
Protective Effects of Maclurin against Benzo[a]pyrene via Aryl Hydrocarbon Receptor and Nuclear Factor Erythroid 2-Related Factor 2 Targeting

 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
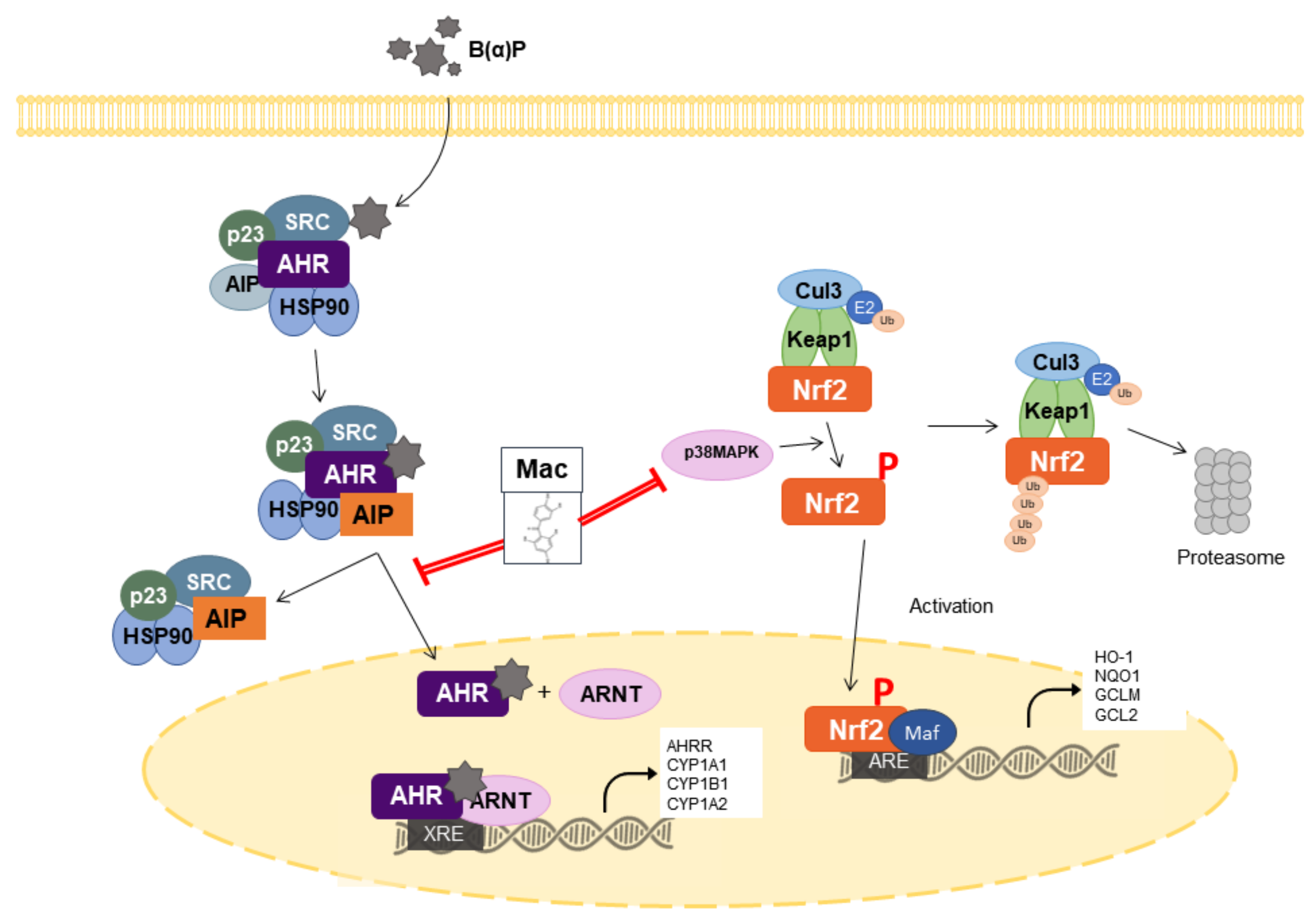{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Culture of Cells
2.3. Assay of Cell Viability Assay
2.4. Measurement of XRE and ARE Activity
2.5. BPDE-DNA Adduct Formation Analysis
2.6. Western Blot Analysis
2.7. DCFDA Cellular Reactive Oxygen Species Detection
2.8. DPPH Radical Scavenging Assay
2.9. ABTS Radical Scavenging Assay
2.10. Immunocytochemistry
2.11. Small-Interfering RNA (siRNA) for Nrf2 and AHR
2.12. Analysis of mRNA Levels Using Real-Time PCR
2.13. Enzyme-Linked Immunosorbent Assay (ELISA)
2.14. Statistical Analysis
3. Results
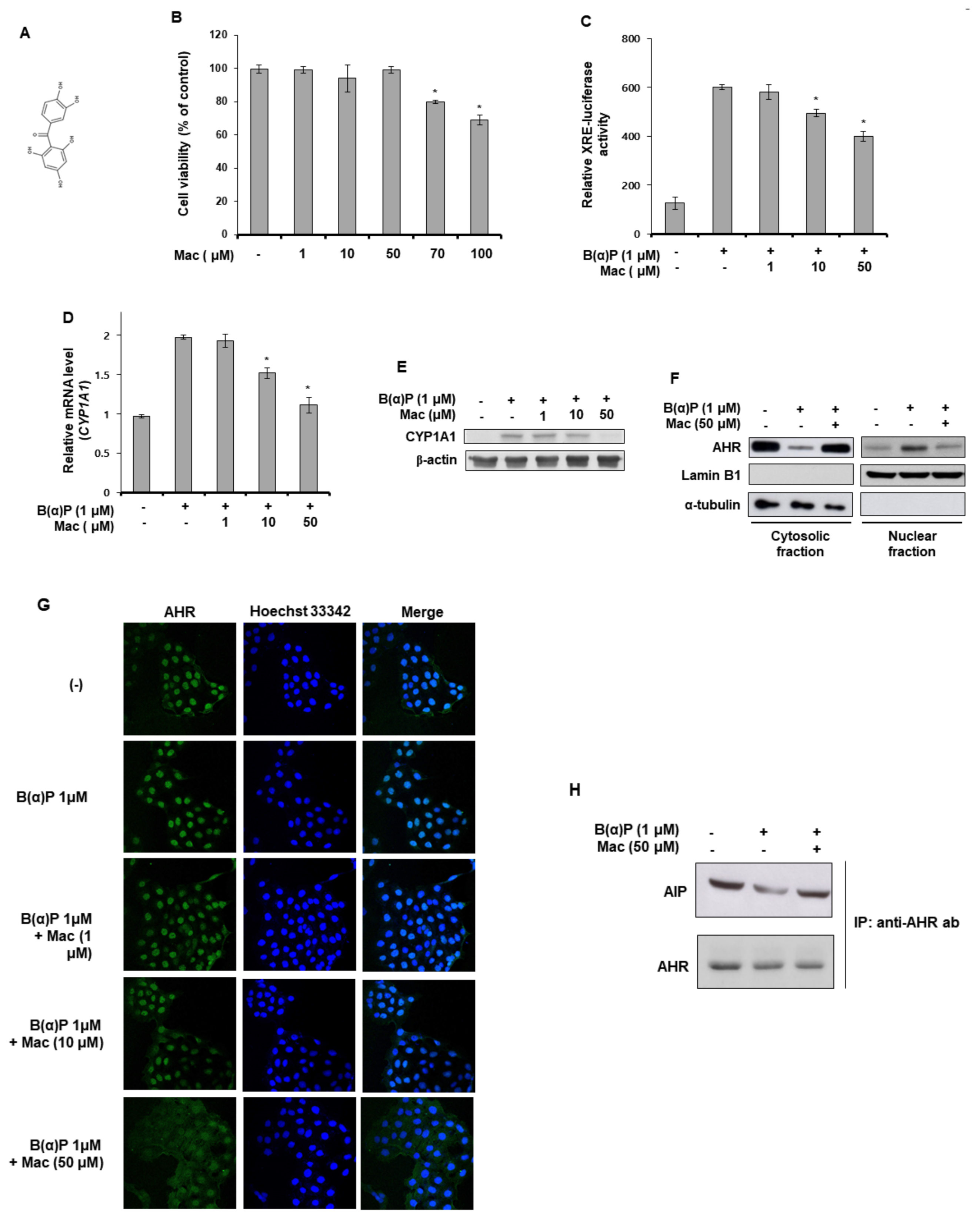
3.1. Maclurin Suppresses B[a]P-Induced AHR Sgnaling and Expression of the CYP1A1 Gene
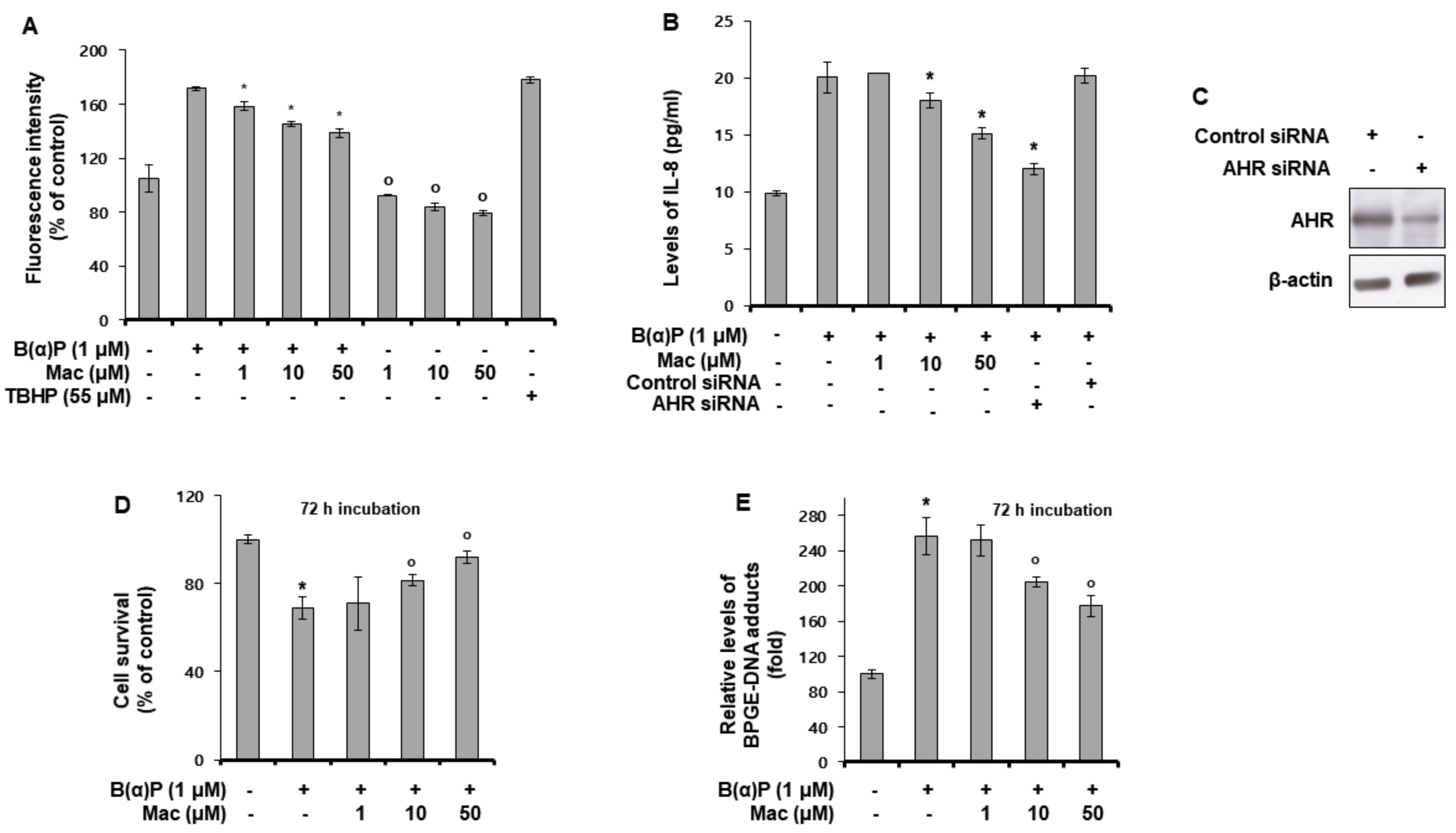
3.2. Maclurin Attenuates B(a)P Effects on the Intracellular Production of ROS and IL-8, Cell Survival, and BPDE-DNA Adduct Formation
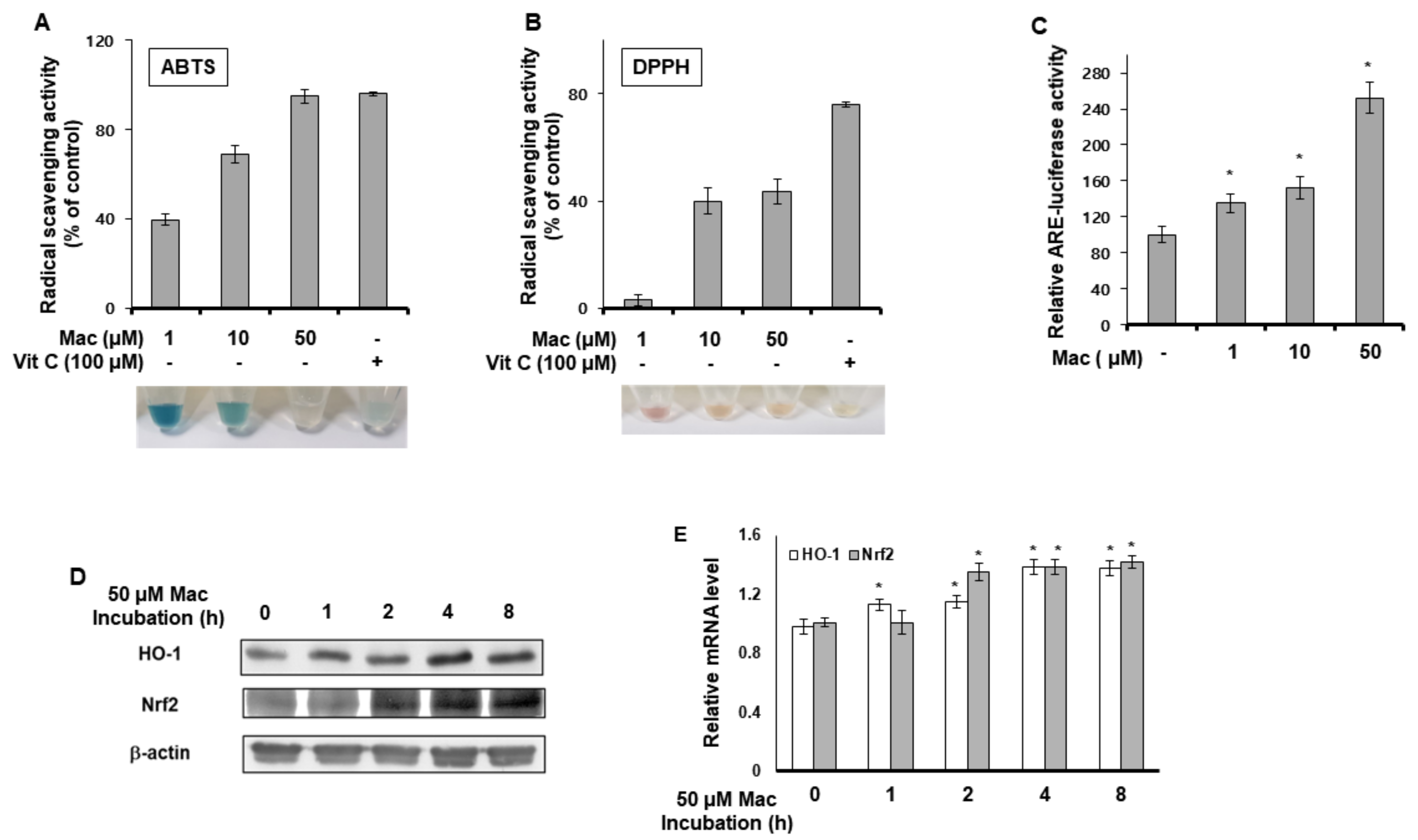
3.3. Maclurin Shows Radical Scavenging Property and Activates ARE Signaling
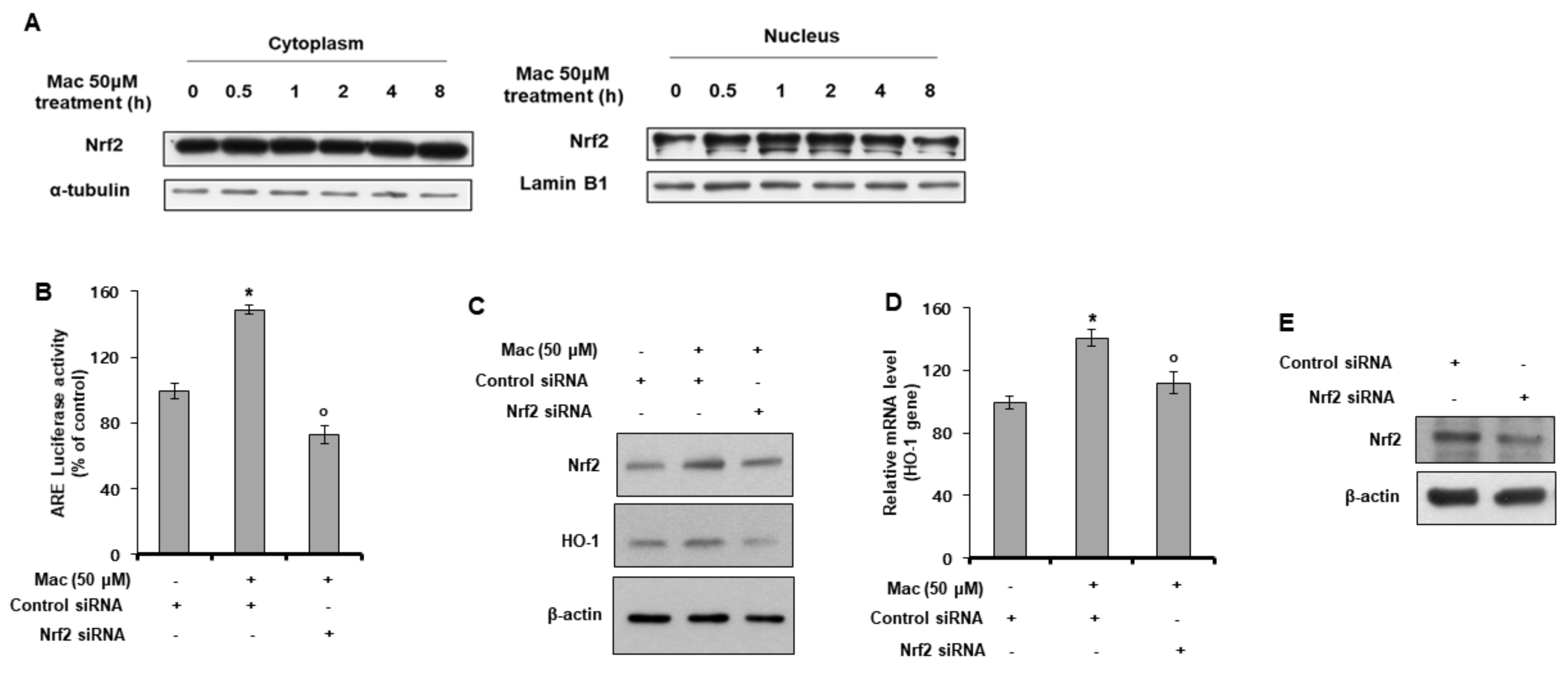
3.4. Maclurin Induces Nuclear Translocation of Nrf2 and Activates Nrf2-Dependent ARE Signaling
3.5. p38 MAPK Activation Mediates Maclurin-Induced Nrf2 Activation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kaiserman, M.J.; Rickert, W.S. Carcinogens in tobacco smoke: Benzo[a]pyrene from Canadian cigarettes and cigarette tobacco. Am. J. Public Health 1992, 82, 1023–1026. [Google Scholar] [CrossRef] [Green Version]
- Larsson, B.K.; Sahlberg, G.P.; Eriksson, A.T.; Busk, L.A. Polycyclic aromatic hydrocarbons in grilled food. J. Agric. Food Chem. 1983, 31, 867–873. [Google Scholar] [CrossRef]
- Courter, L.A.; Pereira, C.; Baird, W.M. Diesel exhaust influences carcinogenic PAH-induced genotoxicity and gene expression in human breast epithelial cells in culture. Mutat. Res. Mol. Mech. Mutagen. 2007, 625, 72–82. [Google Scholar] [CrossRef] [Green Version]
- Ramesh, A.; Inyang, F.; Hood, D.B.; Archibong, A.E.; Knuckles, M.E.; Nyanda, A.M. Metabolism, bioavailability, and toxicokinetics of Benzo(α)pyrenein F-344 rats following oral administration. Exp. Toxicol. Pathol. 2001, 53, 275–290. [Google Scholar] [CrossRef]
- Walle, T.; Walle, U.K.; Sedmera, D.; Klausner, M. Benzo[A]Pyrene-Induced Oral Carcinogenesis and Chemoprevention: Studies in Bioengineered Human Tissue. Drug Metab. Dispos. 2005, 34, 346–350. [Google Scholar] [CrossRef]
- Moody, R.P.; Nadeau, B.; Chu, I. In vivo and in vitro dermal absorption of benzo[a]pyrene in rat, guinea pig, human and tissue-cultured skin. J. Dermatol. Sci. 1995, 9, 48–58. [Google Scholar] [CrossRef]
- Verma, N.; Pink, M.; Rettenmeier, A.W.; Schmitz-Spanke, S. Review on proteomic analyses of benzo[a]pyrene toxicity. Proteomics 2012, 12, 1731–1755. [Google Scholar] [CrossRef] [PubMed]
- Sims, P.; Grover, P.L. Epoxides in polycyclic aromatic hydrocarbon metabolism and carcinogenesis. In Advances in Cancer Research; Klein, G., Weinhouse, S., Haddow, A., Eds.; Academic Press: Cambridge, MA, USA, 1974; pp. 165–274. [Google Scholar]
- Fujii-Kuriyama, Y.; Mimura, J. Molecular mechanisms of AhR functions in the regulation of cytochrome P450 genes. Biochem. Biophys. Res. Commun. 2005, 338, 311–317. [Google Scholar] [CrossRef]
- Hoffman, E.C.; Reyes, H.; Chu, F.F.; Sander, F.; Conley, L.H.; A Brooks, B.; Hankinson, O. Cloning of a factor required for activity of the Ah (dioxin) receptor. Science 1991, 252, 954–958. [Google Scholar] [CrossRef]
- Rothhammer, V.; Quintana, F.J. The aryl hydrocarbon receptor: An environmental sensor integrating immune responses in health and disease. Nat. Rev. Immunol. 2019, 19, 184–197. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.; Nioi, P.; Pickett, C.B. The Nrf2-Antioxidant Response Element Signaling Pathway and Its Activation by Oxidative Stress. J. Biol. Chem. 2009, 284, 13291–13295. [Google Scholar] [CrossRef] [Green Version]
- Gęgotek, A.; Skrzydlewska, E. The role of transcription factor Nrf2 in skin cells metabolism. Arch. Dermatol. Res. 2015, 307, 385–396. [Google Scholar] [CrossRef] [Green Version]
- Motohashi, H.; Yamamoto, M. Nrf2–Keap1 defines a physiologically important stress response mechanism. Trends Mol. Med. 2004, 10, 549–557. [Google Scholar] [CrossRef]
- Itoh, K.; Wakabayashi, N.; Katoh, Y.; Ishii, T.; Igarashi, K.; Engel, J.D.; Yamamoto, M. Keap1 represses nuclear activation of antioxidant responsive elements by Nrf2 through binding to the amino-terminal Neh2 domain. Genes Dev. 1999, 13, 76–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renaud, C.O.; Ziros, P.G.; Chartoumpekis, D.V.; Bongiovanni, M.; Sykiotis, G.P.; Renaud, C.O.; Ziros, P.G.; Chartoumpekis, D.V.; Bongiovanni, M.; Sykiotis, G.P. Keap1/Nrf2 Signaling: A New Player in Thyroid Pathophysiology and Thyroid Cancer. Front. Endocrinol. 2019, 10, 510. [Google Scholar] [CrossRef] [PubMed]
- Knafla, A.; Phillipps, K.; Brecher, R.; Petrovic, S.; Richardson, M. Development of a dermal cancer slope factor for benzo[a]pyrene. Regul. Toxicol. Pharmacol. 2006, 45, 159–168. [Google Scholar] [CrossRef]
- Woo, H.; Lee, J.; Park, D.; Jung, E. Protective Effect of Mulberry (Morus alba L.) Extract against Benzo[a]pyrene Induced Skin Damage through Inhibition of Aryl Hydrocarbon Receptor Signaling. J. Agric. Food Chem. 2017, 65, 10925–10932. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Park, S.-H.; Yoo, J.A.; Kwon, K.; Kim, J.W.; Oh, S.W.; Park, S.J.; Kim, J.; Yu, E.; Han, B.S.; et al. Antagonizing Effects of Clematis apiifolia DC. Extract against Benzo[a]pyrene-Induced Damage to Human Keratinocytes. Oxid. Med. Cell. Longev. 2019, 2019, 2386163. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.P.; Ramos, K.S. Impact Of Cellular Metabolism On The Biological Effects Of Benzo[A]Pyrene And Related Hydrocarbons. Drug Metab. Rev. 2001, 33, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Uchi, H.; Morino-Koga, S.; Nakamura-Satomura, A.; Kita, K.; Shi, W.; Furue, M. Z-Ligustilide inhibits benzo(a)pyrene-induced CYP1A1 upregulation in cultured human keratinocytes via ROS-dependent Nrf2 activation. Exp. Dermatol. 2014, 23, 260–265. [Google Scholar] [CrossRef]
- Uchi, H.; Yasumatsu, M.; Morino-Koga, S.; Mitoma, C.; Furue, M. Inhibition of aryl hydrocarbon receptor signaling and induction of NRF2-mediated antioxidant activity by cinnamaldehyde in human keratinocytes. J. Dermatol. Sci. 2017, 85, 36–43. [Google Scholar] [CrossRef]
- Lee, Y.J.; Jung, O.; Lee, J.; Son, J.; Cho, J.Y.; Ryou, C.; Lee, S.Y. Maclurin exerts anti-cancer effects on PC3 human prostate cancer cells via activation of p38 and inhibitions of JNK, FAK, AKT, and c-Myc signaling pathways. Nutr. Res. 2018, 58, 62–71. [Google Scholar] [CrossRef]
- Li, X.; Gao, Y.; Li, F.; Liang, A.; Xu, Z.; Bai, Y.; Mai, W.; Han, L.; Chen, D. Maclurin protects against hydroxyl radical-induced damages to mesenchymal stem cells: Antioxidant evaluation and mechanistic insight. Chem. Interact. 2014, 219, 221–228. [Google Scholar] [CrossRef]
- Yoo, J.A.; Yu, E.; Park, S.-H.; Oh, S.W.; Kwon, K.; Park, S.J.; Kim, H.; Yang, S.; Park, J.Y.; Cho, J.Y.; et al. Blue Light Irradiation Induces Human Keratinocyte Cell Damage via Transient Receptor Potential Vanilloid 1 (TRPV1) Regulation. Oxid. Med. Cell. Longev. 2020, 2020, 8871745. [Google Scholar] [CrossRef]
- Hwang, Y.S.; Park, S.H.; Kang, M.; Oh, S.W.; Jung, K.; Park, Y.S.; Lee, J. Stemness and differentiation potential-recovery effects of sinapic acid against ultraviolet-A-induced damage through the regulation of p38 MAPK and NF-κB. Sci. Rep. 2017, 7, 909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuyuno, Y.; Uchi, H.; Yasumatsu, M.; Morino-Koga, S.; Tanaka, Y.; Mitoma, C.; Furue, M. Perillaldehyde Inhibits AHR Signaling and Activates NRF2 Antioxidant Pathway in Human Keratinocytes. Oxid. Med. Cell. Longev. 2018, 2018, 9524657. [Google Scholar] [CrossRef]
- Esser, C.; Bargen, I.; Weighardt, H.; Haarmann-Stemmann, T.; Krutmann, J. Functions of the aryl hydrocarbon receptor in the skin. Semin. Immunopathol. 2013, 35, 677–691. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, Z.; Pavek, P. Endogenous and Exogenous Ligands of Aryl Hydrocarbon Receptor: Current State of Art. Curr. Drug Metab. 2011, 12, 198–212. [Google Scholar] [CrossRef] [Green Version]
- Murai, M.; Yamamura, K.; Hashimoto-Hachiya, A.; Tsuji, G.; Furue, M.; Mitoma, C. Tryptophan photo-product FICZ upregulates AHR/MEK/ERK-mediated MMP1 expression: Implications in anti-fibrotic phototherapy. J. Dermatol. Sci. 2018, 91, 97–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manfredi, S.; Federici, C.; Picano, E.; Botto, N.; Rizza, A.; Andreassi, M.G. GSTM1, GSTT1 and CYP1A1 detoxification gene polymorphisms and susceptibility to smoking-related coronary artery disease: A case-only study. Mutat. Res. Mol. Mech. Mutagen. 2007, 621, 106–112. [Google Scholar] [CrossRef]
- Dietrich, C. Antioxidant Functions of the Aryl Hydrocarbon Receptor. Stem Cells Int. 2016, 2016, 7943495. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.C.; Kang, K.A.; Zhang, R.; Piao, M.J.; Kim, G.Y.; Kang, M.Y.; Lee, S.J.; Lee, N.H.; Surh, Y.-J.; Hyun, J.W. Up-regulation of Nrf2-mediated heme oxygenase-1 expression by eckol, a phlorotannin compound, through activation of Erk and PI3K/Akt. Int. J. Biochem. Cell Biol. 2010, 42, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Dong, H.; Song, E.; Xu, X.; Liu, L.; Song, Y. Nrf2/ARE pathway activation, HO-1 and NQO1 induction by polychlorinated biphenyl quinone is associated with reactive oxygen species and PI3K/AKT signaling. Chem. Interact. 2014, 209, 56–67. [Google Scholar] [CrossRef]
- Bellezza, I.; Giambanco, I.; Minelli, A.; Donato, R. Nrf2-Keap1 signaling in oxidative and reductive stress. Biochim. Biophys. Acta (BBA)-Bioenerg. 2018, 1865, 721–733. [Google Scholar] [CrossRef]
- Koyani, C.N.; Kitz, K.; Rossmann, C.; Bernhart, E.; Huber, E.; Trummer, C.; Windischhofer, W.; Sattler, W.; Malle, E. Activation of the MAPK/Akt/Nrf2-Egr1/HO-1-GCLc axis protects MG-63 osteosarcoma cells against 15d-PGJ2-mediated cell death. Biochem. Pharmacol. 2016, 104, 29–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Yuan, X.; Pan, Z.; Shen, G.; Kim, J.H.; Yu, S.; Khor, T.O.; Li, W.; Ma, J.; Kong, A.-N.T. Mechanism of action of isothiocyanates: The induction of ARE-regulated genes is associated with activation of ERK and JNK and the phosphorylation and nuclear translocation of Nrf2. Mol. Cancer Ther. 2006, 5, 1918–1926. [Google Scholar] [CrossRef] [Green Version]
- Velichkova, M.; Hasson, T. Keap1 Regulates the Oxidation-Sensitive Shuttling of Nrf2 into and out of the Nucleus via a Crm1-Dependent Nuclear Export Mechanism. Mol. Cell. Biol. 2005, 25, 4501–4513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuji, G.; Takahara, M.; Uchi, H.; Takeuchi, S.; Mitoma, C.; Moroi, Y.; Furue, M. An environmental contaminant, benzo(a)pyrene, induces oxidative stress-mediated interleukin-8 production in human keratinocytes via the aryl hydrocarbon receptor signaling pathway. J. Dermatol. Sci. 2011, 62, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Uno, S.; Dalton, T.P.; Shertzer, H.G.; Genter, M.B.; Warshawsky, D.; Talaska, G.; Nebert, D.W. Benzo[a]pyrene-Induced Toxicity: Paradoxical Protection in Cyp1a1(−/−) Knockout Mice Having Increased Hepatic BaP–DNA Adduct Levels. Biochem. Biophys. Res. Commun. 2001, 289, 1049–1056. [Google Scholar] [CrossRef]
- Kawachi, Y.; Xu, X.; Taguchi, S.; Sakurai, H.; Nakamura, Y.; Ishii, Y.; Fujisawa, Y.; Furuta, J.; Takahashi, T.; Itoh, K.; et al. Attenuation of UVB-Induced Sunburn Reaction and Oxidative DNA Damage with no Alterations in UVB-Induced Skin Carcinogenesis in Nrf2 Gene-Deficient Mice. J. Investig. Dermatol. 2008, 128, 1773–1779. [Google Scholar] [CrossRef] [Green Version]
- Choi, C.Y.; Kim, J.Y.; Wee, S.Y.; Lee, J.H.; Nam, D.H.; Kim, C.H.; Cho, M.K.; Lee, Y.J.; Nam, H.S.; Lee, S.H.; et al. Expression of Nuclear Factor Erythroid 2 Protein in Malignant Cutaneous Tumors. Arch. Plast. Surg. 2014, 41, 654–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerins, M.J.; Ooi, A. A catalogue of somatic NRF2 gain-of-function mutations in cancer. Sci. Rep. 2018, 8, 12846. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.; Park, S.-H.; Yang, S.; Oh, S.W.; Kwon, K.; Park, S.J.; Yu, E.; Kim, H.; Park, J.Y.; Choi, S.; et al. Protective Effects of Maclurin against Benzo[a]pyrene via Aryl Hydrocarbon Receptor and Nuclear Factor Erythroid 2-Related Factor 2 Targeting. Antioxidants 2021, 10, 1189. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10081189
Kim J, Park S-H, Yang S, Oh SW, Kwon K, Park SJ, Yu E, Kim H, Park JY, Choi S, et al. Protective Effects of Maclurin against Benzo[a]pyrene via Aryl Hydrocarbon Receptor and Nuclear Factor Erythroid 2-Related Factor 2 Targeting. Antioxidants. 2021; 10(8):1189. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10081189
Chicago/Turabian StyleKim, Jangsoon, See-Hyoung Park, Seyoung Yang, Sae Woong Oh, Kitae Kwon, Se Jung Park, Eunbi Yu, Hyeyoun Kim, Jung Yoen Park, Seoyoung Choi, and et al. 2021. "Protective Effects of Maclurin against Benzo[a]pyrene via Aryl Hydrocarbon Receptor and Nuclear Factor Erythroid 2-Related Factor 2 Targeting" Antioxidants 10, no. 8: 1189. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox10081189