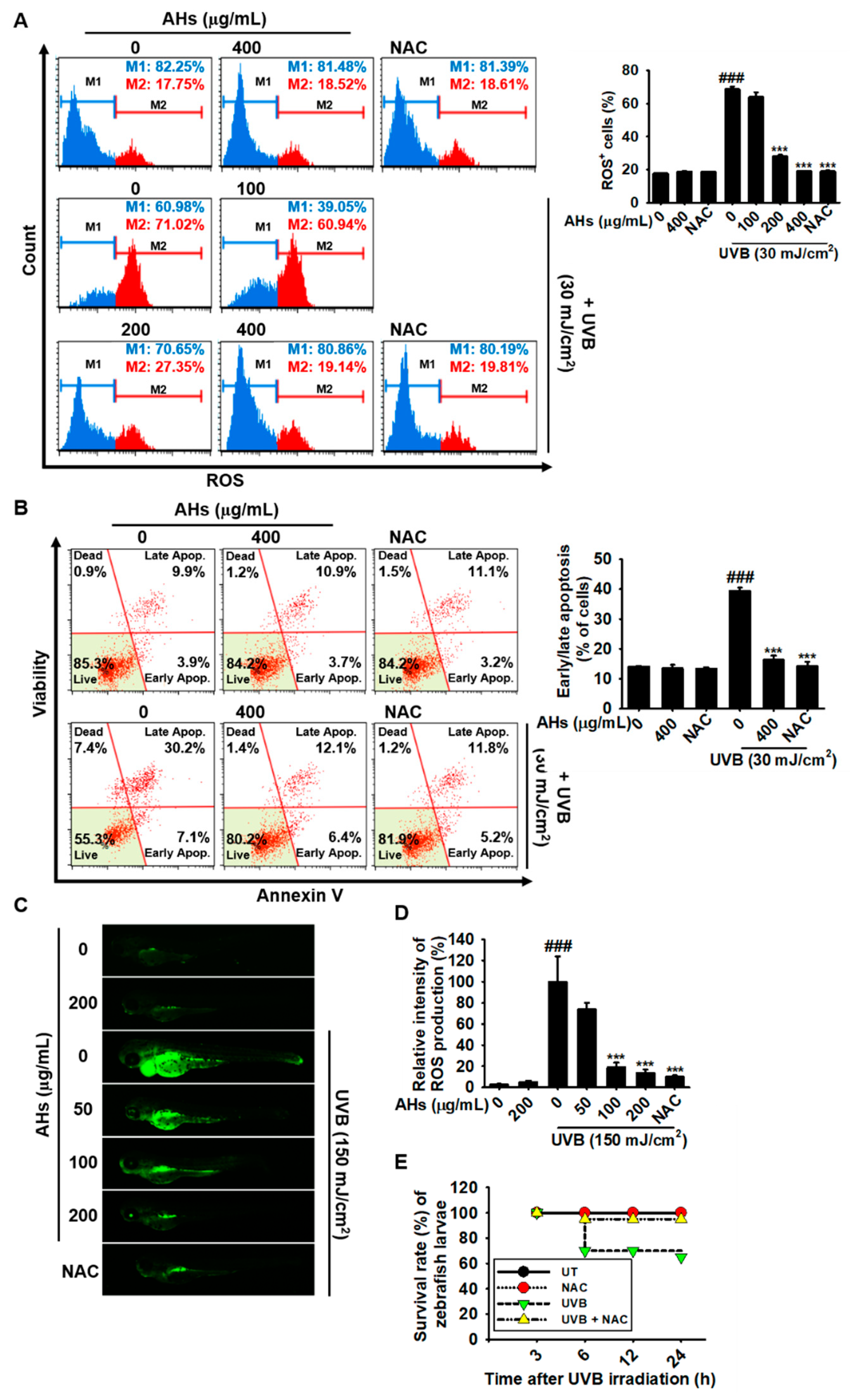
3.5. AHs Prevents UVB-Induced Apoptosis in HaCaT Keratinocytes and Mortality in Zebrafish Larvae by Inhibiting Intracellular ROS Production
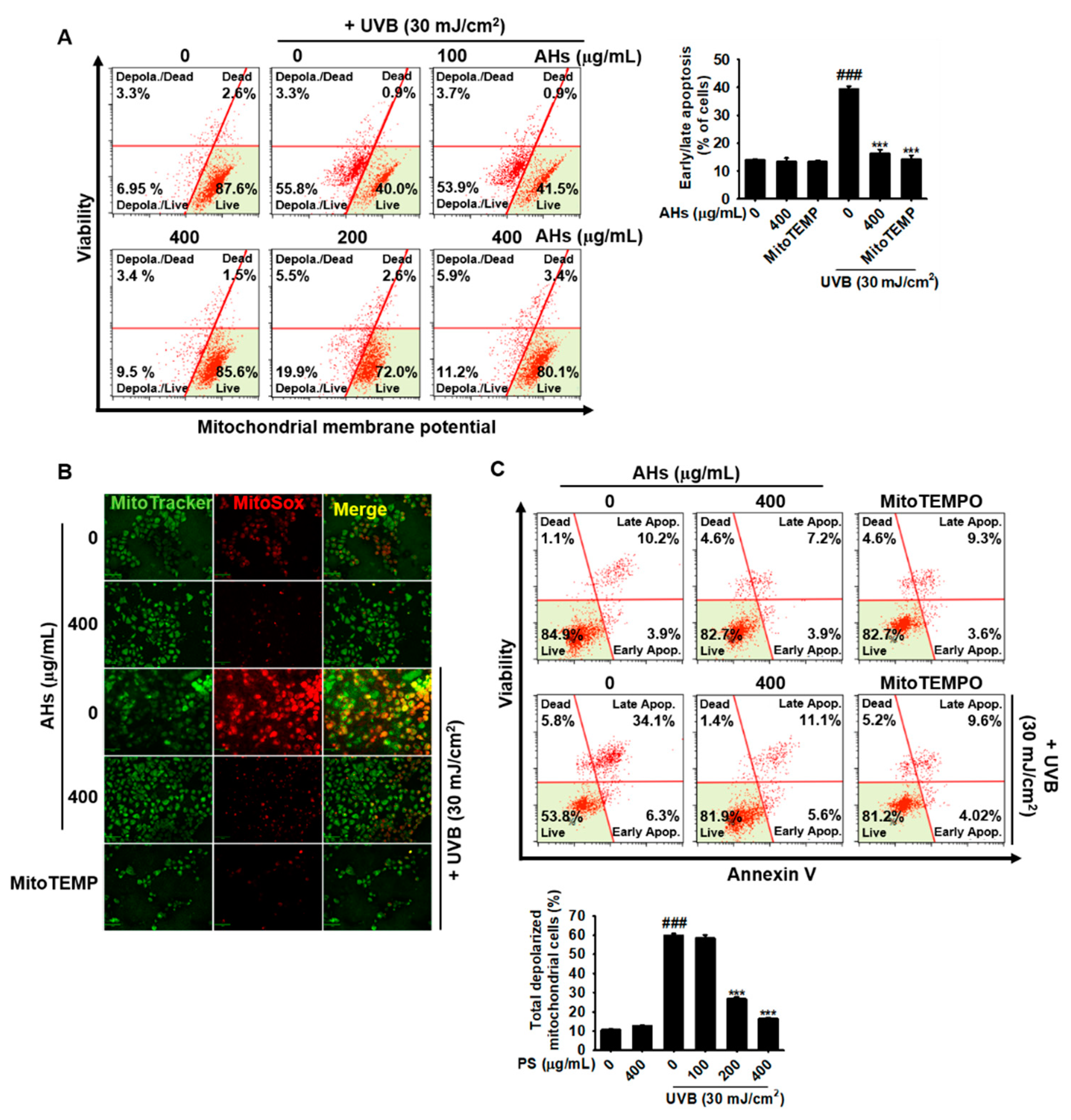
3.6. AHs Decreases the Depolarization of the Mitochondrial Membrane Potential and mtROS Production in UVB-Irradiated HaCaT Keratinocytes
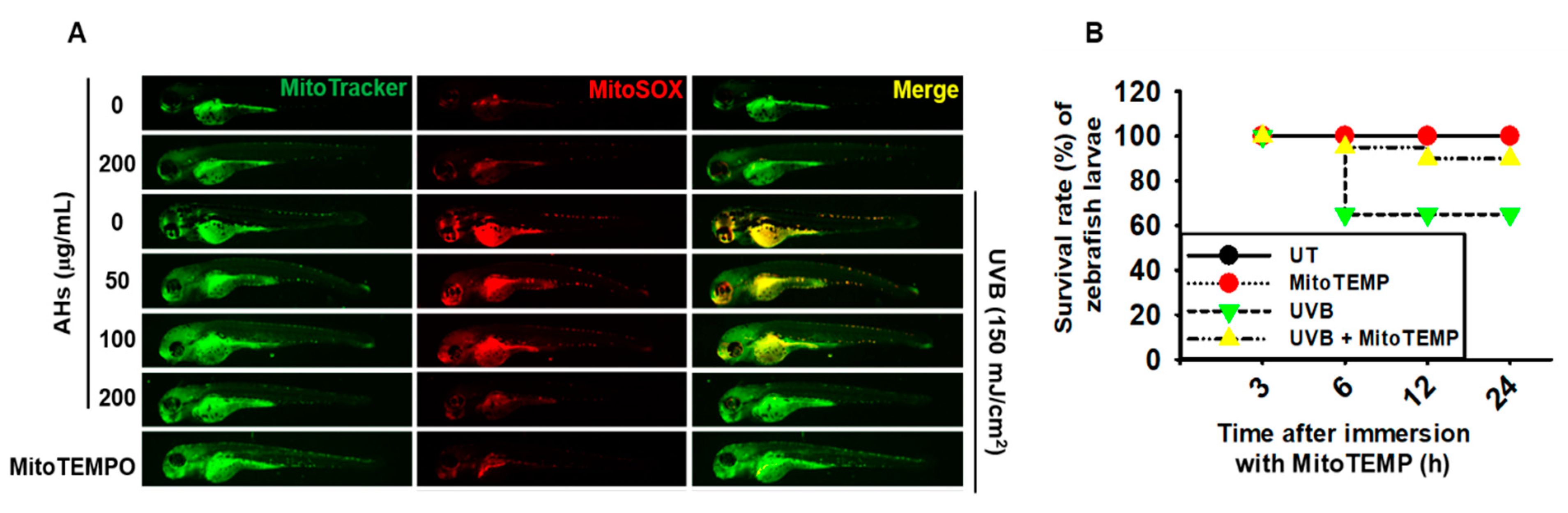
3.7. AHs Reduces mtROS Production in UVB-Irradiated Zebrafish Larvae, Resulting in an Increase in the Survival Rate
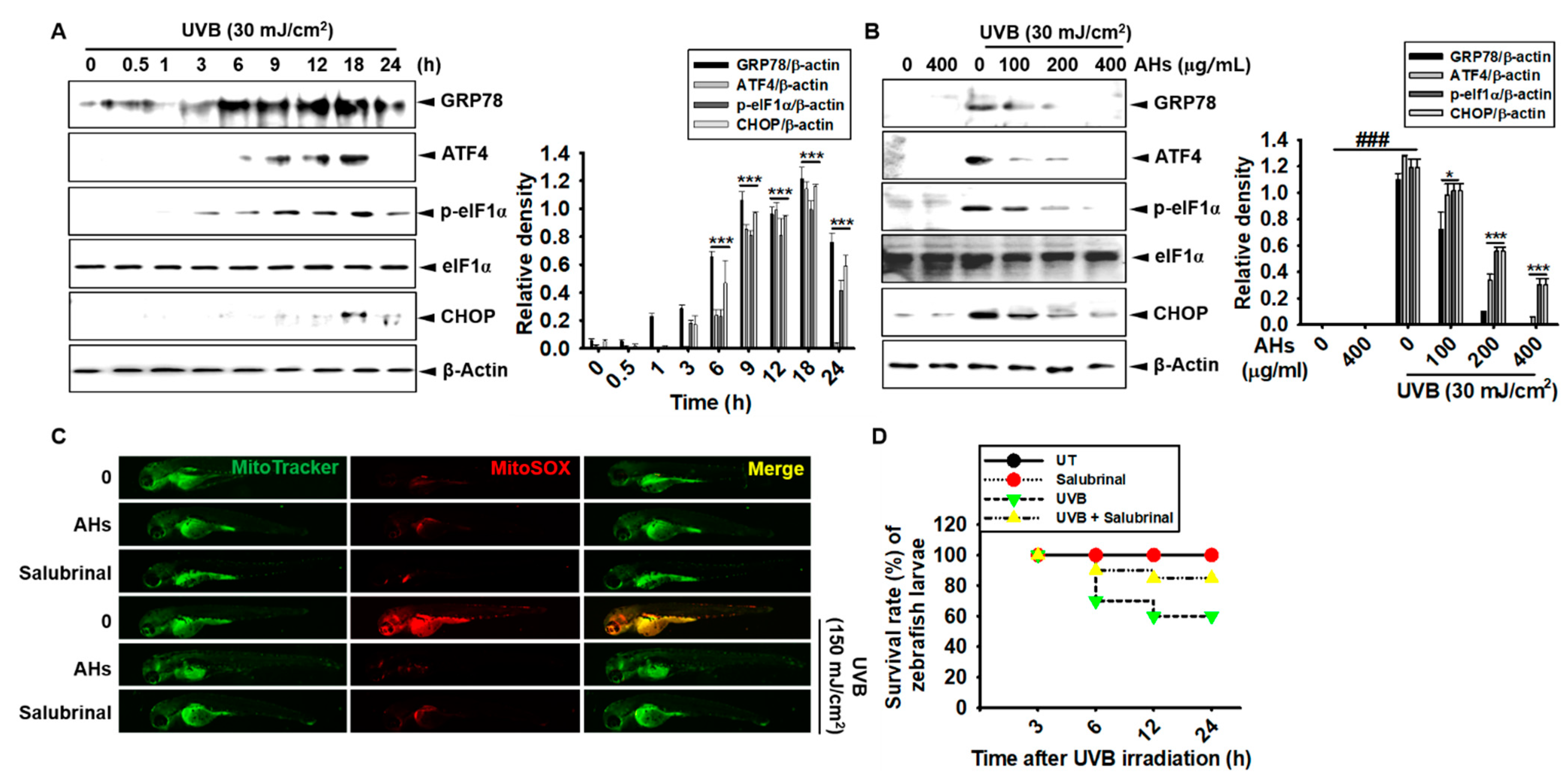
3.8. AHs Inhibits UVB-Induced ER Stress and Consequently Increases the Survival Rate of Zebrafish Larvae through the Inhibition of mtROS Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schuch, A.P.; Moreno, N.C.; Schuch, N.J.; Menck, C.F.M.; Garcia, C.C.M. Sunlight damage to cellular DNA: Focus on oxidatively generated lesions. Free Radic. Biol. Med. 2017, 107, 110–124. [Google Scholar] [CrossRef] [PubMed]
- Wong, D.Y.; Ranganath, T.; Kasko, A.M. Low-dose, long-wave UV light does not affect gene expression of human mesenchymal stem cells. PLoS ONE 2015, 10, e0139307. [Google Scholar] [CrossRef] [Green Version]
- Panich, U.; Sittithumcharee, G.; Rathviboon, N.; Jirawatnotai, S. Ultraviolet radiation-induced skin aging: The role of DNA damage and oxidative stress in epidermal stem cell damage mediated skin aging. Stem Cells Int. 2016, 2016, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Biniek, K.; Levi, K.; Dauskardt, R.H. Solar UV radiation reduces the barrier function of human skin. Proc. Natl. Acad. Sci. USA 2012, 109, 17111–17116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. UV radiation and the skin. Int. J. Mol. Sci. 2013, 14, 12222–12248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, C.E.; Hung, Y.T.; Fang, A.H.; Ching-Shuang, W. Effects of irradiance on UVA-induced skin aging. J. Dermatol. Sci. 2019, 94, 220–228. [Google Scholar] [CrossRef] [Green Version]
- Liebel, F.; Kaur, S.; Ruvolo, E.; Kollias, N.; Southall, M.D. Irradiation of skin with visible light induces reactive oxygen species and matrix-degrading enzymes. J. Investig. Dermatol. 2012, 132, 1901–1907. [Google Scholar] [CrossRef] [Green Version]
- Salucci, S.; Burattini, S.; Curzi, D.; Buontempo, F.; Martelli, A.M.; Zappia, G.; Falcieri, E.; Battistelli, M. Antioxidants in the prevention of UVB-induced keratynocyte apoptosis. J. Photochem. Photobiol. B 2014, 141, 1–9. [Google Scholar] [CrossRef]
- Ellgaard, L.; McCaul, N.; Chatsisvili, A.; Braakman, I. Co- and post-translational protein folding in the ER. Traffic 2016, 17, 615–638. [Google Scholar] [CrossRef] [Green Version]
- Barlowe, C.K.; Miller, E.A. Secretory protein biogenesis and traffic in the early secretory pathway. Genetics 2013, 193, 383–410. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Kaufman, R.J. The impact of the endoplasmic reticulum protein-folding environment on cancer development. Nat. Rev. Cancer 2014, 14, 581–597. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Bailly-Maitre, B.; Reed, J.C. Endoplasmic reticulum stress: Cell life and death decisions. J. Clin. Investig. 2005, 115, 2656–2664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrukh, M.R.; Nissar, U.A.; Afnan, Q.; Rafiq, R.A.; Sharma, L.; Amin, S.; Kaiser, P.; Sharma, P.R.; Tasduq, S.A. Oxidative stress mediated Ca2+ release manifests endoplasmic reticulum stress leading to unfolded protein response in UV-B irradiated human skin cells. J. Dermatol. Sci. 2014, 75, 24–35. [Google Scholar] [CrossRef]
- Yoon, S.W.; Lee, K.P.; Kim, D.Y.; Hwang, D.I.; Won, K.J.; Lee, D.W.; Lee, H.M. Effect of absolute from Hibiscus syriacus L. flower on wound healing in keratinocytes. Pharmacogn. Mag. 2017, 13, 85–89. [Google Scholar] [PubMed]
- Karunarathne, W.; Molagoda, I.M.N.; Park, S.R.; Kim, J.W.; Lee, O.K.; Kwon, H.Y.; Oren, M.; Choi, Y.H.; Ryu, H.W.; Oh, S.R.; et al. Anthocyanins from Hibiscus syriacus L. inhibit melanogenesis by activating the ERK signaling pathway. Biomolecules 2019, 9, 645. [Google Scholar] [CrossRef] [Green Version]
- Karunarathne, W.; Lee, K.T.; Choi, Y.H.; Jin, C.Y.; Kim, G.Y. Anthocyanins isolated from Hibiscus syriacus L. attenuate lipopolysaccharide-induced inflammation and endotoxic shock by inhibiting the TLR4/MD2-mediated NF-kB signaling pathway. Phytomedicine 2020, 76, 153237. [Google Scholar] [CrossRef] [PubMed]
- Molagoda, I.M.N.; Lee, K.T.; Choi, Y.H.; Kim, G.Y. Anthocyanins from Hibiscus syriacus L. inhibit oxidative stress-mediated apoptosis by activating the Nrf2/HO-1 signaling pathway. Antioxidants 2020, 9, 42. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.E.; Park, S.W.; Ngo, H.T.T.; Seo, S.A.; Go, E.B.; Hwang, J.S.; Hwang, E.; Yi, T.H. Skin-protective and anti-inflammatory effects of Hibiscus syriacus L. (Mugunghwa): A comparative study of five parts of the plant. Pharmacog. Mag. 2020, 16, 183–191. [Google Scholar]
- Mohania, D.; Chandel, S.; Kumar, P.; Verma, V.; Digvijay, K.; Tripathi, D.; Choudhury, K.; Mitten, S.K.; Shah, D. Ultraviolet radiations: Skin defense-damage mechanism. Adv. Exp. Med. Biol. 2017, 996, 71–87. [Google Scholar] [PubMed]
- Sander, C.S.; Chang, H.; Hamm, F.; Elsner, P.; Thiele, J.J. Role of oxidative stress and the antioxidant network in cutaneous carcinogenesis. Int. J. Dermatol. 2004, 43, 326–335. [Google Scholar] [CrossRef]
- Salucci, S.; Burattini, S.; Buontempo, F.; Martelli, A.M.; Falcieri, E.; Battistelli, M. Protective effect of different antioxidant agents in UVB-irradiated keratinocytes. Eur. J. Histochem. 2017, 61, 2784. [Google Scholar] [CrossRef]
- Pandel, R.; Poljsak, B.; Godic, A.; Dahmane, R. Skin photoaging and the role of antioxidants in its prevention. Int. Sch. Res. Not. 2013, 2013, 930164. [Google Scholar] [CrossRef]
- Glady, A.; Tanaka, M.; Moniaga, C.S.; Yasui, M.; Hara-Chikuma, M. Involvement of NADPH oxidase 1 in UVB-induced cell signaling and cytotoxicity in human keratinocytes. Biochem. Biophys. Rep. 2018, 14, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Raad, H.; Serrano-Sanchez, M.; Harfouche, G.; Mahfouf, W.; Bortolotto, D.; Bergeron, V.; Kasraian, Z.; Dousset, L.; Hosseini, M.; Taieb, A.; et al. NADPH oxidase-1 plays a key role in keratinocyte responses to UV radiation and UVB-induced skin carcinogenesis. J. Investig. Dermatol. 2017, 137, 1311–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovac, S.; Angelova, P.R.; Holmstrom, K.M.; Zhang, Y.; Dinkova-Kostova, A.T.; Abramov, A.Y. Nrf2 regulates ROS production by mitochondria and NADPH oxidase. Biochim. Biophys. Acta 2015, 1850, 794–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwarz, D.S.; Blower, M.D. The endoplasmic reticulum: Structure, function and response to cellular signaling. Cell. Mol. Life Sci. 2016, 73, 79–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mera, K.; Kawahara, K.; Tada, K.; Kawai, K.; Hashiguchi, T.; Maruyama, I.; Kanekura, T. ER signaling is activated to protect human HaCaT keratinocytes from ER stress induced by environmental doses of UVB. Biochem. Biophys. Res. Commun. 2010, 397, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.; Yang, B.; Huang, S.Y.; Huang, J.W.; Cheng, B. Salubrinal protects human skin fibroblasts against UVB-induced cell death by blocking endoplasmic reticulum (ER) stress and regulating calcium homeostasis. Biochem. Biophys. Res. Commun. 2017, 493, 1371–1376. [Google Scholar] [CrossRef]
- Marchi, S.; Patergnani, S.; Missiroli, S.; Morciano, G.; Rimessi, A.; Wieckowski, M.R.; Giorgi, C.; Pinton, P. Mitochondrial and endoplasmic reticulum calcium homeostasis and cell death. Cell Calcium 2018, 69, 62–72. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karunarathne, W.A.H.M.; Molagoda, I.M.N.; Lee, K.T.; Choi, Y.H.; Yu, S.-M.; Kang, C.-H.; Kim, G.-Y. Protective Effect of Anthocyanin-Enriched Polyphenols from Hibiscus syriacus L. (Malvaceae) against Ultraviolet B-Induced Damage. Antioxidants 2021, 10, 584. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10040584
Karunarathne WAHM, Molagoda IMN, Lee KT, Choi YH, Yu S-M, Kang C-H, Kim G-Y. Protective Effect of Anthocyanin-Enriched Polyphenols from Hibiscus syriacus L. (Malvaceae) against Ultraviolet B-Induced Damage. Antioxidants. 2021; 10(4):584. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10040584
Chicago/Turabian StyleKarunarathne, Wisurumuni Arachchilage Hasitha Maduranga, Ilandarage Menu Neelaka Molagoda, Kyoung Tae Lee, Yung Hyun Choi, Sang-Mi Yu, Chang-Hee Kang, and Gi-Young Kim. 2021. "Protective Effect of Anthocyanin-Enriched Polyphenols from Hibiscus syriacus L. (Malvaceae) against Ultraviolet B-Induced Damage" Antioxidants 10, no. 4: 584. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10040584