
Upregulation of Thioredoxin Reductase 1 Expression by Flavan-3-Ols Protects Human Kidney Proximal Tubular Cells from Hypoxia-Induced Cell Death

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures and Reagents
2.2. Induction of Hypoxia In Vitro
2.3. Flow Cytometric Determination of Cell Death, Reactive Oxygen Species, and Mitochondrial Membrane Potential
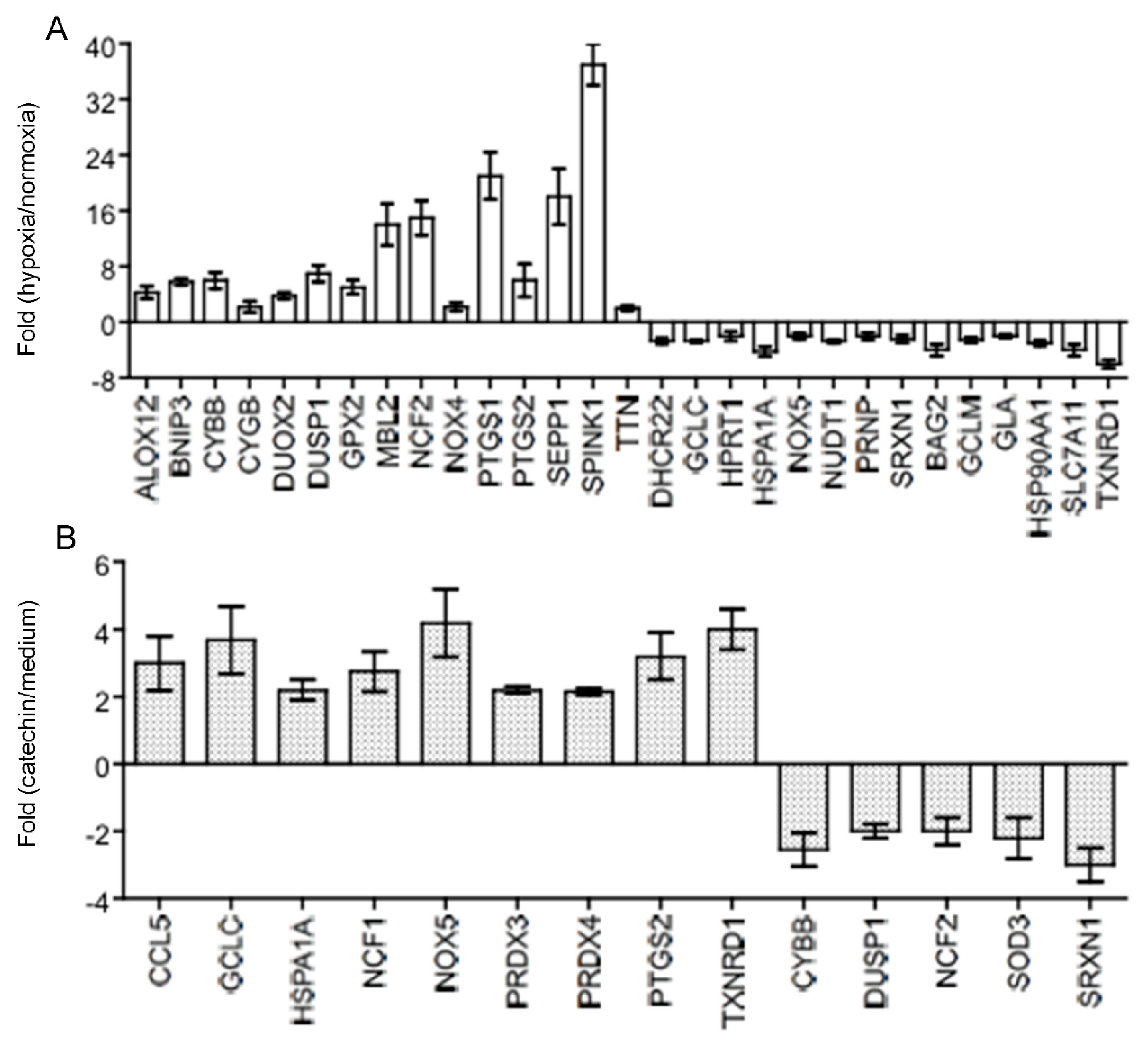
2.4. PCR Array Analysis of Oxidative Stress-Related Gene Expression
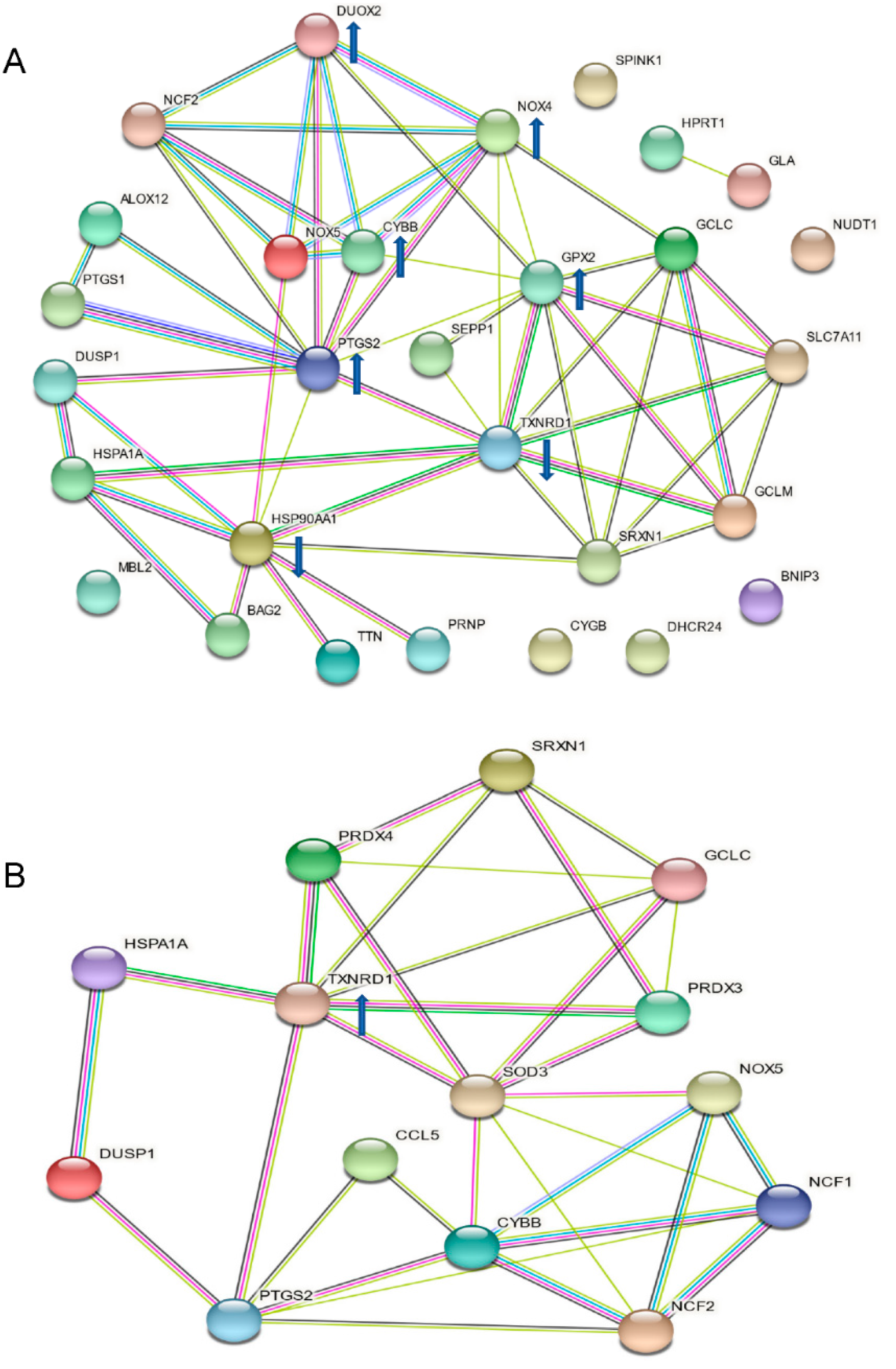
2.5. Analysis of Protein-Protein Interaction and Functional Association Networks
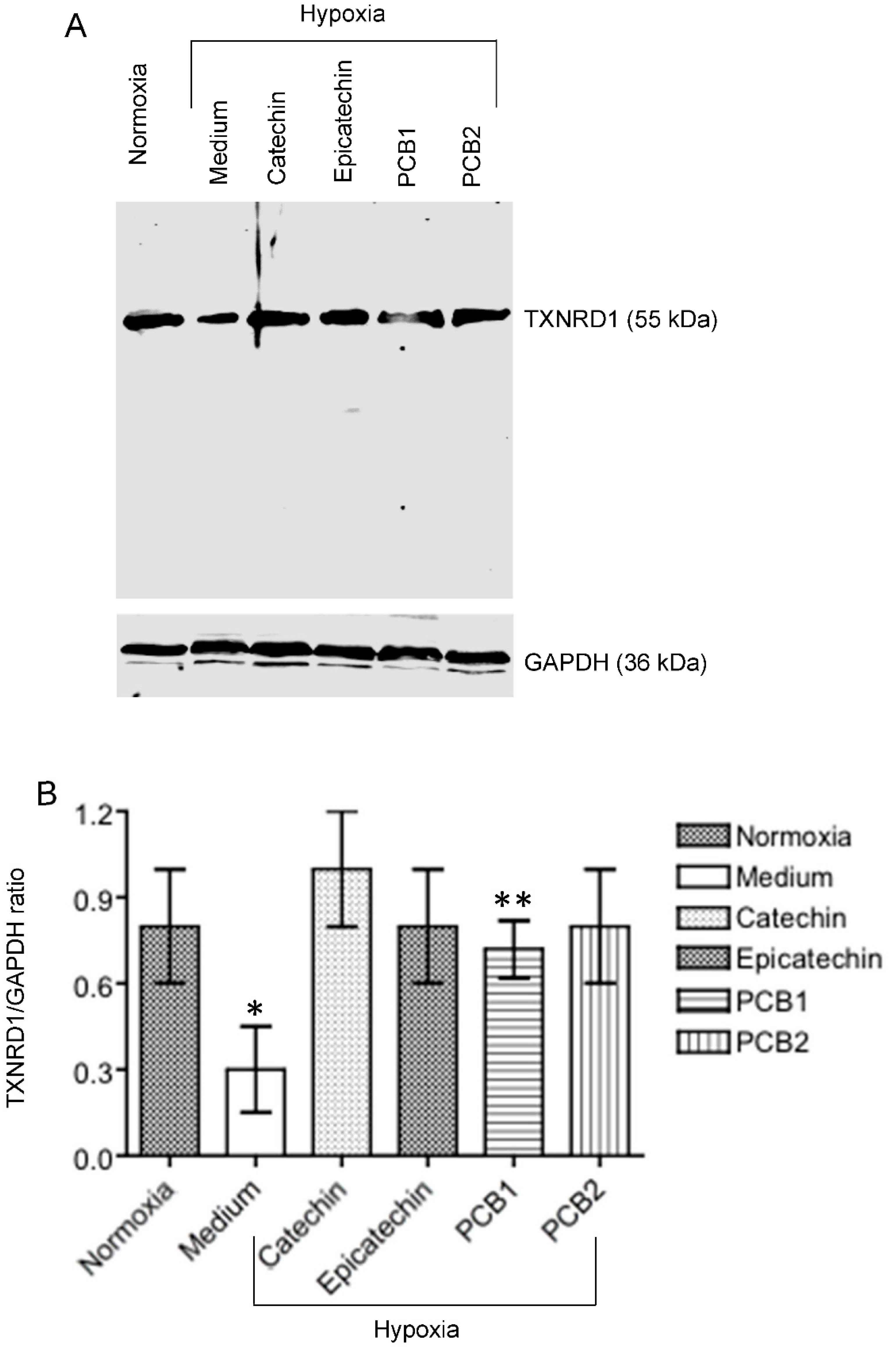
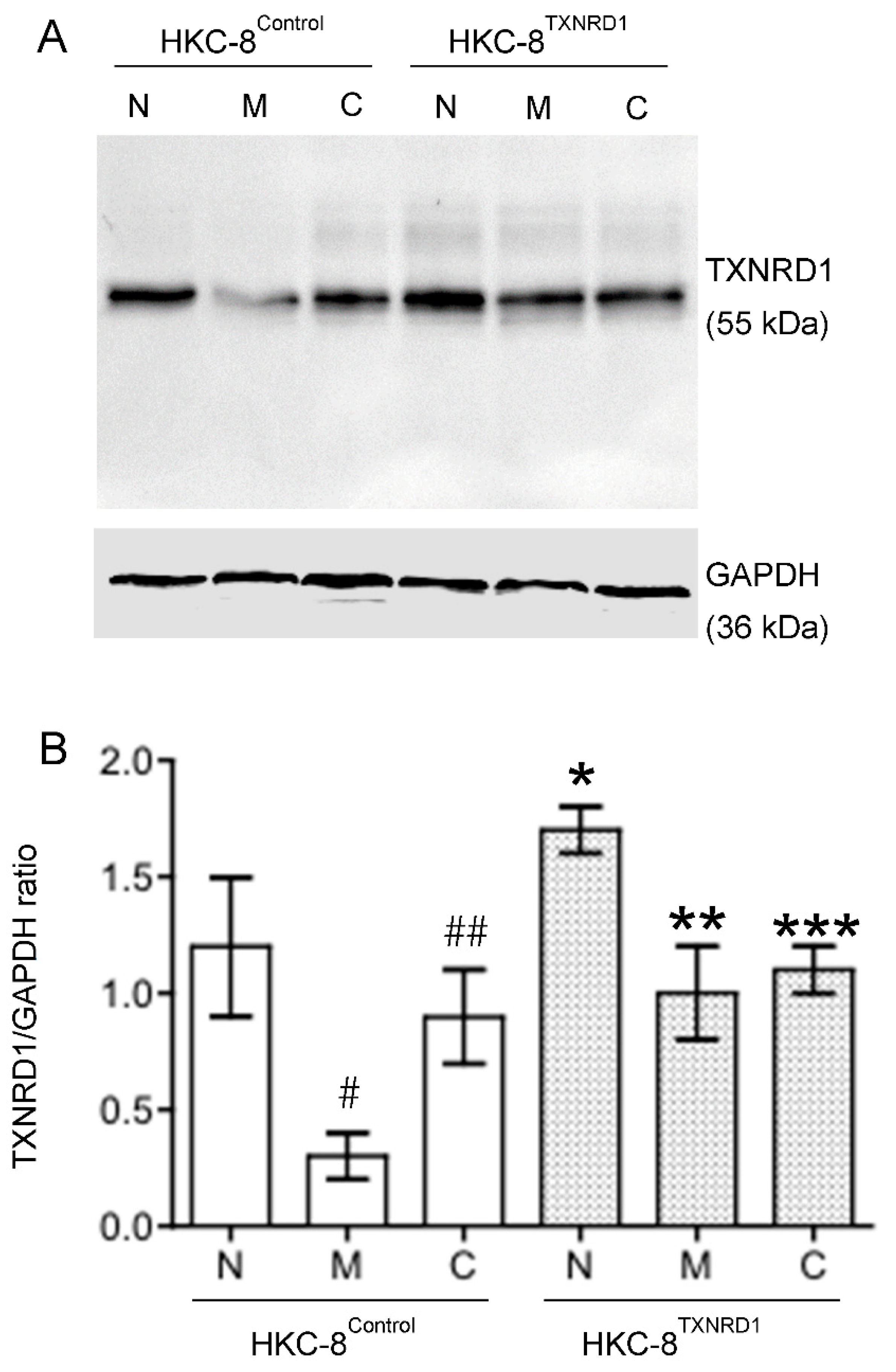
2.6. Western Blot Analysis
2.7. Statistical Analysis
3. Results
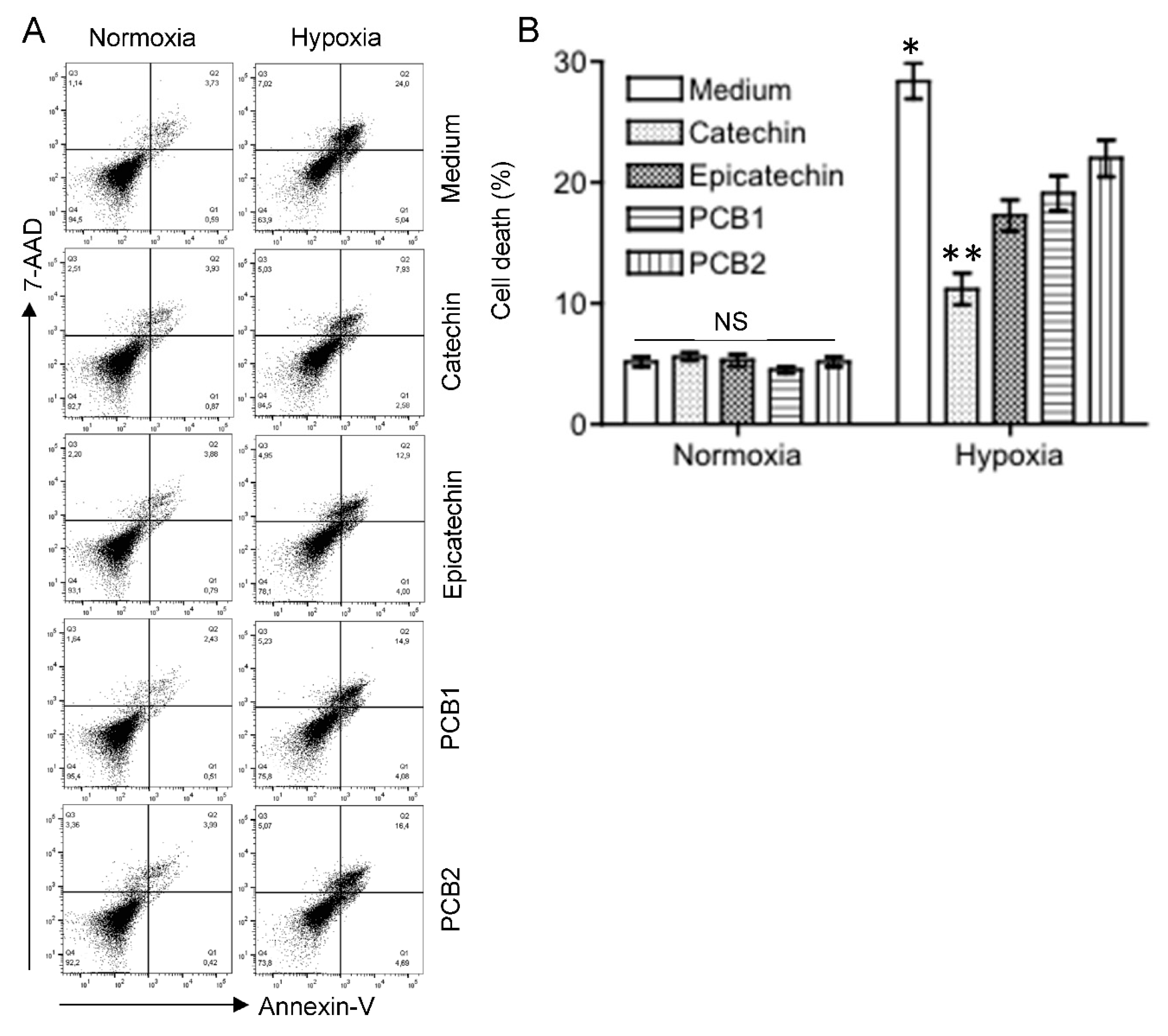
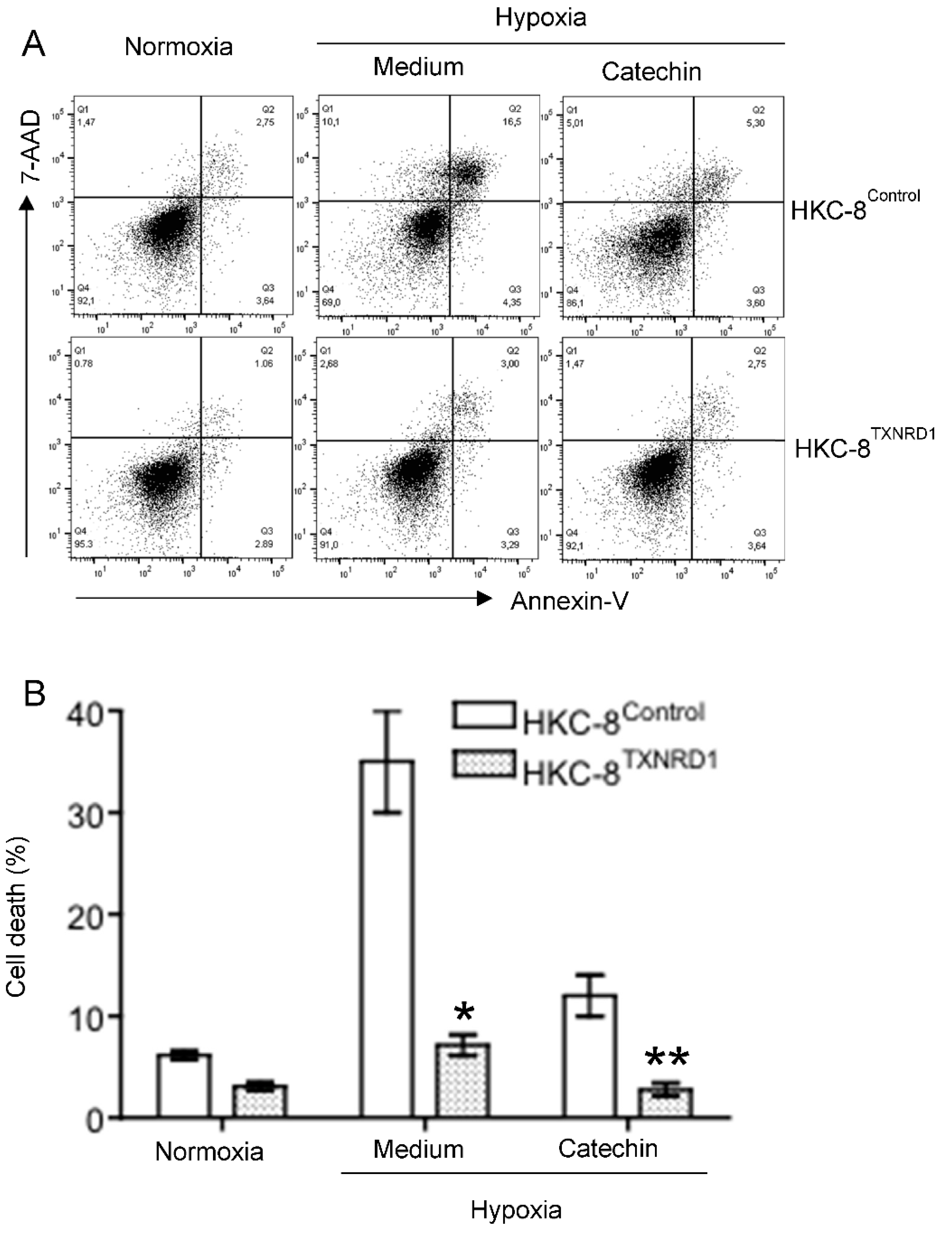
3.1. Reduction of Cell Death by Grape-Derived Flavan-3-Ols in Hypoxic HKC-8 Cells
3.2. Up-Regulated TXNRD 1, a Main Node of the Catechin-Activated Network in Hypoxic HKC-8 Cells
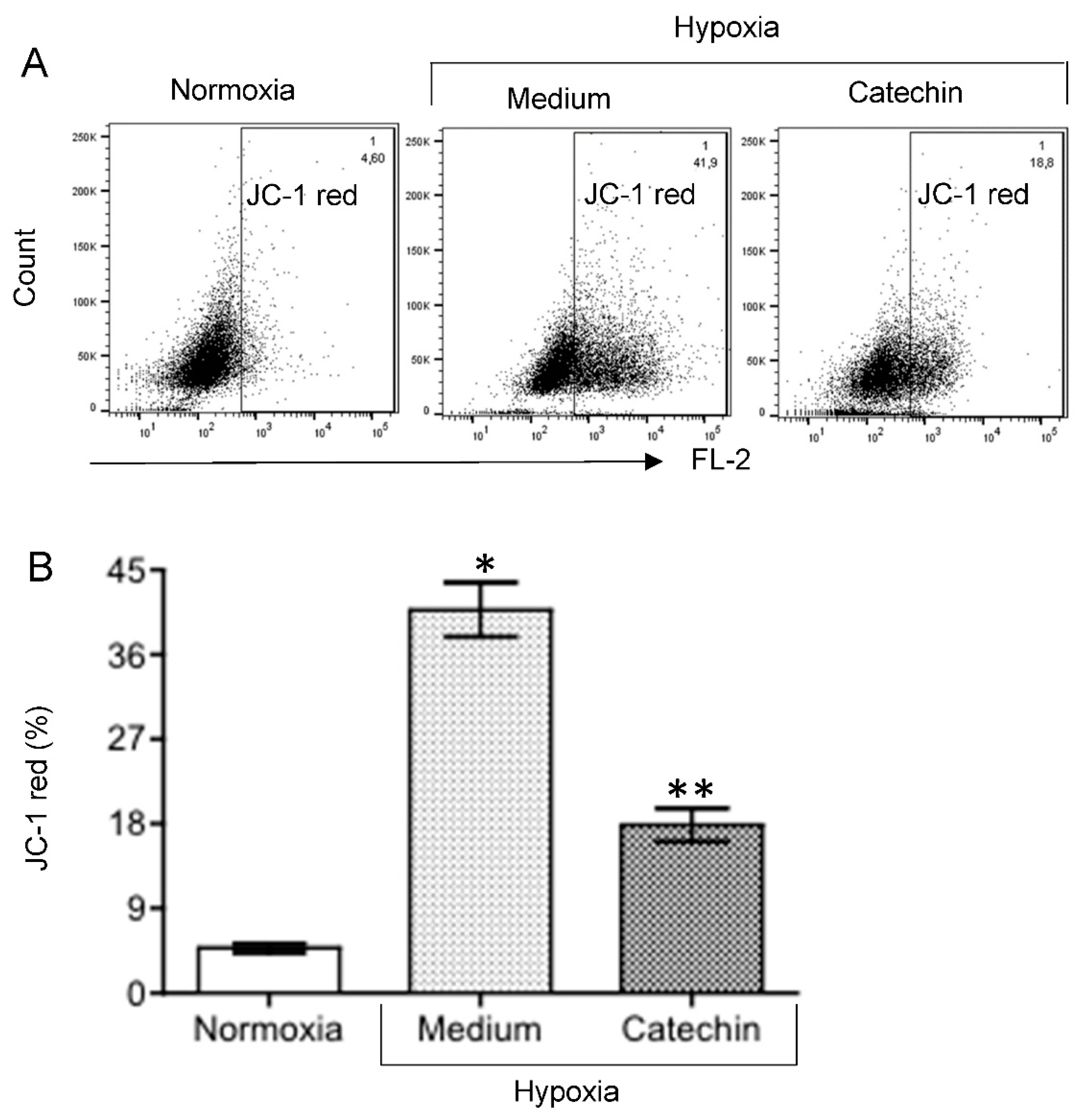
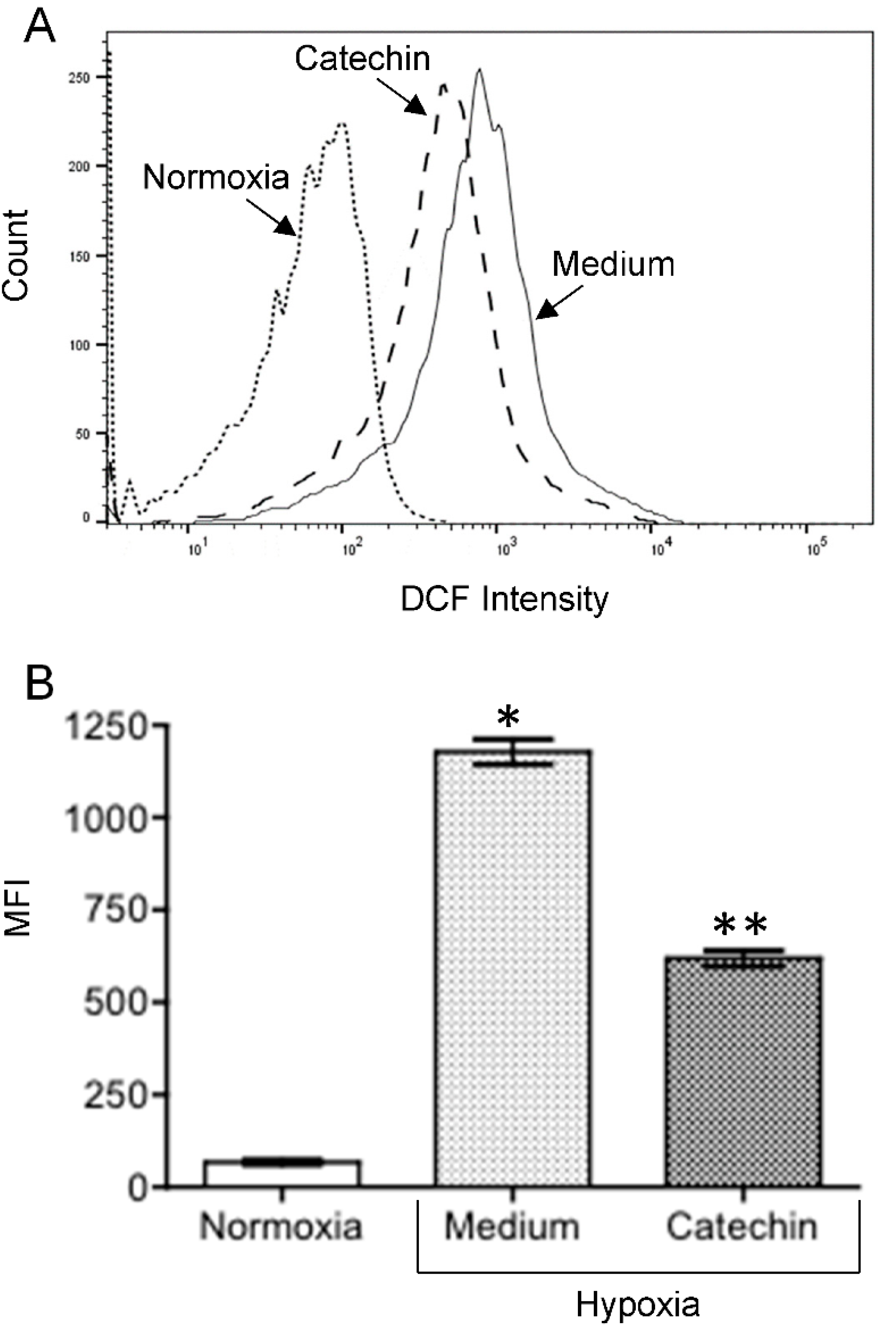
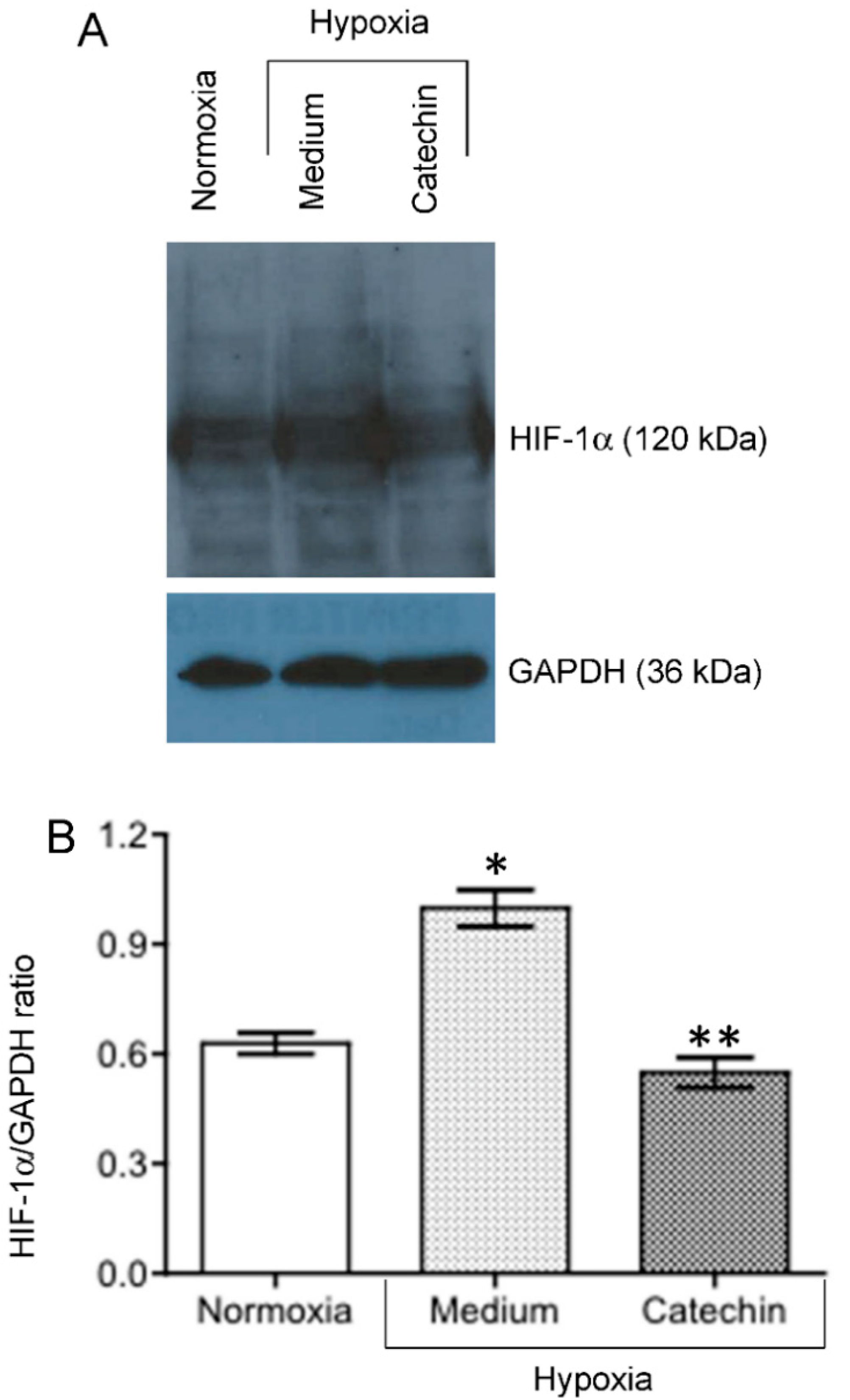
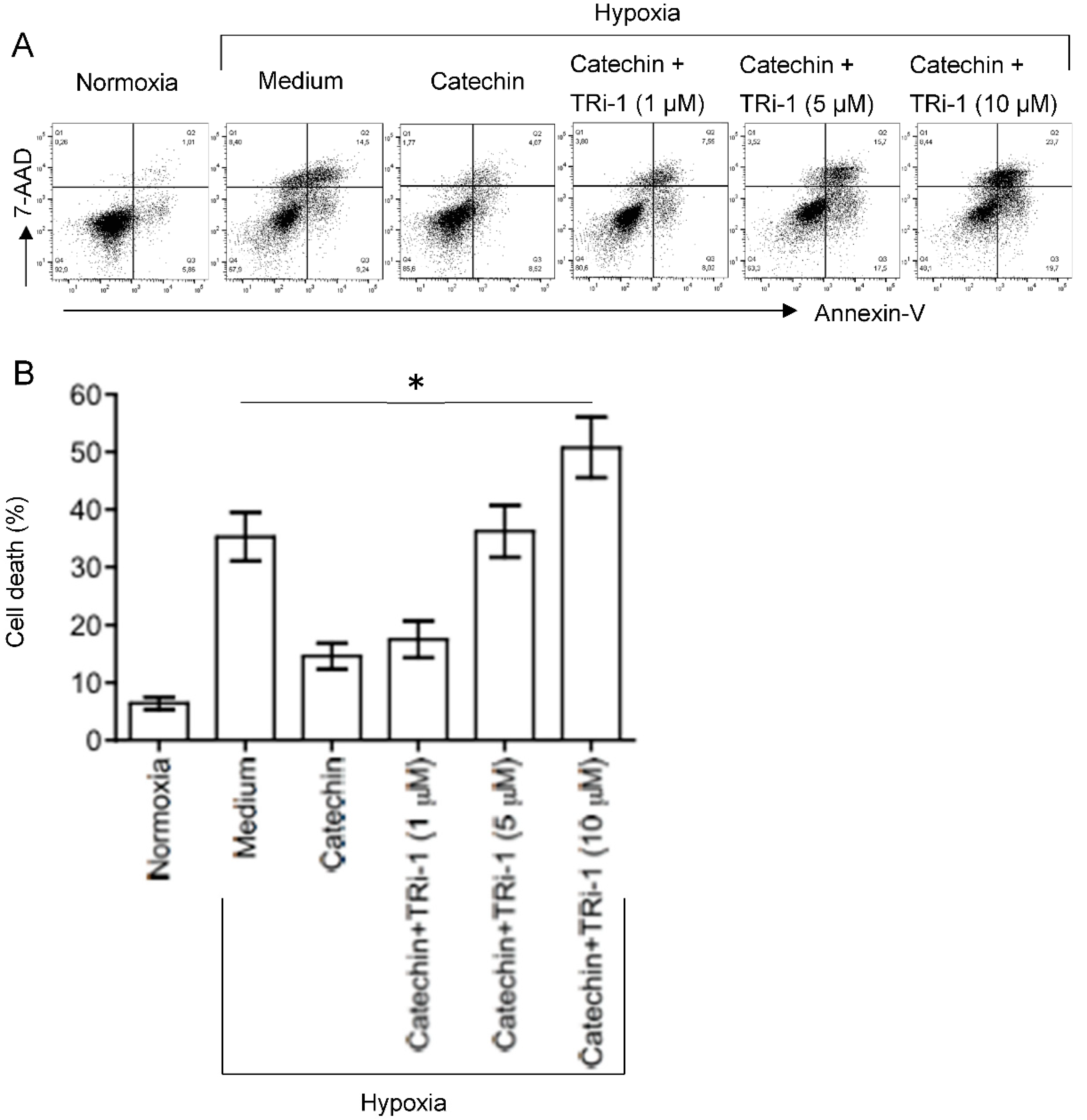
3.3. Up-Regulated TXNRD1, a Cytoprotective Protein Mediating Cytoprotection of Catechin against Cell Death in Hypoxic HKC-8 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nangaku, M. Chronic hypoxia and tubulointerstitial injury: A final common pathway to end-stage renal failure. J. Am. Soc. Nephrol. 2006, 17, 17–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Q.; Colgan, S.P.; Shelley, C.S. Hypoxia: The Force that drives chronic kidney disease. Clin. Med. Res. 2016, 14, 15–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honda, T.; Hirakawa, Y.; Nangaku, M. The role of oxidative stress and hypoxia in renal disease. Kidney Res. Clin. Pract. 2019, 38, 414–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faivre, A.; Scholz, C.C.; de Seigneux, S. Hypoxia in chronic kidney disease: Towards a paradigm shift? Nephrol. Dial. Transplant. 2021, 36, 1782–1790. [Google Scholar] [CrossRef]
- Cook, N.C.; Samman, S. Flavonoids—Chemistry, metabolism, cardioprotective effects, and dietary sources. J. Nutr. Biochem. 1996, 7, 66–76. [Google Scholar] [CrossRef]
- Yao, L.H.; Jiang, Y.M.; Shi, J.; Tomas-Barberan, F.A.; Datta, N.; Singanusong, R.; Chen, S.S. Flavonoids in food and their health benefits. Plant Food Hum. Nutr. 2004, 59, 113–122. [Google Scholar] [CrossRef]
- Mena, P.; Dominguez-Perles, R.; Girones-Vilaplana, A.; Baenas, N.; Garcia-Viguera, C.; Villano, D. Flavan-3-ols, anthocyanins, and inflammation. IUBMB Life 2014, 66, 745–758. [Google Scholar] [CrossRef]
- Oliveira, J.; da Silva, M.A.; Parola, A.J.; Mateus, N.; Bras, N.F.; Ramos, M.J.; de Freitas, V. Structural characterization of a A-type linked trimeric anthocyanin derived pigment occurring in a young Port wine. Food Chem. 2013, 141, 1987–1996. [Google Scholar] [CrossRef]
- Zhu, J.; Du, C. Could grape-based food supplements prevent the development of chronic kidney disease? Crit. Rev. Food Sci. Nutr. 2020, 60, 3054–3062. [Google Scholar] [CrossRef]
- Singh, D.; Chander, V.; Chopra, K. Protective effect of catechin on ischemia-reperfusion-induced renal injury in rats. Pharmacol. Rep. 2005, 57, 70–76. [Google Scholar]
- Ashtiyani, S.C.; Najafi, H.; Firouzifar, M.R.; Shafaat, O. Grape seed extract for reduction of renal disturbances following reperfusion in rats. Iran. J. Kidney Dis. 2013, 7, 28–35. [Google Scholar] [PubMed]
- Ohkita, M.; Hayashi, H.; Ito, K.; Shigematsu, N.; Tanaka, R.; Tsutsui, H.; Matsumura, Y. Preventive effects of grape extract on ischemia/reperfusion-induced acute kidney injury in mice. Biol. Pharm. Bull. 2019, 42, 1883–1890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almomen, S.M.; Guan, Q.; Liang, P.; Yang, K.; Sidiqi, A.M.; Levin, A.; Du, C. Daily intake of grape powder prevents the progression of kidney disease in obese Type 2 diabetic ZSF1 rats. Nutrients 2017, 9, 345. [Google Scholar] [CrossRef] [Green Version]
- Turki, K.; Charradi, K.; Boukhalfa, H.; Belhaj, M.; Limam, F.; Aouani, E. Grape seed powder improves renal failure of chronic kidney disease patients. EXCLI J. 2016, 15, 424–433. [Google Scholar] [CrossRef]
- Racusen, L.C.; Monteil, C.; Sgrignoli, A.; Lucskay, M.; Marouillat, S.; Rhim, J.G.; Morin, J.P. Cell lines with extended in vitro growth potential from human renal proximal tubule: Characterization, response to inducers, and comparison with established cell lines. J. Lab. Clin. Med. 1997, 129, 318–329. [Google Scholar] [CrossRef]
- Du, C.; Guan, Q.; Diao, H.; Yin, Z.; Jevnikar, A.M. Nitric oxide induces apoptosis in renal tubular epithelial cells through activation of caspase-8. Am. J. Physiol.-Renal Physiol. 2006, 290, F1044–F1054. [Google Scholar] [CrossRef]
- Chang, H.; Huang, H.; Huang, T.; Yang, P.; Wang, Y.; Juan, H. Flow cytometric detection of reactive oxygen species. Bio-protocol 2013, 3, e431. [Google Scholar] [CrossRef] [Green Version]
- Sivandzade, F.; Bhalerao, A.; Cucullo, L. Analysis of the mitochondrial membrane potential using the cationic JC-1 dye as a sensitive fluorescent probe. Bio-protocol 2019, 9, e3128. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Weinberg, J.M.; Venkatachalam, M.A.; Roeser, N.F.; Nissim, I. Mitochondrial dysfunction during hypoxia/reoxygenation and its correction by anaerobic metabolism of citric acid cycle intermediates. Proc. Natl. Acad. Sci. USA 2000, 97, 2826–2831. [Google Scholar] [CrossRef] [Green Version]
- Schiffer, T.A.; Friederich-Persson, M. Mitochondrial reactive oxygen species and kidney hypoxia in the development of diabetic nephropathy. Front. Physiol. 2017, 8, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial reactive oxygen species (ROS) and ROS-induced ROS release. Physiol. Rev. 2014, 94, 909–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberger, C.; Mandriota, S.; Jurgensen, J.S.; Wiesener, M.S.; Horstrup, J.H.; Frei, U.; Ratcliffe, P.J.; Maxwell, P.H.; Bachmann, S.; Eckardt, K.U. Expression of hypoxia-inducible factor-1α and -2α in hypoxic and ischemic rat kidneys. J. Am. Soc. Nephrol. 2002, 13, 1721–1732. [Google Scholar] [CrossRef] [Green Version]
- Wiesener, M.S.; Jurgensen, J.S.; Rosenberger, C.; Scholze, C.K.; Horstrup, J.H.; Warnecke, C.; Mandriota, S.; Bechmann, I.; Frei, U.A.; Pugh, C.W.; et al. Widespread hypoxia-inducible expression of HIF-2alpha in distinct cell populations of different organs. FASEB J. 2003, 17, 271–273. [Google Scholar] [CrossRef] [Green Version]
- Sabatier, P.; Beusch, C.M.; Gencheva, R.; Cheng, Q.; Zubarev, R.; Arner, E.S.J. Comprehensive chemical proteomics analyses reveal that the new TRi-1 and TRi-2 compounds are more specific thioredoxin reductase 1 inhibitors than auranofin. Redox Biol. 2021, 48, 102184. [Google Scholar] [CrossRef]
- Stafford, W.C.; Peng, X.; Olofsson, M.H.; Zhang, X.; Luci, D.K.; Lu, L.; Cheng, Q.; Tresaugues, L.; Dexheimer, T.S.; Coussens, N.P.; et al. Irreversible inhibition of cytosolic thioredoxin reductase 1 as a mechanistic basis for anticancer therapy. Sci. Transl. Med. 2018, 10, eaaf7444. [Google Scholar] [CrossRef] [Green Version]
- Halliwell, B. Oxidative stress and neurodegeneration: Where are we now? J. Neurochem. 2006, 97, 1634–1658. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Wang, X.; Chen, H. Oxidative stress to renal tubular epithelial cells—A common pathway in renal pathologies. In Systems Biology of Free Radicals and Oxidants; Laher, I., Ed.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 2605–2624. [Google Scholar] [CrossRef]
- Liu, Y.; Fiskum, G.; Schubert, D. Generation of reactive oxygen species by the mitochondrial electron transport chain. J. Neurochem. 2002, 80, 780–787. [Google Scholar] [CrossRef]
- Hernandez-Garcia, D.; Wood, C.D.; Castro-Obregon, S.; Covarrubias, L. Reactive oxygen species: A radical role in development? Free Radic. Biol. Med. 2010, 49, 130–143. [Google Scholar] [CrossRef]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Ryter, S.W.; Kim, H.P.; Hoetzel, A.; Park, J.W.; Nakahira, K.; Wang, X.; Choi, A.M. Mechanisms of cell death in oxidative stress. Antioxid. Redox Signal. 2007, 9, 49–89. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; McMillan-Ward, E.; Kong, J.; Israels, S.J.; Gibson, S.B. Oxidative stress induces autophagic cell death independent of apoptosis in transformed and cancer cells. Cell Death Differ. 2008, 15, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Bleier, L.; Drose, S. Superoxide generation by complex III: From mechanistic rationales to functional consequences. Biochim. Biophys. Acta 2013, 1827, 1320–1331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuhrmann, D.C.; Brune, B. Mitochondrial composition and function under the control of hypoxia. Redox. Biol. 2017, 12, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Kierans, S.J.; Taylor, C.T. Regulation of glycolysis by the hypoxia-inducible factor (HIF): Implications for cellular physiology. J. Physiol. 2021, 599, 23–37. [Google Scholar] [CrossRef] [PubMed]
- Greijer, A.E.; van der Wall, E. The role of hypoxia inducible factor 1 (HIF-1) in hypoxia induced apoptosis. J. Clin. Pathol. 2004, 57, 1009–1014. [Google Scholar] [CrossRef]
- Cimmino, F.; Avitabile, M.; Lasorsa, V.A.; Montella, A.; Pezone, L.; Cantalupo, S.; Visconte, F.; Corrias, M.V.; Iolascon, A.; Capasso, M. HIF-1 transcription activity: HIF1A driven response in normoxia and in hypoxia. BMC Med. Genet. 2019, 20, 37. [Google Scholar] [CrossRef] [Green Version]
- Bartoszewski, R.; Moszynska, A.; Serocki, M.; Cabaj, A.; Polten, A.; Ochocka, R.; Dell’Italia, L.; Bartoszewska, S.; Kroliczewski, J.; Dabrowski, M.; et al. Primary endothelial cell-specific regulation of hypoxia-inducible factor (HIF)-1 and HIF-2 and their target gene expression profiles during hypoxia. FASEB J. 2019, 33, 7929–7941. [Google Scholar] [CrossRef]
- Weidemann, A.; Johnson, R.S. Biology of HIF-1α. Cell Death Differ. 2008, 15, 621–627. [Google Scholar] [CrossRef] [Green Version]
- Yevshin, I.; Sharipov, R.; Valeev, T.; Kel, A.; Kolpakov, F. GTRD: A database of transcription factor binding sites identified by ChIP-seq experiments. Nucleic Acids Res. 2017, 45, D61–D67. [Google Scholar] [CrossRef]
- Bernatoniene, J.; Kopustinskiene, D.M. The Role of Catechins in Cellular Responses to Oxidative Stress. Molecules 2018, 23, 965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yilmaz, Y.; Toledo, R.T. Major flavonoids in grape seeds and skins: Antioxidant capacity of catechin, epicatechin, and gallic acid. J. Agric. Food Chem. 2004, 52, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Iacopini, P.; Baldi, M.; Storchi, P.; Sebastiani, L. Catechin, epicatechin, quercetin, rutin and resveratrol in red grape: Content, in vitro antioxidant activity and interactions. J. Food Compos. Anal. 2008, 21, 589–598. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GO-Term | Description of Functions | Count in Network | Strength | False Discovery Rate |
|---|---|---|---|---|
| 0004666 | Prostaglandin-endoperoxide synthase activity | 2 (PTGS1, PTGS2) of 2 | 2.83 | 0.0032 |
| 0004357 | Glutamate-cysteine ligase activity | 2 (GCLC, GCLM) of 2 | 2.83 | 0.0032 |
| 0016175 | Superoxide-generating NAD(P)H oxidase activity | 5 (NCF2, DUOX2, NOX4, NOX5, CYBB) of 11 | 2.49 | 3.54 × 10−8 |
| 0016174 | NAD(P)H oxidase H2O2-forming activity | 2 (DUOX2, NOX4) of 9 | 2.18 | 0.0197 |
| 0004601 | Peroxidase activity | 5 (PTGS1, PTGS2, DUOX2, GPX2, CYGB) of 41 | 1.92 | 3.00 × 10−6 |
| 0016209 | Antioxidant activity | 7 (PTGS1, PTGS2, DUOX2, GPX2, TXNRD1, SRXN1, CYGB) of 74 | 1.8 | 3.54 × 10−8 |
| 0050660 | Flavin adenine dinucleotide binding | 5 (NOX4, NOX5, CYBB, TXNRD1, DHCR24) of 82 | 1.61 | 4.94 × 10−5 |
| 0016651 | Oxidoreductase activity, acting on NAD(P)H | 6 (NCF2, DUOX2, NOX4, NOX5, CYBB, TXNRD1) of 107 | 1.58 | 4.93 × 10−6 |
| 0009055 | Electron transfer activity | 4 (NCF2, NOX4, GPX2, TXNRD1) of 103 | 1.42 | 0.0039 |
| 0016491 | Oxidoreductase activity | 13 (PTGS1, PTGS2, ALOX12, NCF2, DUOX2, NOX4, NOX5, CYBB, GPX2, TXNRD1, SRXN1, CYGB, DHCR24) of 726 | 1.08 | 3.54 × 10−8 |
| GO-Term | Description of Functions | Count in Network | Strength | False Discovery Rate |
|---|---|---|---|---|
| 0008379 | Thioredoxin peroxidase activity | 2 (PRDX3, PRDX4) of 5 | 2.75 | 0.0036 |
| 0016175 | Superoxide-generating NAD(P)H oxidase activity | 4 (NCF1, NCF2, NOX5, CYBB) of 11 | 2.71 | 1.80 × 10−7 |
| 0016176 | Superoxide-generating NADPH oxidase activator activity | 2 (NCF1. NCF2) of 9 | 2.49 | 0.0070 |
| 0016668 | Oxidoreductase activity, acting on a sulfur group of donors | 2 (TXNRD1, PRDX3) of 12 | 2.37 | 0.0099 |
| 0016209 | Antioxidant activity | 6 (PRDX4, TXNRD1, SRXN1, PRDX3, SOD3, PTGS2) of 74 | 2.05 | 1.82 × 10−8 |
| 0004601 | Peroxidase activity | 3 (PRDX3, PRDX4, PTGS2) of 41 | 2.01 | 0.0018 |
| 0016651 | Oxidoreductase activity, acting on NAD(P)H | 6 (TXNRD1, PRDX3, NCF1, NCF2, NOX5, CYBB) of 107 | 1.89 | 1.01 × 10−7 |
| 0016667 | Oxidoreductase activity, acting on a sulfur group of donors | 3 (TXNRD1, PRDX3, SRXN1) of 59 | 1.85 | 0.0036 |
| 0050660 | Flavin adenine dinucleotide binding | 3 (TXNRD1, CYBB, NOX5) of 82 | 1.71 | 0.0070 |
| 0009055 | Electron transfer activity | 3 (TXNRD1, NCF1, NCF2) of 103 | 1.61 | 0.0116 |
| 0016491 | Oxidoreductase activity | 10 (TXNRD1, PRDX3, PRDX4, SRXN1, SOD3, NOX5, NCF1, NCF2, CYBB, PTGS2) of 726 | 1.28 | 1.50 × 10−8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, J.; Fu, M.; Gao, J.; Dai, G.; Guan, Q.; Du, C. Upregulation of Thioredoxin Reductase 1 Expression by Flavan-3-Ols Protects Human Kidney Proximal Tubular Cells from Hypoxia-Induced Cell Death. Antioxidants 2022, 11, 1399. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11071399
Zhu J, Fu M, Gao J, Dai G, Guan Q, Du C. Upregulation of Thioredoxin Reductase 1 Expression by Flavan-3-Ols Protects Human Kidney Proximal Tubular Cells from Hypoxia-Induced Cell Death. Antioxidants. 2022; 11(7):1399. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11071399
Chicago/Turabian StyleZhu, Jixiao, Manqin Fu, Jian Gao, Guoyu Dai, Qiunong Guan, and Caigan Du. 2022. "Upregulation of Thioredoxin Reductase 1 Expression by Flavan-3-Ols Protects Human Kidney Proximal Tubular Cells from Hypoxia-Induced Cell Death" Antioxidants 11, no. 7: 1399. https://0-doi-org.brum.beds.ac.uk/10.3390/antiox11071399