
High-Level Carbapenem Resistance among OXA-48-Producing Klebsiellapneumoniae with Functional OmpK36 Alterations: Maintenance of Ceftazidime/Avibactam Susceptibility

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Carbapenems and Ceftazidime/Avibactam Susceptibility, β-Lactamases and Molecular Epidemiology
2.2. Molecular Analysis of OmpK35 and OmpK36 in OXA-48 Producers
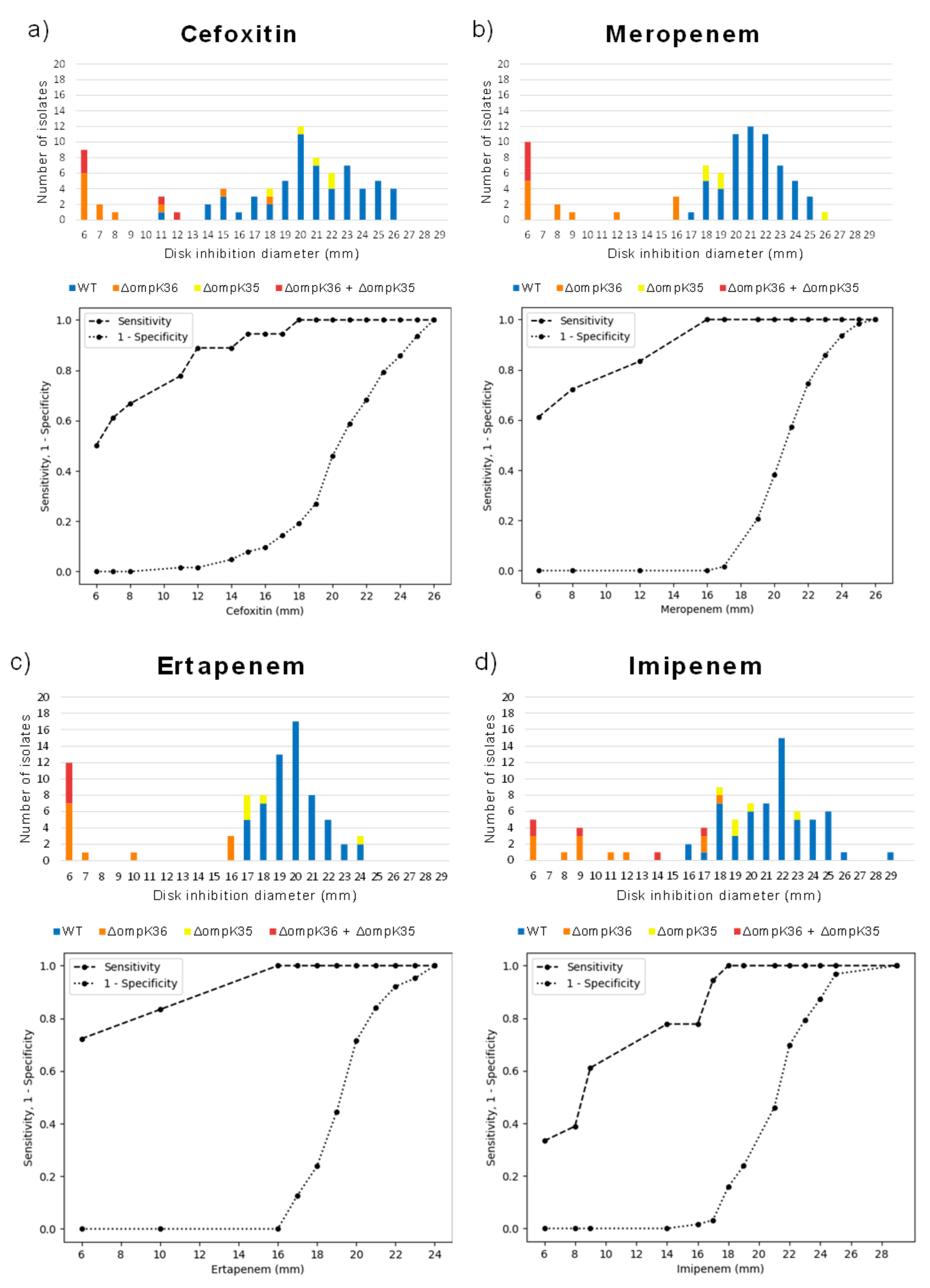
2.3. Disk Diffusion Assay Using Carbapenems and Cefoxitin for Detection of Functional Alteration in OmpK36:
3. Materials and Methods
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vardakas, K.; Tansarli, G.; Rafailidis, P.; Falagas, M. Carbapenems versus alternative antibiotics for the treatment of bacteraemia due to Enterobacteriaceae producing extended-spectrum β-lactamases: A systematic review and meta-analysis. J. Antimicrob. Chemother. 2012, 67, 2793–2803. [Google Scholar] [CrossRef] [PubMed]
- Tamma, P.; Han, J.; Rock, C.; Harris, A.; Lautenbach, E.; Hsu, A.; Avdic, E.; Cosgrove, S. Carbapenem therapy is associated with improved survival compared with piperacillin-tazobactam for patients with extended-spectrum β-lactamase bacteremia. Clin. Infect. Dis. 2015, 60, 1319–13125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haidar, G.; Clancy, C.; Chen, L.; Samanta, P.; Shields, R.; Kreiswirth, B.; Nguyen, M. Identifying spectra of activity and therapeutic niches for ceftazidime-avibactam and imipenem-relebactam against carbapenem-resistant Enterobacteriaceae. Antimicrob. Agents Chemother. 2017, 61, e00642–17. [Google Scholar] [CrossRef] [Green Version]
- Hamzaoui, Z.; Ocampo-Sosa, A.; Fernandez Martinez, M.; Landolsi, S.; Ferjani, S.; Maamar, E.; Saidani, M.; Slim, A.; Martinez-Martinez, L.; Boutiba-Ben Boubaker, I. Role of association of OmpK35 and OmpK36 alteration and blaESBL and/or blaAmpC genes in conferring carbapenem resistance among non-carbapenemase producing Klebsiella pneumoniae. Int. J. Antimicrob. Agents 2018, 52, 898–905. [Google Scholar] [CrossRef]
- Van Duin, D.; Doi, Y. The global epidemiology of carbapenemase-producing Enterobacteriaceae. Virulence 2016, 8, 460–469. [Google Scholar] [CrossRef]
- Papp-Wallace, K.; Endimiani, A.; Taracila, M.; Bonomo, R. Carbapenems: Past, Present, and Future. Antimicrob. Agents Chemother. 2011, 55, 4943–4960. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Baño, J.; Gutiérrez-Gutiérrez, B.; Machuca, I.; Pascual, A. Treatment of infections caused by extended-spectrum-beta-lactamase-, AmpC-, and carbapenemase-producing Enterobacteriaceae. Clin. Microb. Rev. 2018, 31, e00079–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez-Gutiérrez, B.; Salamanca, E.; de Cueto, M.; Hsueh, P.; Viale, P.; Paño-Pardo, J.; Venditti, M.; Tumbarello, M.; Daikos, G.; Cantón, R.; et al. Effect of appropriate combination therapy on mortality of patients with bloodstream infections due to carbapenemase-producing Enterobacteriaceae (INCREMENT): A retrospective cohort study. Lancet Infect. Dis. 2017, 17, 726–734. [Google Scholar] [CrossRef]
- Poirel, L.; Potron, A.; Nordmann, P. OXA-48-like carbapenemases: The phantom menace. J. Antimicrob. Chemother. 2012, 67, 1597–1606. [Google Scholar] [CrossRef] [Green Version]
- Pitout, J.; Peirano, G.; Kock, M.; Strydom, K.; Matsumura, Y. The Global Ascendency of OXA-48-Type Carbapenemases. Clin. Microb. Rev. 2019, 33, 00102–19. [Google Scholar] [CrossRef] [PubMed]
- Zhanel, G.; Lawson, C.; Adam, H.; Schweizer, F.; Zelenitsky, S.; Lagacé-Wiens, P.; Denisuik, A.; Rubinstein, E.; Gin, A.; Hoban, D.; et al. Ceftazidime-Avibactam: A Novel Cephalosporin/β-lactamase Inhibitor Combination. Drugs 2013, 73, 159–177. [Google Scholar] [CrossRef] [Green Version]
- García-Fernaández, A.; Miriagou, V.; Papagiannitsis, C.; Giordano, A.; Venditti, M.; Mancini, C.; Carattoli, A. An ertapenem-resistant extended-spectrum-β-lactamase-producing Klebsiella pneumoniae clone carries a novel OmpK36 porin variant. Antimicrob. Agents Chemother. 2010, 54, 4178–4184. [Google Scholar] [CrossRef] [Green Version]
- Netikul, T.; Kiratisin, P. Genetic characterization of carbapenem-resistant Enterobacteriaceae and the spread of carbapenem-resistant Klebsiella pneumonia ST340 at a university hospital in Thailand. PLoS ONE 2015, 10, e0139116. [Google Scholar] [CrossRef]
- Tsai, Y.; Fung, C.; Lin, J.; Chen, J.; Chang, F.; Chen, T.; Siu, L. Klebsiella pneumoniae outer membrane porins OmpK35 and OmpK36 play roles in both antimicrobial resistance and virulence. Antimicrob. Agents Chemother. 2011, 55, 1485–1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clancy, C.; Chen, L.; Hong, J.; Cheng, S.; Hao, B.; Shields, R.; Farrell, A.; Doi, Y.; Zhao, Y.; Perlin, D.; et al. Mutations of the ompK36 porin gene and promoter impact responses of sequence type 258, KPC-2-producing Klebsiella pneumoniae strains to doripenem and doripenem-colistin. Antimicrob. Agents Chemother. 2013, 57, 5258–5265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Jiang, X.; Wang, Y.; Li, G.; Tian, Y.; Liu, H.; Ai, F.; Ma, Y.; Wang, B.; Ruan, F.; et al. Contribution of β-lactamases and porin proteins OmpK35 and OmpK36 to carbapenem resistance in clinical isolates of KPC-2-producing Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2013, 58, 1214–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, J.; Romano, F.; Kelly, L.; Kwong, H.; Low, W.; Brett, S.; Clements, A.; Beis, K.; Frankel, G. OmpK36-mediated Carbapenem resistance attenuates ST258 Klebsiella pneumoniae in vivo. Nat. Com. 2019, 10, 3957. [Google Scholar] [CrossRef] [Green Version]
- Lunha, K.; Chanawong, A.; Lulitanond, A.; Wilailuckana, C.; Charoensri, N.; Wonglakorn, L.; Saenjamla, P.; Chaimanee, P.; Angkititrakul, S.; Chetchotisakd, P. High-level carbapenem-resistant OXA-48-producing Klebsiella pneumoniae with a novel OmpK36 variant and low-level, carbapenem-resistant, non-porin-deficient, OXA-181-producing Escherichia coli from Thailand. Diag. Microb. Infect. Dis. 2016, 85, 221–226. [Google Scholar] [CrossRef]
- Navon-Venezia, S.; Kondratyeva, K.; Carattoli, A. Klebsiella pneumoniae: A major worldwide source and shuttle for antibiotic resistance. FEMS Microb. Rev. 2017, 41, 252–275. [Google Scholar] [CrossRef]
- Peirano, G.; Chen, L.; Kreiswirth, B.; Pitout, J. Emerging antimicrobial-resistant high-risk Klebsiella pneumoniae Clones ST307 and ST147. Antimicrob. Agents Chemother. 2020, 64, e01148–20. [Google Scholar] [CrossRef]
- Cornaglia, G.; Mazzariol, A.; Fontana, R.; Satta, G. Diffusion of carbapenems through the outer membrane of Enterobacteriaceae and correlation of their activities with their periplasmic concentrations. Microb. Drug. Res. 1996, 2, 273–276. [Google Scholar] [CrossRef]
- Sugawara, E.; Kojima, S.; Nikaido, H. Klebsiella pneumoniae major porins OmpK35 and OmpK36 allow more efficient diffusion of β-lactams than their Escherichia coli homologs OmpF and OmpC. J. Bacter. 2016, 198, 3200–3208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microb. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagès, J.; Peslier, S.; Keating, T.; Lavigne, J.; Nichols, W.; Nichols, W. Role of the outer membrane and porins in susceptibility of β-lactamase-producing Enterobacteriaceae to ceftazidime-avibactam. Antimicrob. Agents Chemother. 2015, 60, 1349–1359. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Martínez, L.; Hernández-Allés, S.; Albertí, S.; Tomás, S.; Benedi, V.; Jacoby, G. In vivo selection of porin-deficient mutants of Klebsiella pneumoniae with increased resistance to cefoxitin and expanded-spectrum-cephalosporins. Antimicrob. Agents Chemother. 1996, 40, 342–348. [Google Scholar] [CrossRef] [Green Version]
- European Committee on Antimicrobial Susceptibility Testing (EUCAST). Breakpoint Tables for Interpretation of Mics and Zone Diameters. Version 11.0. 2021. Available online: https://www.eucast.org/clinical_breakpoints/ (accessed on 2 March 2021).
- Rodríguez-Lucas, C.; Rodicio, M.; Rosete, Y.; Fernández, J. Prospective evaluation of an easy and reliable work flow for the screening of OXA-48-producing Klebsiella pneumoniae in endemic settings. J. Hosp. Infect. 2020, 105, 659–662. [Google Scholar] [CrossRef]
- Zerbino, D.; Birney, E. Velvet: Algorithms for de novo short read assembly using de Bruijn graphs. Genome Res. 2008, 18, 821–829. [Google Scholar] [CrossRef] [Green Version]
- CGE. Center for Genomic Epidemiology. 2020. Available online: http://www.genomicepidemiology.org (accessed on 25 January 2021).
- Madeira, F.; Park, Y.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.; Potter, S.; Finn, R.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucl. Acids Res 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, A.; Procter, J.; Martin, D.; Clamp, M.; Barton, G. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [Green Version]
- Hunter, J. Matplotlib: A 2D graphics environment. Comput. Sci. Eng. 2017, 9, 90–95. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Strain | ST | Sample | Previous Carbapenem Exposure (days) 1 | ESBL Enzyme | FOX MIC/IZD 2 | MER MIC/IZD 2 | ERT MIC/IZD2 | IPM vMIC/IZD 2 | CAZ/AVI MIC | ompK35 | ompK36 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Kp_HUCA_Bac_5 | 326 | Blood culture | Yes (10) | CTX-M-15 | >16/6 | >32/6 | >4/6 | >8/6 | 0.5 | WT | ::IS5 |
| Kp_HUCA_Bac_7 | 326 | Blood culture | No | CTX-M-15 | >16/6 | 16/6 | >4/6 | >8/6 | 0.75 | WT | ::IS5 |
| Kp_HUCA_Bac_10 | 326 | Blood culture | Yes (21) | CTX-M-15 | >16/6 | >32/8 | >4/6 | >8/9 | 0.5 | WT | ΔC201 |
| Kp_HUCA_Bac_13 | 326 | Blood culture | Yes (11) | CTX-M-15 | >16/6 | >32/9 | >4/7 | >8/11 | 0.75 | WT | G259T |
| Kp_HUCA_Bac_18 | 326 | Blood culture | No | CTX-M-15 | >16/7 | 32/8 | >4/6 | >8/9 | 0.5 | WT | ΔC201 |
| Kp_HUCA_Bac_24 | 326 | Blood culture | Yes (5) | CTX-M-15 | >16/11 | >32/6 | >1/6 | >8/9 | 0.5 | ::623T | ::IS5 |
| Kp_HUCA_Bac_64 | 16 | Blood culture | Yes (5) | CTX-M-15 | 16/11 | 8/16 | >4/16 | 8/18 | 0.5 | WT | ΔA74 |
| Kp_HUCA_Bac_65 | 147 | Blood culture | No | SHV-12 | ≤8/15 | 8/16 | >1/16 | 4/19 | 0.38 | WT | ::IS5 |
| Kp_HUCA_Bac_76 | 405 | Blood culture | Yes (7) | CTX-M-15 | >16/12 | >32/6 | >1/6 | >8/6 | 0.75 | ::623T | T612A |
| Kp_HUCA_Bac_85 | 273 | Blood culture | Yes (19) | Non-ESBL | >16/6 | >32/6 | >1/6 | >8/6 | 0.5 | ::623T | ::29TG |
| Kp_HUCA_Bac_88 | 405 | Blood culture | Yes (6) | Non-ESBL | 16/18 | 8/16 | >1/16 | 8/17 | 0.19 | WT | ::818ACAAAGCGCAGAACTTCGAACCTGGGCTTTGCAA |
| Kp_HUCA_Bac_90 | 101 | Blood culture | Yes (2) | CTX-M-15 | >16/6 | 24/6 | >1/6 | 4/17 | 0.5 | ΔG185 | ::403GACGGC |
| Kp_HUCA_2 | 326 | Rectal swab | No | CTX-M-15 | >16/6 | >32/6 | >1/6 | >8/8 | 0.5 | WT | G844T |
| Kp_HUCA_3 | 15 | Sputum | No | CTX-M-15 | >16/6 | >32/6 | >1/6 | >8/14 | 0.75 | ::623T | ::403GACGGC |
| Kp_HUCA_4 | 15 | Urine | Yes (15) | CTX-M-15 | >16/7 | >32/6 | >1/6 | >8/6 | 0.75 | WT | Δ(886)-184 |
| Kp_HUCA_5 | 193 | Urine | No | Non-ESBL | >16/8 | 16/12 | >1/10 | >8/12 | 0.5 | WT | ::IS1 |
| Kp_HUCA_8 | 326 | Rectal swab | Yes (4) | CTX-M-15 | >16/6 | 32/6 | >1/6 | >8/9 | 0.5 | WT | ΔA4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lumbreras-Iglesias, P.; Rodicio, M.R.; Valledor, P.; Suárez-Zarracina, T.; Fernández, J. High-Level Carbapenem Resistance among OXA-48-Producing Klebsiellapneumoniae with Functional OmpK36 Alterations: Maintenance of Ceftazidime/Avibactam Susceptibility. Antibiotics 2021, 10, 1174. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10101174
Lumbreras-Iglesias P, Rodicio MR, Valledor P, Suárez-Zarracina T, Fernández J. High-Level Carbapenem Resistance among OXA-48-Producing Klebsiellapneumoniae with Functional OmpK36 Alterations: Maintenance of Ceftazidime/Avibactam Susceptibility. Antibiotics. 2021; 10(10):1174. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10101174
Chicago/Turabian StyleLumbreras-Iglesias, Pilar, María Rosario Rodicio, Pablo Valledor, Tomás Suárez-Zarracina, and Javier Fernández. 2021. "High-Level Carbapenem Resistance among OXA-48-Producing Klebsiellapneumoniae with Functional OmpK36 Alterations: Maintenance of Ceftazidime/Avibactam Susceptibility" Antibiotics 10, no. 10: 1174. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10101174