
Prevalence of the Genes Associated with Biofilm and Toxins Synthesis amongst the Pseudomonas aeruginosa Clinical Strains

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. The MBL Genes
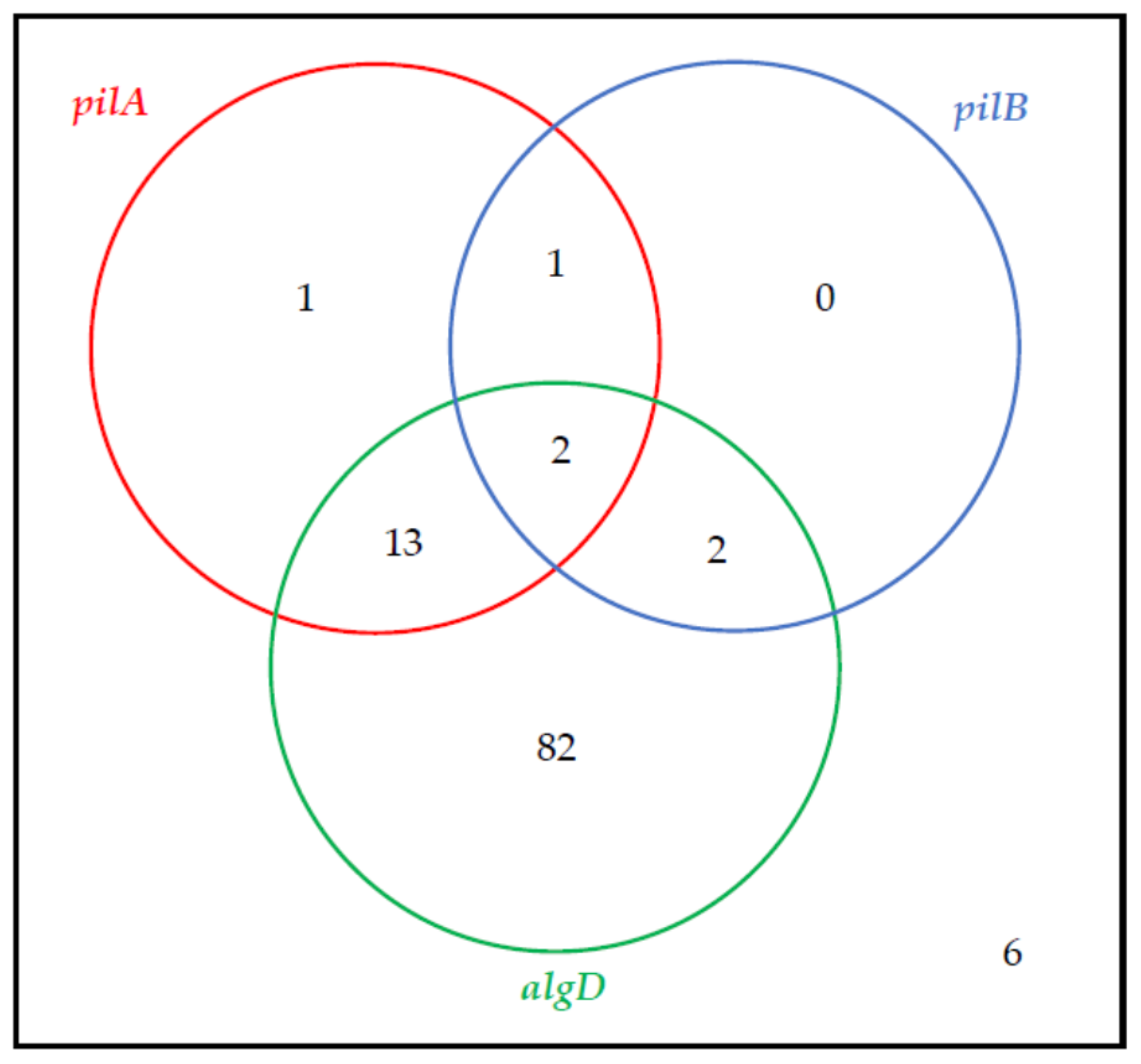
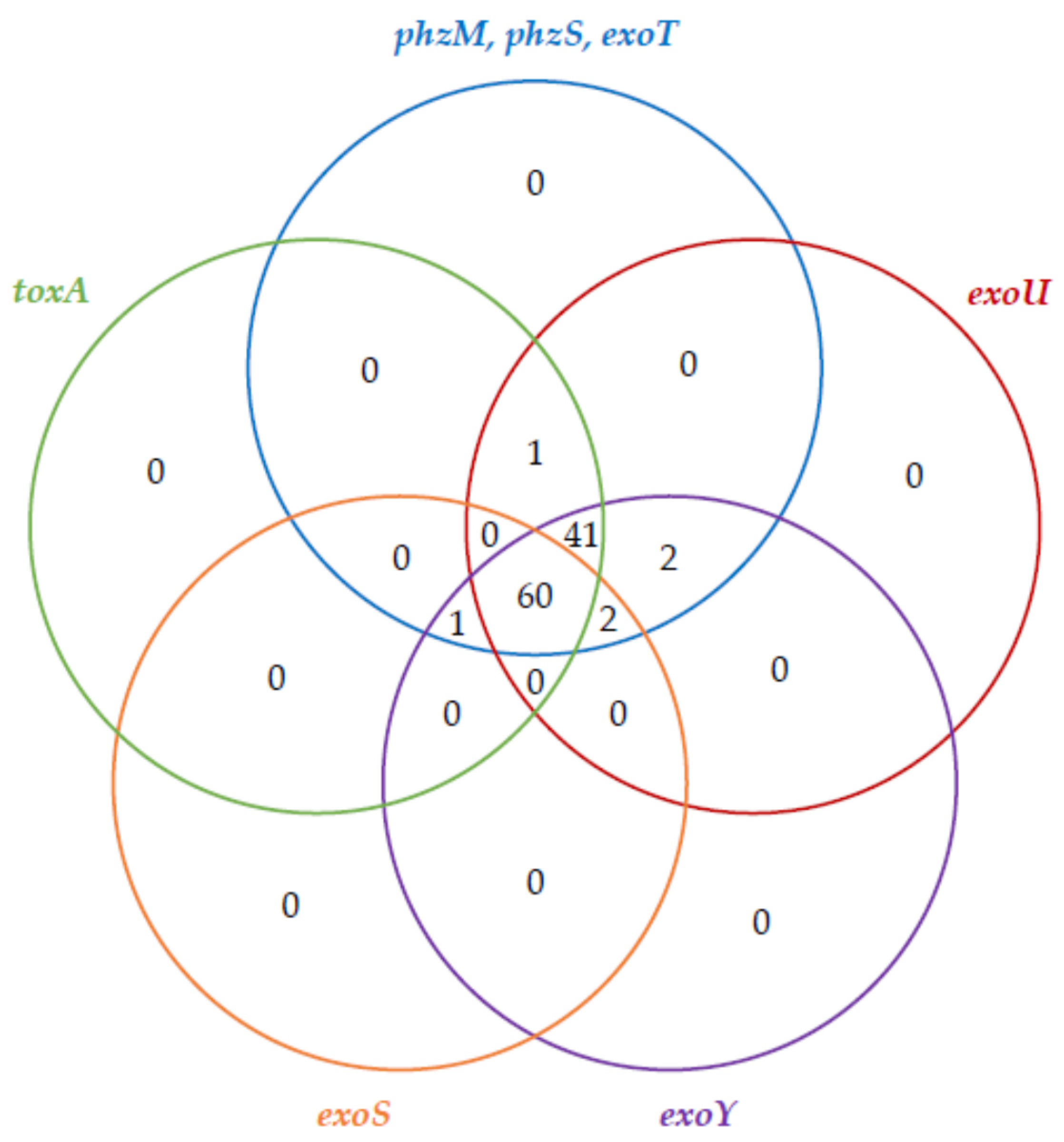
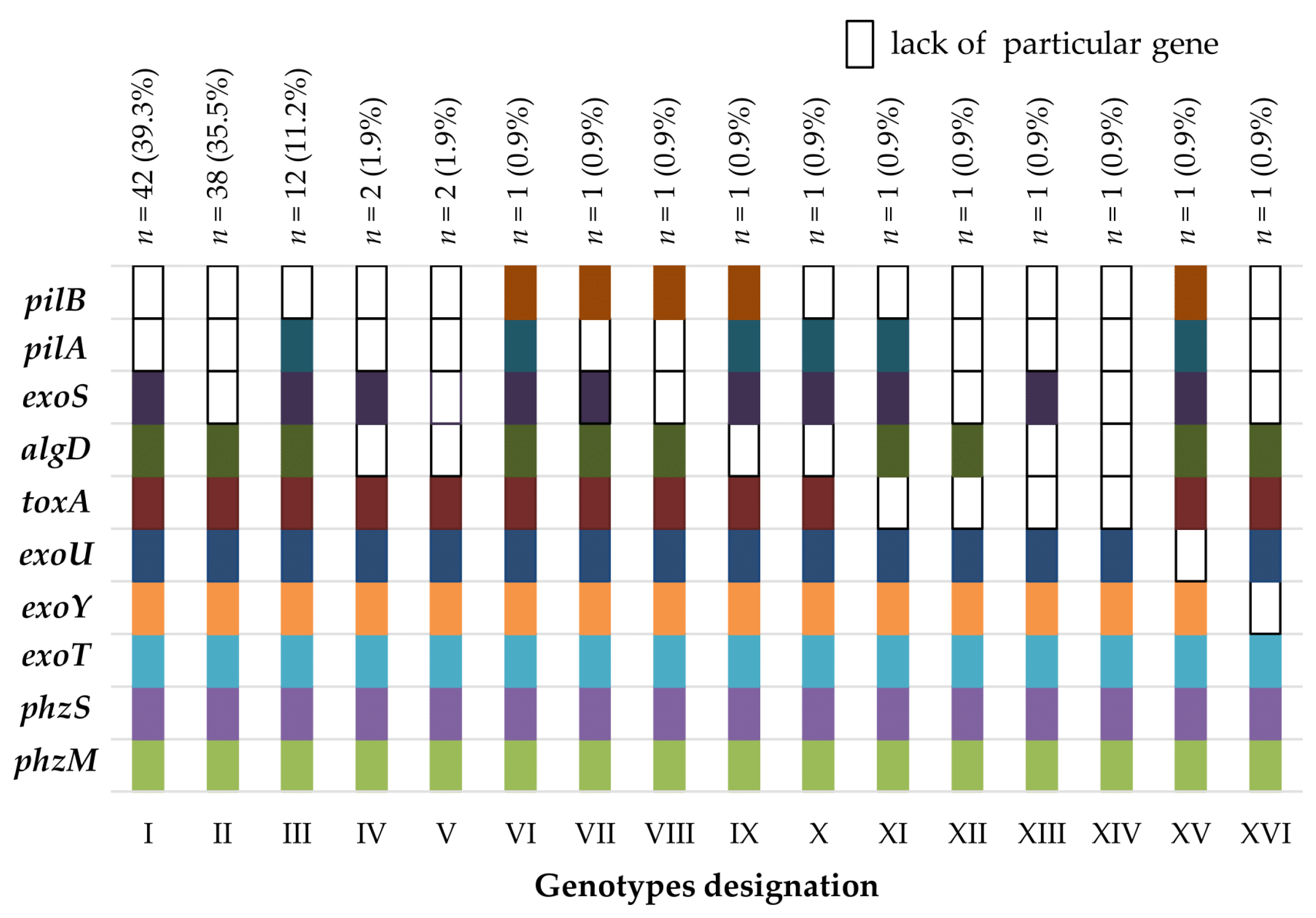
2.2. The Virulence Factor Genes
3. Discussion
4. Materials and Methods
4.1. Origin of Strains and Selection Criteria
4.2. The Susceptibility to Carbapenems and Carbapenemase Activity Evaluation
4.3. Bacterial DNA Isolation
4.4. MBL Genes Detection
4.5. Virulence Factor Genes Detection
4.6. Statistical Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kucisec-Tepes, N. Pseudomonas aeruginosa--a significant hospital pathogen and resistance to carbapenem. Acta Med. Croat. Cas. Hravatske Akad. Med. Znan. 2004, 58, 313–321. [Google Scholar]
- Bogiel, T.; Mikucka, A.; Skalski, T.; Gospodarek, E. Occurrence and susceptibility to antibiotics of carbapenem-resistant Pseudomonas aeruginosa strains between 1998 and 2009. Med. Dosw. Mikrobiol. 2010, 62, 221–229. [Google Scholar]
- Papagiannitsis, C.C.; Medvecky, M.; Chudejova, K.; Skalova, A.; Rotova, V.; Spanelova, P.; Jakubu, V.; Zemlickova, H.; Hrabak, J. Czech Participants of the European Antimicrobial Resistance Surveillance Network Molecular Characterization of Carbapenemase-Producing Pseudomonas Aeruginosa of Czech Origin and Evidence for Clonal Spread of Extensively Resistant Sequence Type 357 Expressing IMP-7 Metallo-β-Lactamase. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Al-Wrafy, F.; Brzozowska, E.; Górska, S.; Gamian, A. Pathogenic Factors of Pseudomonas Aeruginosa—The Role of Biofilm in Pathogenicity and as a Target for Phage Therapy. Postepy Hig. Med. Dosw. 2017, 71, 78–91. [Google Scholar] [CrossRef]
- Stover, C.K.; Pham, X.Q.; Erwin, A.L.; Mizoguchi, S.D.; Warrener, P.; Hickey, M.J.; Brinkman, F.S.; Hufnagle, W.O.; Kowalik, D.J.; Lagrou, M.; et al. Complete Genome Sequence of Pseudomonas Aeruginosa PAO1, an Opportunistic Pathogen. Nature 2000, 406, 959–964. [Google Scholar] [CrossRef]
- Ramsay, K.A.; Wardell, S.J.T.; Patrick, W.M.; Brockway, B.; Reid, D.W.; Winstanley, C.; Bell, S.C.; Lamont, I.L. Genomic and Phenotypic Comparison of Environmental and Patient-Derived Isolates of Pseudomonas Aeruginosa Suggest That Antimicrobial Resistance Is Rare within the Environment. J. Med. Microbiol. 2019, 68, 1591–1595. [Google Scholar] [CrossRef]
- Van Delden, C.; Iglewski, B.H. Cell-to-Cell Signaling and Pseudomonas Aeruginosa Infections. Emerg. Infect. Dis. 1998, 4, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Ellappan, K.; Belgode Narasimha, H.; Kumar, S. Coexistence of Multidrug Resistance Mechanisms and Virulence Genes in Carbapenem-Resistant Pseudomonas Aeruginosa Strains from a Tertiary Care Hospital in South India. J. Glob. Antimicrob. Resist. 2018, 12, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Morita, Y.; Tomida, J.; Kawamura, Y. Responses of Pseudomonas Aeruginosa to Antimicrobials. Front. Microbiol. 2014, 4, 422. [Google Scholar] [CrossRef] [Green Version]
- Pobiega, M.; Maciąg, J.; Chmielarczyk, A.; Romaniszyn, D.; Pomorska-Wesolowska, M.; Ziolkowski, G.; Heczko, P.B.; Bulanda, M.; Wojkowska-Mach, J. Molecular Characterization of Carbapenem-Resistant Pseudomonas Aeruginosa Strains Isolated from Patients with Urinary Tract Infections in Southern Poland. Diagn. Microbiol. Infect. Dis 2015, 83, 295–297. [Google Scholar] [CrossRef]
- Rossi Gonçalves, I.; Dantas, R.C.C.; Ferreira, M.L.; Batistão, D.W.d.F.; Gontijo-Filho, P.P.; Ribas, R.M. Carbapenem-Resistant Pseudomonas Aeruginosa: Association with Virulence Genes and Biofilm Formation. Braz. J. Microbiol. 2017, 48, 211–217. [Google Scholar] [CrossRef] [Green Version]
- Dehbashi, S.; Tahmasebi, H.; Arabestani, M.R. Association between Beta-Lactam Antibiotic Resistance and Virulence Factors in AmpC Producing Clinical Strains of P. Aeruginosa. Osong Public Health Res. Perspect 2018, 9, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Sawa, T.; Shimizu, M.; Moriyama, K.; Wiener-Kronish, J.P. Association between Pseudomonas Aeruginosa Type III Secretion, Antibiotic Resistance, and Clinical Outcome: A Review. Crit. Care 2014, 18, 668. [Google Scholar] [CrossRef] [Green Version]
- Cho, H.H.; Kwon, K.C.; Kim, S.; Koo, S.H. Correlation between Virulence Genotype and Fluoroquinolone Resistance in Carbapenem-Resistant Pseudomonas Aeruginosa. Ann. Lab. Med. 2014, 34, 286–292. [Google Scholar] [CrossRef] [Green Version]
- Wołkowicz, T.; Patzer, J.A.; Kamińska, W.; Gierczyński, R.; Dzierżanowska, D. Distribution of Carbapenem Resistance Mechanisms in Pseudomonas Aeruginosa Isolates among Hospitalised Children in Poland: Characterisation of Two Novel Insertion Sequences Disrupting the OprD Gene. J. Glob. Antimicrob. Resist. 2016, 7, 119–125. [Google Scholar] [CrossRef]
- Sharifi, H.; Pouladfar, G.; Shakibaie, M.R.; Pourabbas, B.; Mardaneh, J.; Mansouri, S. Prevalence of β-Lactamase Genes, Class 1 Integrons, Major Virulence Factors and Clonal Relationships of Multidrug-Resistant Pseudomonas Aeruginosa Isolated from Hospitalized Patients in Southeast of Iran. Iran. J. Basic Med. Sci. 2019, 22, 806–812. [Google Scholar] [CrossRef]
- Pobiega, M.; Maciag, J.; Pomorska-Wesolowska, M.; Chmielarczyk, A.; Romaniszyn, D.; Ziolkowski, G.; Heczko, P.B.; Wojkowska-Mach, J.; Bulanda, M. Urinary Tract Infections Caused by Pseudomonas Aeruginosa among Children in Southern Poland: Virulence Factors and Antibiotic Resistance. J. Pediatr. Urol. 2016, 12, 36.e1–36.e6. [Google Scholar] [CrossRef]
- Rostami, S.; Farajzadeh Sheikh, A.; Shoja, S.; Farahani, A.; Tabatabaiefar, M.A.; Jolodar, A.; Sheikhi, R. Investigating of Four Main Carbapenem-Resistance Mechanisms in High-Level Carbapenem Resistant Pseudomonas Aeruginosa Isolated from Burn Patients. J. Chin. Med. Assoc. 2018, 81, 127–132. [Google Scholar] [CrossRef]
- Al-Charrakh, A.H.; Al-Awadi, S.J.; Mohammed, A.S. Detection of Metallo-β-Lactamase Producing Pseudomonas Aeruginosa Isolated from Public and Private Hospitals in Baghdad, Iraq. Acta Med. Iran. 2016, 54, 107–113. [Google Scholar]
- Fan, X.; Wu, Y.; Xiao, M.; Xu, Z.-P.; Kudinha, T.; Bazaj, A.; Kong, F.; Xu, Y.-C. Diverse Genetic Background of Multidrug-Resistant Pseudomonas Aeruginosa from Mainland China, and Emergence of an Extensively Drug-Resistant ST292 Clone in Kunming. Sci. Rep. 2016, 6, 26522. [Google Scholar] [CrossRef]
- Al Dawodeyah, H.Y.; Obeidat, N.; Abu-Qatouseh, L.F.; Shehabi, A.A. Antimicrobial Resistance and Putative Virulence Genes of Pseudomonas Aeruginosa Isolates from Patients with Respiratory Tract Infection. Germs 2018, 8, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Amirmozafari, N.; Fallah Mehrabadi, J.; Habibi, A. Association of the Exotoxin A and Exoenzyme S with Antimicrobial Resistance in Pseudomonas Aeruginosa Strains. Arch. Iran. Med. 2016, 19, 353–358. [Google Scholar]
- El-Mahdy, R.; El-Kannishy, G. Virulence Factors of Carbapenem-Resistant Pseudomonas Aeruginosa in Hospital-Acquired Infections in Mansoura, Egypt. Infect. Drug Resist. 2019, 12, 3455–3461. [Google Scholar] [CrossRef] [Green Version]
- Kamali, E.; Jamali, A.; Ardebili, A.; Ezadi, F.; Mohebbi, A. Evaluation of Antimicrobial Resistance, Biofilm Forming Potential, and the Presence of Biofilm-Related Genes among Clinical Isolates of Pseudomonas Aeruginosa. BMC Res. Notes 2020, 13, 27. [Google Scholar] [CrossRef] [PubMed]
- Fuse, K.; Fujimura, S.; Kikuchi, T.; Gomi, K.; Iida, Y.; Nukiwa, T.; Watanabe, A. Reduction of Virulence Factor Pyocyanin Production in Multidrug-Resistant Pseudomonas Aeruginosa. J. Infect. Chemother. 2013, 19, 82–88. [Google Scholar] [CrossRef]
- Liew, S.M.; Rajasekaram, G.; Puthucheary, S.A.; Chua, K.H. Antimicrobial Susceptibility and Virulence Genes of Clinical and Environmental Isolates of Pseudomonas Aeruginosa. PeerJ 2019, 7, e6217. [Google Scholar] [CrossRef] [Green Version]
- Javanmardi, F.; Emami, A.; Pirbonyeh, N.; Keshavarzi, A.; Rajaee, M. A Systematic Review and Meta-Analysis on Exo-Toxins Prevalence in Hospital Acquired Pseudomonas Aeruginosa Isolates. Infect. Genet. Evol. 2019, 75, 104037. [Google Scholar] [CrossRef]
- Khodayary, R.; Nikokar, I.; Mobayen, M.R.; Afrasiabi, F.; Araghian, A.; Elmi, A.; Moradzadeh, M. High Incidence of Type III Secretion System Associated Virulence Factors (Exoenzymes) in Pseudomonas Aeruginosa Isolated from Iranian Burn Patients. BMC Res. Notes 2019, 12, 28. [Google Scholar] [CrossRef] [PubMed]
- Bellés, A.; Bueno, J.; Rojo-Bezares, B.; Torres, C.; Javier Castillo, F.; Sáenz, Y.; Seral, C. Characterisation of VIM-2-Producing Pseudomonas Aeruginosa Isolates from Lower Tract Respiratory Infections in a Spanish Hospital. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 1847–1856. [Google Scholar] [CrossRef]
- Oliver, A.; Mulet, X.; López-Causapé, C.; Juan, C. The Increasing Threat of Pseudomonas Aeruginosa High-Risk Clones. Drug Resist. Updat. 2015, 21–22, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Takata, I.; Yamagishi, Y.; Mikamo, H. Association of the ExoU Genotype with a Multidrug Non-Susceptible Phenotype and MRNA Expressions of Resistance Genes in Pseudomonas Aeruginosa. J. Infect. Chemother. 2018, 24, 45–52. [Google Scholar] [CrossRef]
- Rodulfo, H.; Arcia, A.; Hernández, A.; Michelli, E.; Martinez, D.D.V.; Guzman, M.; Sharma, A.; Donato, M.D. Virulence Factors and Integrons Are Associated with MDR and XDR Phenotypes in Nosocomial Strains of Pseudomonas Aeruginosa in a Venezuelan University Hospital. Rev. Inst. Med. Trop Sao Paulo 2019, 61, e20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruczek, C.; Kottapalli, K.R.; Dissanaike, S.; Dzvova, N.; Griswold, J.A.; Colmer-Hamood, J.A.; Hamood, A.N. Major Transcriptome Changes Accompany the Growth of Pseudomonas Aeruginosa in Blood from Patients with Severe Thermal Injuries. PLoS ONE 2016, 11, e0149229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmassry, M.M.; Mudaliar, N.S.; Kottapalli, K.R.; Dissanaike, S.; Griswold, J.A.; San Francisco, M.J.; Colmer-Hamood, J.A.; Hamood, A.N. Pseudomonas Aeruginosa Alters Its Transcriptome Related to Carbon Metabolism and Virulence as a Possible Survival Strategy in Blood from Trauma Patients. mSystems 2019, 4. [Google Scholar] [CrossRef] [Green Version]
- Golpayegani, A.; Nodehi, R.N.; Rezaei, F.; Alimohammadi, M.; Douraghi, M. Real-Time Polymerase Chain Reaction Assays for Rapid Detection and Virulence Evaluation of the Environmental Pseudomonas Aeruginosa Isolates. Mol. Biol. Rep. 2019, 46, 4049–4061. [Google Scholar] [CrossRef]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.-J.; Cheng, Z. Antibiotic Resistance in Pseudomonas Aeruginosa: Mechanisms and Alternative Therapeutic Strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef]
- Rezzoagli, C.; Archetti, M.; Mignot, I.; Baumgartner, M.; Kümmerli, R. Combining Antibiotics with Antivirulence Compounds Can Have Synergistic Effects and Reverse Selection for Antibiotic Resistance in Pseudomonas Aeruginosa. PLoS Biol. 2020, 18, e3000805. [Google Scholar] [CrossRef]
- Soto-Aceves, M.P.; Cocotl-Yañez, M.; Merino, E.; Castillo-Juárez, I.; Cortés-López, H.; González-Pedrajo, B.; Díaz-Guerrero, M.; Servín-González, L.; Soberón-Chávez, G. Inactivation of the Quorum-Sensing Transcriptional Regulators LasR or RhlR Does Not Suppress the Expression of Virulence Factors and the Virulence of Pseudomonas Aeruginosa PAO1. Microbiology 2019, 165, 425–432. [Google Scholar] [CrossRef]
- Hnamte, S.; Parasuraman, P.; Ranganathan, S.; Ampasala, D.R.; Reddy, D.; Kumavath, R.N.; Suchiang, K.; Mohanty, S.K.; Busi, S. Mosloflavone Attenuates the Quorum Sensing Controlled Virulence Phenotypes and Biofilm Formation in Pseudomonas Aeruginosa PAO1: In Vitro, in Vivo and in Silico Approach. Microb. Pathog. 2019, 131, 128–134. [Google Scholar] [CrossRef]
- Ahmed, S.A.K.S.; Rudden, M.; Smyth, T.J.; Dooley, J.S.G.; Marchant, R.; Banat, I.M. Natural Quorum Sensing Inhibitors Effectively Downregulate Gene Expression of Pseudomonas Aeruginosa Virulence Factors. Appl. Microbiol. Biotechnol. 2019, 103, 3521–3535. [Google Scholar] [CrossRef] [Green Version]
- Kong, W.; Dong, M.; Yan, R.; Liang, Q.; Zhang, H.; Luo, W.; Zhang, Y.; Liang, H.; Duan, K. A Unique ATPase, ArtR (PA4595), Represses the Type III Secretion System in Pseudomonas Aeruginosa. Front. Microbiol. 2019, 10, 560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casciaro, B.; Lin, Q.; Afonin, S.; Loffredo, M.R.; de Turris, V.; Middel, V.; Ulrich, A.S.; Di, Y.P.; Mangoni, M.L. Inhibition of Pseudomonas Aeruginosa Biofilm Formation and Expression of Virulence Genes by Selective Epimerization in the Peptide Esculentin-1a(1-21)NH2. FEBS J. 2019, 286, 3874–3891. [Google Scholar] [CrossRef]
- Islamieh, D.I.; Afshar, D.; Esmaeili, D. Effect of Satureja Khuzistanica Essential Oil (SKEO) Extract on Expression of LasA and LasB Genes in Pseudomonas Aeruginosa. Iran. J. Microbiol. 2019, 11, 55–59. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, M.G.; Vossler, J.L.; Nomura, C.T.; Moffat, J.F. Blocking RpoN Reduces Virulence of Pseudomonas Aeruginosa Isolated from Cystic Fibrosis Patients and Increases Antibiotic Sensitivity in a Laboratory Strain. Sci. Rep. 2019, 9, 6677. [Google Scholar] [CrossRef] [Green Version]
- Mahenthiralingam, E.; Campbell, M.E.; Foster, J.; Lam, J.S.; Speert, D.P. Random Amplified Polymorphic DNA Typing of Pseudomonas Aeruginosa Isolates Recovered from Patients with Cystic Fibrosis. J. Clin. Microbiol. 1996, 34, 1129–1135. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Lim, Y.S.; Yong, D.; Yum, J.H.; Chong, Y. Evaluation of the Hodge Test and the Imipenem-EDTA Double-Disk Synergy Test for Differentiating Metallo-β-Lactamase-Producing Isolates of Pseudomonas Spp. and Acinetobacter Spp. J. Clin. Microbiol. 2003, 41, 4623–4629. [Google Scholar] [CrossRef] [Green Version]
- Yong, D.; Lee, K.; Yum, J.H.; Shin, H.B.; Rossolini, G.M.; Chong, Y. Imipenem-EDTA Disk Method for Differentiation of Metallo-β-Lactamase-Producing Clinical Isolates of Pseudomonas Spp. and Acinetobacter Spp. J. Clin. Microbiol. 2002, 40, 3798–3801. [Google Scholar] [CrossRef] [Green Version]
- Pitout, J.D.D.; Gregson, D.B.; Poirel, L.; McClure, J.-A.; Le, P.; Church, D.L. Detection of Pseudomonas Aeruginosa Producing Metallo-Beta-Lactamases in a Large Centralized Laboratory. J. Clin. Microbiol. 2005, 43, 3129–3135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanotte, P.; Watt, S.; Mereghetti, L.; Dartiguelongue, N.; Rastegar-Lari, A.; Goudeau, A.; Quentin, R. Genetic Features of Pseudomonas Aeruginosa Isolates from Cystic Fibrosis Patients Compared with Those of Isolates from Other Origins. J. Med. Microbiol. 2004, 53, 73–81. [Google Scholar] [CrossRef]
- Finnan, S.; Morrissey, J.P.; O’Gara, F.; Boyd, E.F. Genome Diversity of Pseudomonas Aeruginosa Isolates from Cystic Fibrosis Patients and the Hospital Environment. J. Clin. Microbiol. 2004, 42, 5783–5792. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Subgroup | Gene | phzM | phzS | exoT | exoY | exoU | toxA | algD | exoS * | pilA | pilB |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Carbapenemase (−) | n = 74 | 74 | 74 | 74 | 73 | 73 | 72 | 69 | 57 | 15 | 4 |
| % | 100.0 | 100.0 | 100.0 | 98.6 | 98.6 | 97.3 | 93.2 | 77.0 | 20.3 | 5.4 | |
| Carbapenemase (+) | n = 33 | 33 | 33 | 33 | 33 | 33 | 31 | 30 | 6 | 2 | 1 |
| % | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 93.9 | 90.9 | 18.2 | 6.1 | 3.0 |
| Gene Detected | PCR Primer Name | Manufacturer | Primer Sequence 5′→3′ | Tm (°C) | Annealing Temperature (°C) | Product Size (bp) |
|---|---|---|---|---|---|---|
| phzS | phzS F | Sigma | TCGCCATGACCGATACGCTC | 55.9 | 63 | 1752 |
| phzS R | ACAACCTGAGCCAGCCTTCC | 55.9 | ||||
| exoT | exoT F | AATCGCCGTCCAACTGCATGCG | 58.6 | 64 | 152 | |
| exoT R | TGTTCGCCGAGGTACTGCTC | 55.9 | ||||
| exoY | exoY F | CGGATTCTATGGCAGGGAGG | 55.9 | 64 | 289 | |
| exoY R | GCCCTTGATGCACTCGACCA | 55.9 | ||||
| exoU | exoU F | CCGTTGTGGTGCCGTTGAAG | 55.9 | 64 | 134 | |
| exoU R | CCAGATGTTCACCGACTCGC | 55.9 | ||||
| toxA | toxA F | Integrated DNA Technologies | GGTAACCAGCTCAGCCACAT | 57.4 | 52 | 352 |
| toxA R | TGATGTCCAGGTCATGCTTC | 54.8 | ||||
| algD | algD F | ATGCGAATCAGCATCTTTGGT | 55.2 | 50 | 1310 | |
| algD R | CTACCAGCAGATGCCCTCGGC | 62.5 | ||||
| exoS | exoS F | CTTGAAGGGACTCGACAAGG | 55.2 | 53 | 504 | |
| exoS R | TTCAGGTCCGCGTAGTGAAT | 56.2 | ||||
| phzM | phzM F | Genomed | ATGGAGAGCGGGATCGACAG | 55.9 | 54 | 875 |
| phzM R | ATGCGGGTTTCCATCGGCAG | 55.9 | ||||
| pilA | pilA F | ACAGCATCCAACTGAGCG | 50.3 | 59 | 1675 | |
| pilA R | TTGACTTCCTCCAGGCTG | 50.3 | ||||
| pilB | pilB F | TCGAACTGATGATCGTGG | 48.0 | 56 | 408 | |
| pilB R | CTTTCGGAGTGAACATCG | 48.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bogiel, T.; Depka, D.; Rzepka, M.; Kwiecińska-Piróg, J.; Gospodarek-Komkowska, E. Prevalence of the Genes Associated with Biofilm and Toxins Synthesis amongst the Pseudomonas aeruginosa Clinical Strains. Antibiotics 2021, 10, 241. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10030241
Bogiel T, Depka D, Rzepka M, Kwiecińska-Piróg J, Gospodarek-Komkowska E. Prevalence of the Genes Associated with Biofilm and Toxins Synthesis amongst the Pseudomonas aeruginosa Clinical Strains. Antibiotics. 2021; 10(3):241. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10030241
Chicago/Turabian StyleBogiel, Tomasz, Dagmara Depka, Mateusz Rzepka, Joanna Kwiecińska-Piróg, and Eugenia Gospodarek-Komkowska. 2021. "Prevalence of the Genes Associated with Biofilm and Toxins Synthesis amongst the Pseudomonas aeruginosa Clinical Strains" Antibiotics 10, no. 3: 241. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10030241