
Antimicrobial Activity of a Lipidated Temporin L Analogue against Carbapenemase-Producing Klebsiella pneumoniae Clinical Isolates

 , and
, and
Abstract
:1. Introduction
2. Results
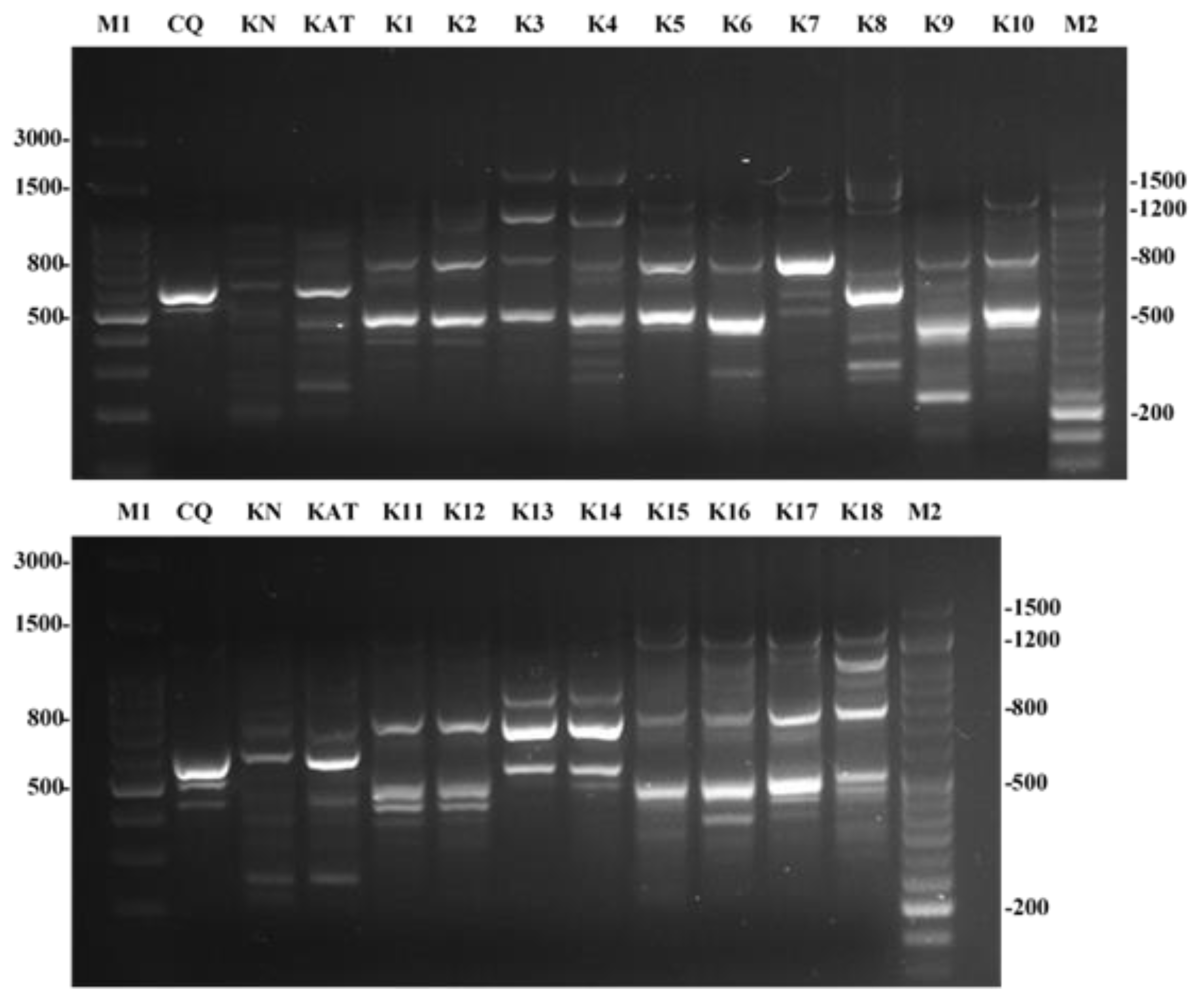
2.1. Antibiotic Susceptibility and RAPD Profiles
2.2. Antimicrobial Activity of Peptides
3. Discussion
4. Materials and Methods
4.1. Synthesis
4.2. Bacterial Strains and Culture Conditions
4.3. Antimicrobial Assays: Determination of the Minimum Inhibitory Concentration (MIC) and the Minimum Bactericidal Concentration (MBC)
4.4. RAPD Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Vading, M.; Nauclér, P.; Kalin, M.; Giske, C.G. Invasive infection caused by Klebsiella pneumoniae is a disease affecting patients with high comorbidity and associated with high long-term mortality. PLoS ONE 2018, 13, e0195258. [Google Scholar] [CrossRef]
- Vardakas, K.Z.; Tansarli, G.S.; Rafailidis, P.I.; Falagas, M.E. Carbapenems versus alternative antibiotics for the treatment of bacteraemia due to Enterobacteriaceae producing extended-spectrum β-lactamases: A systematic review and meta-analysis. J. Antimicrob. Chemother. 2012, 67, 2793–2803. [Google Scholar] [CrossRef]
- Walsh, T.R. Emerging carbapenemases: A global perspective. Int. J. Antimicrob. Agents 2010, 36, 8–14. [Google Scholar] [CrossRef]
- Pitout, J.D.D.; Nordmann, P.; Poirel, L. Carbapenemase-Producing Klebsiella pneumoniae, a Key Pathogen Set for Global Nosocomial Dominance. Antimicrob. Agents Chemother. 2015, 59, 5873–5884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, X.; Zhang, H.; Du, H. Carbapenemases in Enterobacteriaceae: Detection and Antimicrobial Therapy. Front. Microbiol. 2019, 10, 1823. [Google Scholar] [CrossRef]
- Logan, L.K.; Weinstein, R.A. The Epidemiology of Carbapenem-Resistant Enterobacteriaceae: The Impact and Evolution of a Global Menace. J. Infect. Dis. 2017, 215 (Suppl. 1), S28–S36. [Google Scholar] [CrossRef] [Green Version]
- Ambler, R.P. The structure of beta-lactamases. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1980, 289, 321–331. [Google Scholar]
- Bonomo, R.A.; Burd, E.M.; Conly, J.; Limbago, B.M.; Poirel, L.; Segre, J.L.; Westblade, L.F. Carbapenemase-Producing Organisms: A Global Scourge. Clin. Infect. Dis. 2018, 66, 1290–1297. [Google Scholar] [CrossRef]
- Trecarichi, E.M.; Tumbarello, M. Therapeutic options for carbapenem-resistant Enterobacteriaceae infections. Virulence 2017, 8, 470–484. [Google Scholar] [CrossRef]
- WHO. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics. World Health Organization, 2017. Available online: https://www.who.int/medicines/publications/WHO-PPL-Short_Summary_25Feb-ET_NM_WHO.pdf (accessed on 3 August 2021).
- Bassetti, M.; Vena, A.; Sepulcri, C.; Giacobbe, D.R.; Peghin, M. Treatment of Bloodstream Infections Due to Gram-Negative Bacteria with Difficult-to-Treat Resistance. Antibiotics 2020, 9, 632. [Google Scholar] [CrossRef] [PubMed]
- Mangoni, M.L.; Grazia, A.D.; Cappiello, F.; Casciaro, B.; Luca, V. Naturally occurring peptides from Rana temporaria: Antimicrobial properties and more. Curr. Top. Med. Chem. 2016, 16, 54–64. [Google Scholar] [CrossRef]
- Rinaldi, A.C.; Mangoni, M.L.; Rufo, A.; Luzi, C.; Barra, D.; Zhao, H.; Kinnunen, P.K.; Bozzi, A.; Di Giulio, A.; Simmaco, M. Temporin L: Antimicrobial, haemolytic and cytotoxic activities, and effects on membrane permeabilization in lipid vesicles. Biochem. J. 2002, 368, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Merlino, F.; Carotenuto, A.; Casciaro, B.; Martora, F.; Loffredo, M.R.; Di Grazia, A.; Yousif, A.M.; Brancaccio, D.; Palomba, L.; Novellino, E.; et al. Glycine-replaced derivatives of [Pro(3),DLeu(9)]TL, a temporin L analogue: Evaluation of antimicrobial, cytotoxic and hemolytic activities. Eur. J. Med. Chem. 2017, 139, 750–761. [Google Scholar] [CrossRef] [PubMed]
- Bellavita, R.; Casciaro, B.; Di Maro, S.; Brancaccio, D.; Carotenuto, A.; Falanga, A.; Cappiello, F.; Buommino, E.; Galdiero, S.; Novellino, E.; et al. First-in-class cyclic Temporin L analogue: Design, synthesis, and antimicrobial assessment. J. Med. Chem. 2021, 64, 11675–11694. [Google Scholar] [CrossRef] [PubMed]
- Buommino, E.; Carotenuto, A.; Antignano, I.; Bellavita, R.; Casciaro, B.; Loffredo, M.R.; Merlino, F.; Novellino, E.; Mangoni, M.L.; Nocera, F.P.; et al. The outcomes of decorated prolines in the discovery of antimicrobial peptides from Temporin-L. Chem. Med. Chem. 2019, 14, 1283–1290. [Google Scholar] [CrossRef] [PubMed]
- Bellavita, R.; Vollaro, A.; Catania, M.R.; Merlino, F.; De Martino, L.; Nocera, F.P.; DellaGreca, M.; Lembo, F.; Grieco, P.; Buommino, E. Novel antimicrobial peptide from Temporin L in the treatment of Staphylococcus pseudintermedius and Malassezia pachydermatis in polymicrobial inter-kingdom infection. Antibiotics 2020, 9, 530. [Google Scholar] [CrossRef]
- Bellavita, R.; Raucci, F.; Merlino, F.; Piccolo, M.; Ferraro, M.G.; Irace, C.; Santamaria, R.; Iqbal, A.J.; Novellino, E.; Grieco, P.; et al. Temporin L-derived peptide as a regulator of the acute inflammatory response in zymosan-induced peritonitis. Biomed. Pharmacother. 2020, 123, 109788. [Google Scholar] [CrossRef]
- Bellavita, R.; Falanga, A.; Buommino, E.; Merlino, F.; Casciaro, B.; Cappiello, F.; Mangoni, M.L.; Novellino, E.; Catania, M.R.; Paolillo, R.; et al. Novel temporin L antimicrobial peptides: Promoting self-assembling by lipidic tags to tackle superbugs. J. Enzym Inhib. Med. Chem. 2020, 35, 1751–1764. [Google Scholar] [CrossRef]
- Plazak, M.E.; Tamma, P.D.; Heil, E.L. The antibiotic arms race: Current and emerging therapy for Klebsiella pneumoniae carbapenemase (KPC)—Producing bacteria. Expert. Opin. Pharmacother. 2018, 19, 2019–2031. [Google Scholar] [CrossRef]
- Lee, C.R.; Lee, J.H.; Park, K.S.; Kim, Y.B.; Jeong, B.C.; Lee, S.H. Global Dissemination of Carbapenemase-Producing Klebsiella pneumoniae: Epidemiology, Genetic Context, Treatment Options, and Detection Methods. Front. Microbiol. 2016, 7, 895. [Google Scholar] [CrossRef] [Green Version]
- Navon-Venezia, S.; Kondratyeva, K.; Carattoli, A. Klebsiella pneumoniae: A major worldwide source and shuttle for antibiotic resistance. FEMS Microbiol. Rev. 2017, 41, 252–275. [Google Scholar] [CrossRef]
- Yusuf, E.; Bax, H.I.; Verkaik, N.J.; van Westreenen, M. An Update on Eight “New” Antibiotics against Multidrug-Resistant Gram-Negative Bacteria. J. Clin. Med. 2021, 10, 1068. [Google Scholar] [CrossRef]
- Göttig, S.; Frank, D.; Mungo, E.; Nolte, A.; Hogardt, M.; Besier, S.; Wichelhaus, T.A. Emergence of ceftazidime/avibactam resistance in KPC-3-producing Klebsiella pneumoniae in vivo. J. Antimicrob. Chemother. 2019, 74, 3211–3216. [Google Scholar] [CrossRef]
- Gaibani, P.; Re, M.C.; Campoli, C.; Viale, P.L.; Ambretti, S. Bloodstream infection caused by KPC-producing Klebsiella pneumoniae resistant to ceftazidime/avibactam: Epidemiology and genomic characterization. Clin. Microbiol. Infect. 2020, 26, 516.e1–516.e4. [Google Scholar] [CrossRef]
- Wang, J.; Dou, X.; Song, J.; Lyu, Y.; Zhu, X.; Xu, L.; Li, W.; Shan, A. Antimicrobial peptides: Promising alternatives in the post feeding antibiotic era. Med. Res. Rev. 2019, 39, 831–859. [Google Scholar] [CrossRef] [PubMed]
- Falanga, A.; Galdiero, M.; Galdiero, S. Membranotropic cell penetrating peptides: The outstanding journey. Int. J. Mol. Sci. 2015, 16, 25323–25337. [Google Scholar] [CrossRef]
- Kumar, P.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial peptides: Diversity, mechanism of action and strategies to improve the activity and biocompatibility in vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Falanga, A.; Lombardi, L.; Franci, G.; Vitiello, M.; Iovene, M.R.; Morelli, G.; Galdiero, M.; Galdiero, S. Marine antimicrobial peptides: Nature provides templates for the design of novel compounds against pathogenic bacteria. Int. J. Mol. Sci. 2016, 17, 785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollmann, A.; Martinez, M.; Maturana, P.; Semorile, L.C.; Maffia, P.C. Antimicrobial peptides: Interaction with model and biological membranes and synergism with chemical antibiotics. Front. Chem. 2018, 6, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bechinger, B.; Gorr, S.U. Antimicrobial Peptides: Mechanisms of Action and Resistance. J. Dent. Res. 2017, 96, 254–260. [Google Scholar] [CrossRef] [Green Version]
- Zharkova, M.S.; Orlov, D.S.; Golubeva, O.Y.; Chakchir, O.B.; Eliseev, I.E.; Grinchuk, T.M.; Shamova, O.V. Application of antimicrobial peptides of the innate immune system in combination with conventional antibiotics-a novel way to combat antibiotic resistance? Front. Cell. Infect. Microbiol. 2019, 9, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasfi, R.; Elkhatib, W.F.; Ashour, H.M. Molecular typing and virulence analysis of multidrug resistant Klebsiella pneumoniae clinical isolates recovered from Egyptian hospitals. Sci. Rep. 2016, 6, 38929. [Google Scholar] [CrossRef]
- Boisson, M.; Gregoire, N.; Couet, W.; Mimoz, O. Colistin in critically ill patients. Minerva Anestesiol. 2013, 79, 200–208. [Google Scholar] [PubMed]
- Merlino, F.; Tomassi, S.; Yousif, A.M.; Messere, A.; Marinelli, L.; Grieco, P.; Novellino, E.; Cosconati, S.; Di Maro, S. Boosting Fmoc solid-phase peptide synthesis by ultrasonication. Org. Lett. 2019, 21, 6378–6382. [Google Scholar] [CrossRef] [PubMed]
- Roscetto, E.; Masi, M.; Esposito, M.; Di Lecce, R.; Delicato, A.; Maddau, L.; Calabrò, V.; Evidente, A.; Catania, M.R. Anti-Biofilm Activity of the Fungal Phytotoxin Sphaeropsidin A Against Clinical Isolates of Antibiotic-Resistant Bacteria. Toxins 2020, 12, 444. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Strain Name | Source | Strain Name | Source |
|---|---|---|---|
| KNKp | AF | KPCKp12 | TR |
| KPCKp1 | TR | KPCKp13 | E |
| KPCKp2 | TFr | KPCKp14 | B |
| KPCKp3 | UC | KPCKp15 | TF |
| KPCKp4 | UC | KPCKp16 | TR |
| KPCKp5 | AF | KPCKp17 | U |
| KPCKp6 | U | KPCKp18 | U |
| KPCKp7 | E | ||
| KPCKp8 | TF | VIMKp | AF |
| KPCKp9 | C | NDMKp1 | Tfr |
| KPCKp10 | E | NDMKp2 | U |
| KPCKp11 | P | NDMKp3 | E |
| Strains | AMC | CTX | CAZ | TZP | GM | AK | SXT | CIP | MEM | ERT | CAZ/AVI |
|---|---|---|---|---|---|---|---|---|---|---|---|
| KNKp | 4 (S) | ≤1 (S) | ≤0.5 (S) | ≤4 (S) | ≤1 (S) | ≤4 (S) | ≤1 (S) | ≤0.25 (S) | ≤0.125 (S) | ≤0.25 (S) | 1 (S) |
| KPCKp1 | >16 (R) | >32 (R) | >32 (R) | >64 (R) | ≤1 (S) | ≤1 (S) | 4 (I) | >2 (R) | >8 (R) | >4 (R) | 8 (S) |
| KPCKp2 | >16 (R) | >32 (R) | >32 (R) | >64 (R) | ≤1 (S) | ≤1 (S) | 4 (I) | >2 (R) | >8 (R) | >4 (R) | 1 (S) |
| KPCKp3 | >32 (R) | >4 (R) | >8 (R) | >16 (R) | ≤1 (S) | 4 (S) | >4 (R) | >1 (R) | >8 (R) | >1 (R) | 4 (S) |
| KPCKp4 | >32 (R) | >4 (R) | >8 (R) | >16 (R) | >4 (R) | 4 (S) | >4 (R) | >1 (R) | >8 (R) | >1 (R) | 1 (S) |
| KPCKp5 | >16 (R) | >32 (R) | >32 (R) | >64 (R) | >8 (R) | 4 (S) | >8 (R) | 0.25 (S) | >8 (R) | >4 (R) | 4 (S) |
| KPCKp6 | >32 (R) | >4 (R) | >32 (R) | >64 (R) | 2 (S) | 8 (S) | >4 (R) | >1 (R) | 8 (I) | >1 (R) | 1 (S) |
| KPCKp7 | >32 (R) | >4 (R) | >8 (R) | >16 (R) | 4 (I) | ≤4 (S) | ≤1 (S) | >1 (R) | >8 (R) | >1 (R) | 1 (S) |
| KPCKp8 | >16 (R) | >32 (R) | >32 (R) | >64 (R) | >8 (R) | 32 (R) | >8 (R) | >2 (R) | >8 (R) | >4 (R) | >8 (R) |
| KPCKp9 | >16 (R) | >32 (R) | >32 (R) | >64 (R) | ≤1 (S) | 4 (S) | >8 (R) | >2 (R) | >8 (R) | >4 (R) | 4 (S) |
| KPCKp10 | >16 (R) | >32 (R) | >32 (R) | >64 (R) | ≤1 (S) | ≤1 (S) | >8 (R) | >2 (R) | 8 (I) | >4 (R) | 4 (S) |
| KPCKp11 | >32 (R) | >4 (R) | >8 (R) | >16 (R) | >4 (R) | >16 (R) | >8 (R) | >1 (R) | >8 (R) | >1 (R) | 1 (S) |
| KPCKp12 | >16 (R) | >32 (R) | >32 (R) | >64 (R) | 2 (S) | >32 (R) | ≤1 (S) | >2 (R) | >8 (R) | >4 (R) | 4 (S) |
| KPCKp13 | >16 (R) | >32 (R) | >32 (R) | >64 (R) | >8 (R) | >16 (R) | >8 (R) | >2 (R) | >8 (R) | >4 (R) | 4 (S) |
| KPCKp14 | >16 (R) | >32 (R) | >16 (R) | >64 (R) | >8 (R) | 32 (R) | >8 (R) | >2 (R) | >8 (R) | >4 (R) | 1 (S) |
| KPCKp15 | >16 (R) | >32 (R) | >32 (R) | >64 (R) | >8 (R) | 32 (R) | >8 (R) | >2 (R) | >8 (R) | >1 (R) | >8 (R) |
| KPCKp16 | >16 (R) | >32 (R) | >32 (R) | >64 (R) | 2 (S) | 4 (S) | >8 (R) | >2 (R) | >8 (R) | >4 (R) | 4 (S) |
| KPCKp17 | >16 (R) | >32 (R) | >32 (R) | >64 (R) | ≤1 (S) | ≤1 (S) | ≤1 (S) | >2 (R) | >8 (R) | >4 (R) | 4 (S) |
| KPCKp18 | >32 (R) | >4 (R) | >8 (R) | >16 (R) | 2 (S) | 4 (S) | ≤1 (S) | >1 (R) | >8 (R) | >1 (R) | 4 (S) |
| VIMKp | >32 (R) | >4 (R) | >8 (R) | >16 (R) | 2 (S) | ≤4 (I) | >4 (R) | 1 (R) | 8 (R) | >1 (R) | >8 (R) |
| NDMKp1 | >16 (R) | >32 (R) | >32 (R) | >64 (R) | >8 (R) | 16 (I) | ≤1 (S) | >2 (R) | 8 (R) | >1 (R) | >8 (R) |
| NDMKp2 | >16 (R) | 32 (R) | >32 (R) | >64 (R) | >8 (R) | >16 (R) | >8 (R) | ≤0.06 (S) | >8 (R) | >4 (R) | >8 (R) |
| NDMKp3 | >16 (R) | >32 (R) | >32 (R) | >64 (R) | >8 (R) | 32 (R) | ≤1 (S) | >2 (R) | >8 (R) | >1 (R) | >8 (R) |
| Strains | MIC (µM) | MIC (µg/mL) | |
|---|---|---|---|
| 1B | C | Polymyxin E | |
| KCQ | 6.25 | 6.25 | <2 (S) |
| KAT | 12.5 | 6.25 | <2 (S) |
| KNKp | 6.25 | 6.25 | <2 (S) |
| KPCKp1 | 50 | 6.25 | 8 (R) |
| KPCKp3 | 100 | 25 | <2 (S) |
| KPCKp4 | 25 | 25 | <2 (S) |
| KPCKp5 | 50 | 12.5 | <2 (S) |
| KPCKp6 | 12.5 | 12.5 | <2 (S) |
| KPCKp7 | 12.5 | 12.5 | <2 (S) |
| KPCKp8 | 50 | 12.5 | <2 (S) |
| KPCKp9 | 12.5 | 12.5 | <2 (S) |
| KPCKp10 | 50 | 12.5 | <2 (S) |
| KPCKp11 | 25 | 12.5 | <2 (S) |
| KPCKp12 | 25 | 25 | <2 (S) |
| KPCKp13 | 25 | 12.5 | <2 (S) |
| KPCKp15 | 50 | 25 | 8 (R) |
| KPCKp16 | 25 | 25 | <2 (S) |
| KPCKp17 | 25 | 12.5 | <2 (S) |
| KPCKp18 | 25 | 12.5 | <2 (S) |
| VIMKp | 50 | 25 | <2 (S) |
| NDMKp1 | 25 | 25 | <2 (S) |
| NDMKp2 | 50 | 25 | <2 (S) |
| NDMKp3 | 100 | 25 | <2 (S) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roscetto, E.; Bellavita, R.; Paolillo, R.; Merlino, F.; Molfetta, N.; Grieco, P.; Buommino, E.; Catania, M.R. Antimicrobial Activity of a Lipidated Temporin L Analogue against Carbapenemase-Producing Klebsiella pneumoniae Clinical Isolates. Antibiotics 2021, 10, 1312. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10111312
Roscetto E, Bellavita R, Paolillo R, Merlino F, Molfetta N, Grieco P, Buommino E, Catania MR. Antimicrobial Activity of a Lipidated Temporin L Analogue against Carbapenemase-Producing Klebsiella pneumoniae Clinical Isolates. Antibiotics. 2021; 10(11):1312. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10111312
Chicago/Turabian StyleRoscetto, Emanuela, Rosa Bellavita, Rossella Paolillo, Francesco Merlino, Nicola Molfetta, Paolo Grieco, Elisabetta Buommino, and Maria Rosaria Catania. 2021. "Antimicrobial Activity of a Lipidated Temporin L Analogue against Carbapenemase-Producing Klebsiella pneumoniae Clinical Isolates" Antibiotics 10, no. 11: 1312. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10111312