The combined analysis of variance across sites included balanced datasets for 169 testing breeding clones and released cultivars revealed highly significant (
p < 0.001) genotype, environment and genotype × environment interaction terms for all the traits evaluated. Tuber starch percentage and total tuber yield had the lowest coefficient of variation across the trial sites (6.04 and 17.94%, respectively), while the coefficient of variation was high for tuber weight by grading sizes (above 25%). The low coefficient of variation for tuber starch percentage indicates the high precision of the experiment for this characteristic. There were highly significant differences among the BLUPs of breeding clones and released cultivars for all the traits (
Supplementary Table S2) across and within the sites. These results clearly show the great heritable variation for traits evaluated, particularly tuber starch percentage and total tuber weight, and potential for further improving these through selection and recombination. Of the 10 best performing potatoes across the sites, according to their BLUPs for total tuber weight per plot, seven were breeding clones and three were released cultivars (
Table 1). The cultivar check Connect was, on average, the top yielding in the field trials, while the tuber yield of the other four cultivar checks was below that of the best performing breeding clones and cultivars. SLU 1402009 and SLU 1314015 were rated among the breeding clones that were most highly resistant to late blight at Mosslunda, where the oomycete
Phytophthora infestans was ubiquitous. Both were also among the top yielding breeding clones and cultivars at the high yield potential site under fungicide treatment in Helgegården, but the former was unselected for the table potato market due to the very high percentage (>2/3) of tubers in the largest size (60 mm). Their reducing sugar content were the same (0.25%), but cultivar checks such as ‘Bintje’ and ‘King Edward (0.175%) were lower. Hence, these breeding clones do not seem to be appropriate for crisps because a reading above 0.1% when using the color chart potato glucose test paper will likely produce a dark color.
Table 2 lists the estimates of variance components and broad-sense heritability (
H2), which confirm the significant genetic variation available in Svensk potatisföradling clones and European released cultivars. Tuber starch percentage and total tuber weight were the traits with the highest
H2, while the weight for the smallest tuber size (<40 mm) has the highest
H2 among the different tuber size categories. The
H2 estimates within each site were larger than those across sites (
Table 3), thus confirming the importance of the significant genotype × environment interaction. The highest estimates were at the high yield potential testing site in Helgegården, except for the total tuber weight and for the weight of tubers 50–60 mm in size, both of which had the largest
H2 in the long daylength testing site at Umeå. This difference could result from true genetic variation between the sites since the coefficients of variation for both tuber weight and for weight of tubers 50–60 mm in size were smaller in Helgegården (12.32 and 22.98%, respectively) than in Umeå (13.78 and 33.46%, respectively). Mosslunda had the smallest
H2 estimates (
Table 3) and the highest coefficient of variation (>25%—except for tuber starch: 6.74) for all the characteristics. In this late blight-prone site, the
H2 estimates for total tuber weight and tuber starch percentages were smaller (0.7522 and 0.9174, respectively), after adjusting their BLUPs using the area under the disease progress curve (AUDPC) as a co-variate, thus showing the effect of the disease on both traits. The
H2 for reducing sugars in the tuber was 0.79 at Umeå, while it was 0.94 for host plant resistance to late blight, as measured by the AUDPC in Mosslunda. The smallest coefficient of variation in each site was always for tuber starch percentage (ranging from 3.60% at Helgegården to 8.44 at Umeå), while it was 8.64% for AUDPC in Mosslunda. As indicated by the
H2 estimates, there was enough genetic variation for selecting for starch content tuber weight (varying on sizes) across the sites, reducing sugars in Umeå and host plant resistance to late blight in Skåne. When using large sample sizes (N > 150), the
H2 estimates remain relatively high in both sites in Skåne, even when late blight occurs.
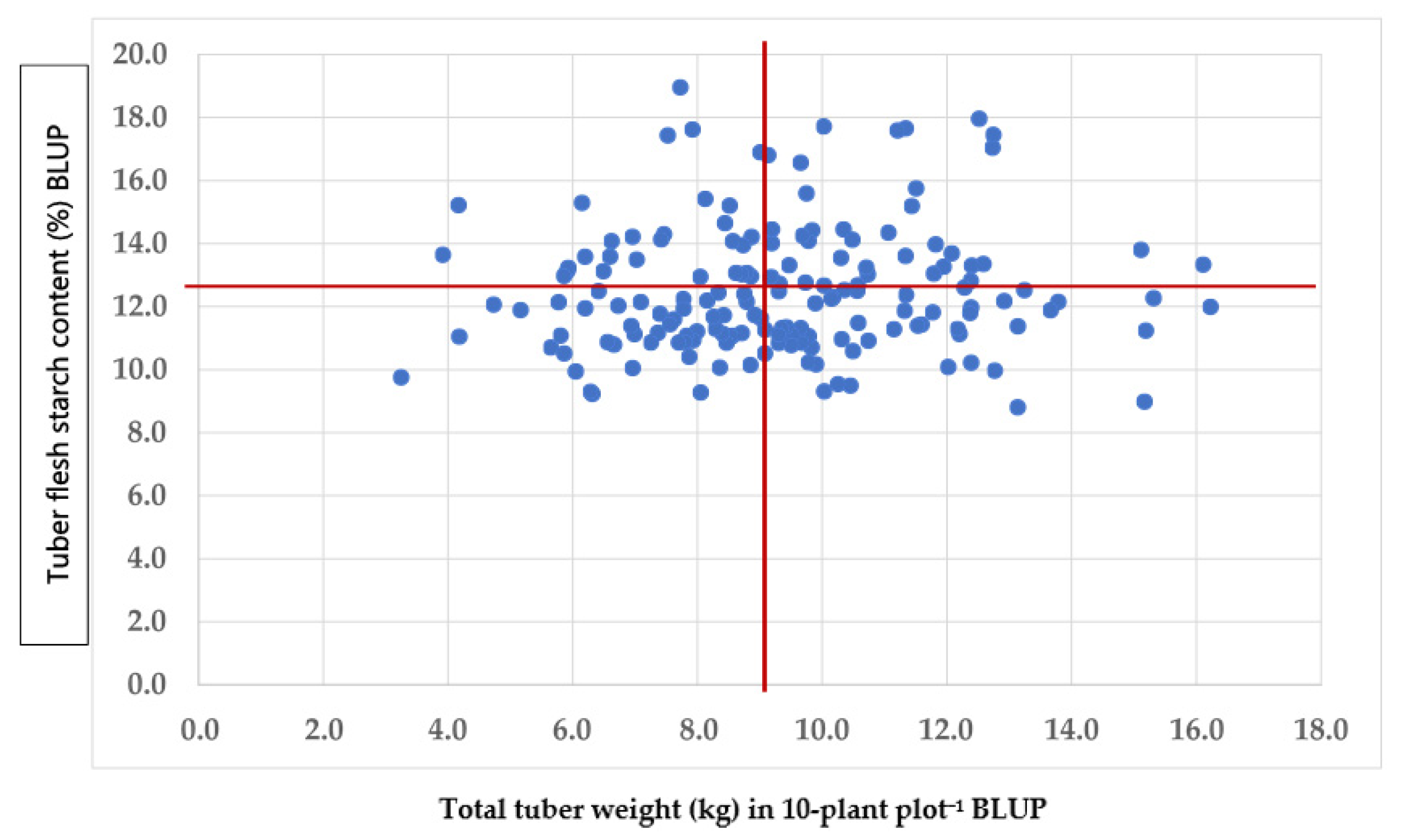
There were significant (
p ≤ 0.05) genetic and phenotypic correlations among the tuber weight characteristics irrespective of their sizes, but none were significant (
p > 0.05) with tuber starch percentages (
Table 4,
Figure 1). Most genetic correlations were larger than their respective phenotypic correlations when they were estimated across sites or within each site. The correlations among the tuber weight characteristics were not always significant in each site (
Table 5,
Table 6 and
Table 7). The genetic and phenotypic correlations between tuber starch percentage and tuber weight (total and for different sizes) in the three testing sites, or between tuber weight with reducing sugars at Umeå were not significant (
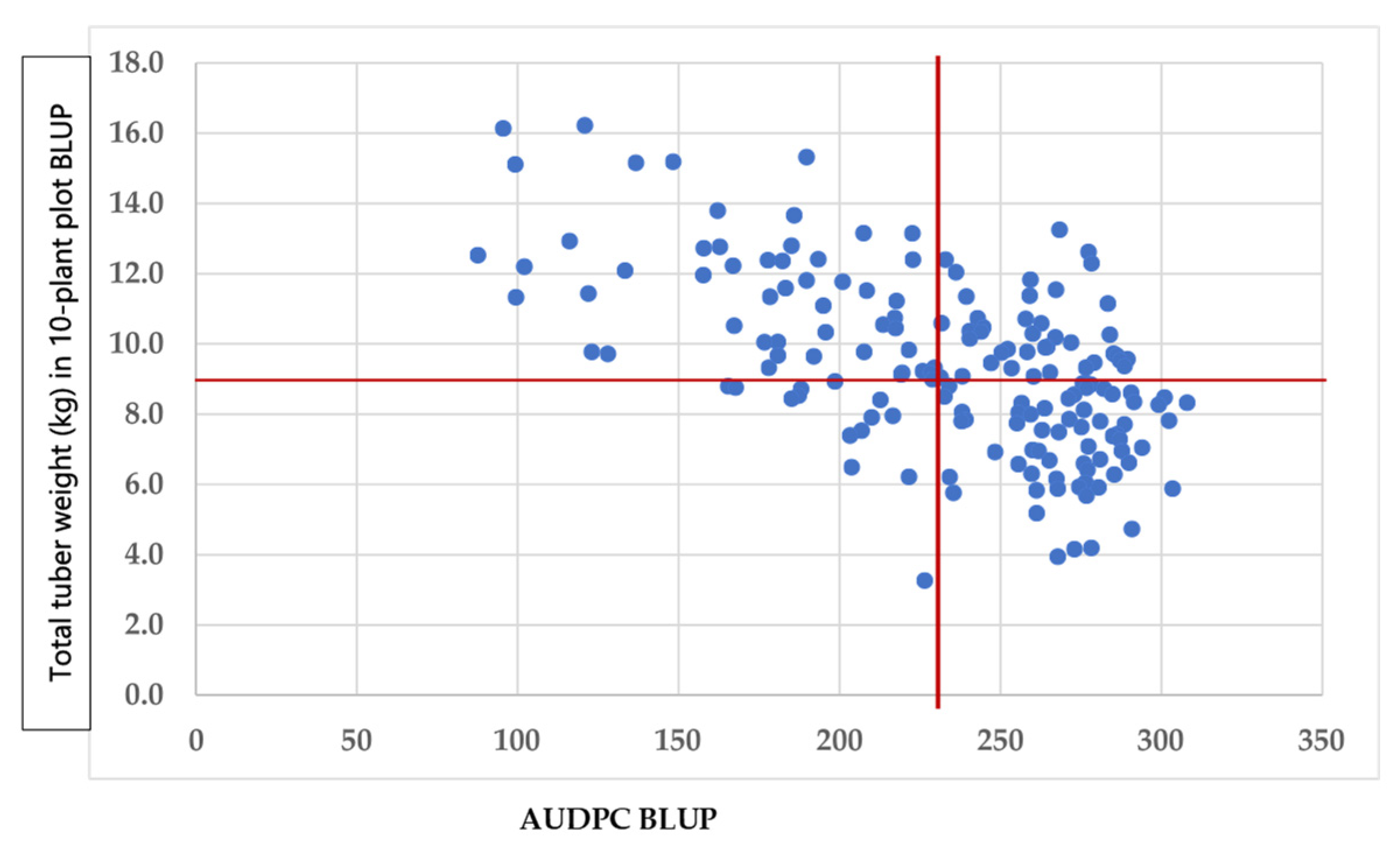
p > 0.05). Hence, the quality traits of the tubers may be improved without affecting the tuber weight. The AUDPC had negative and significant (
p < 0.05) genetic (ranging from −0.25 to −0.78) and phenotypic (varying from −0.19 to −0.71) correlations with the tuber traits, except for the weight of tubers 40 mm in size (0.08 and 0.07, respectively) at Mosslunda. These results indicate that the lower the area under disease progress, i.e., partial resistance, the higher the tuber weight (total and for tubers above 40 mm size) or starch percentage (
Figure 2).







{kind=link}
{kind=link}