
Climatic Alterations Influence Bacterial Growth, Biofilm Production and Antimicrobial Resistance Profiles in Aeromonas spp.

 ,
,
Abstract
:1. Introduction
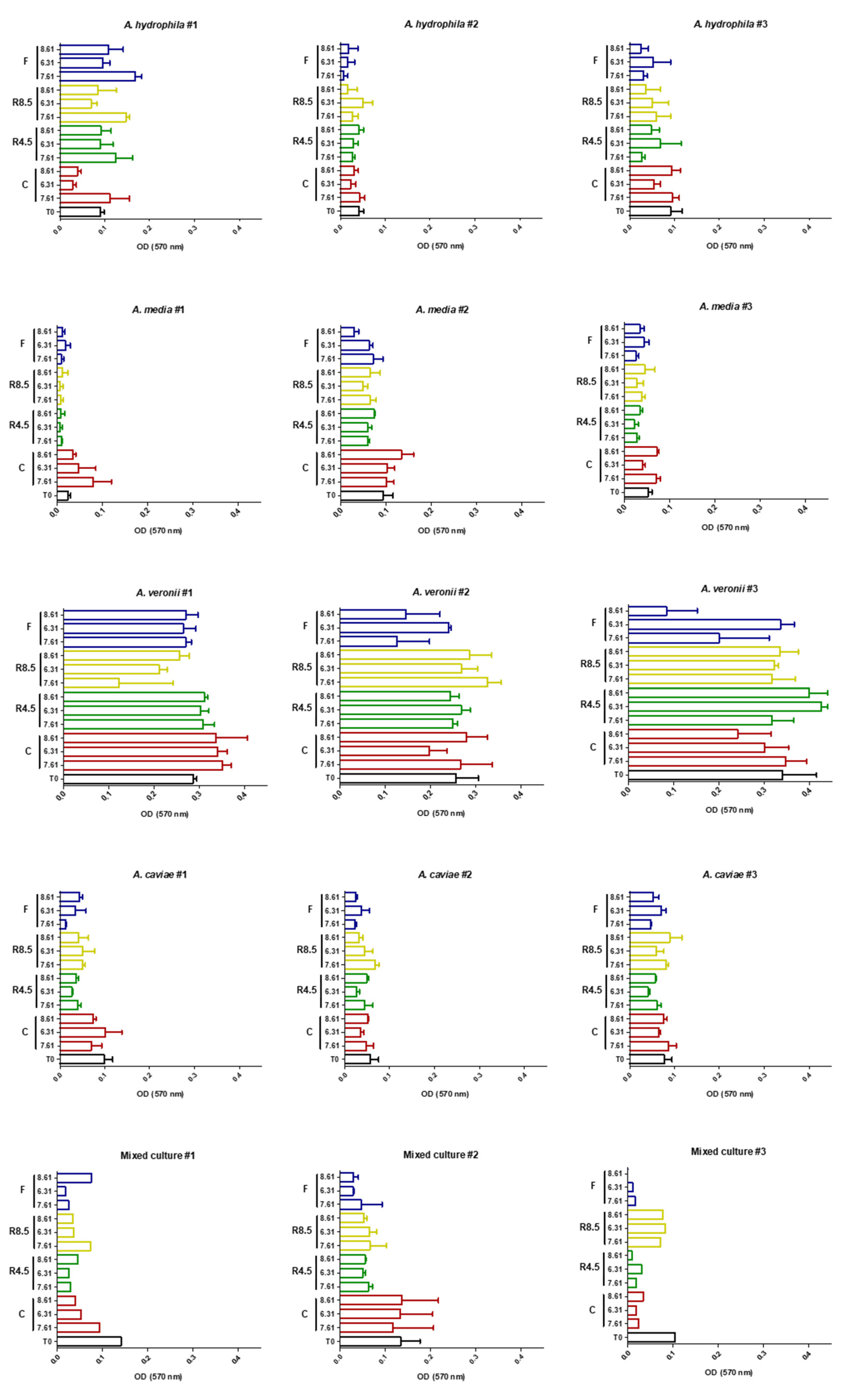
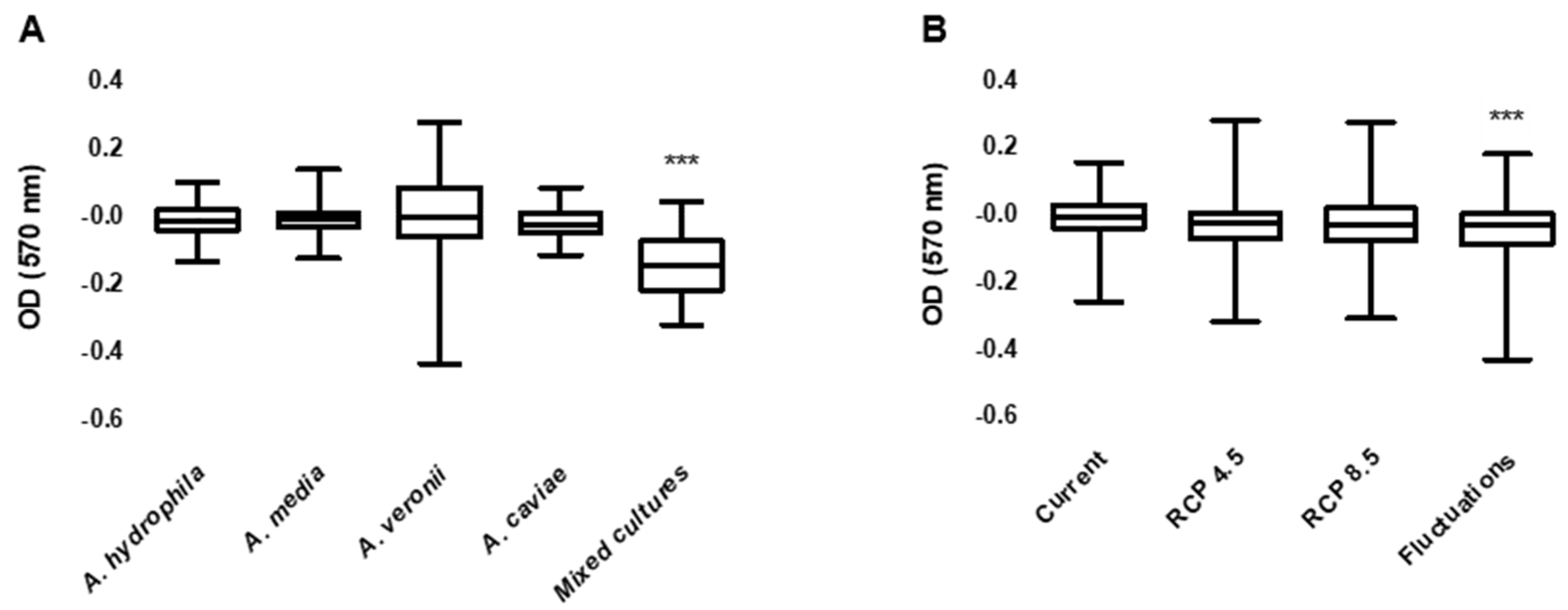
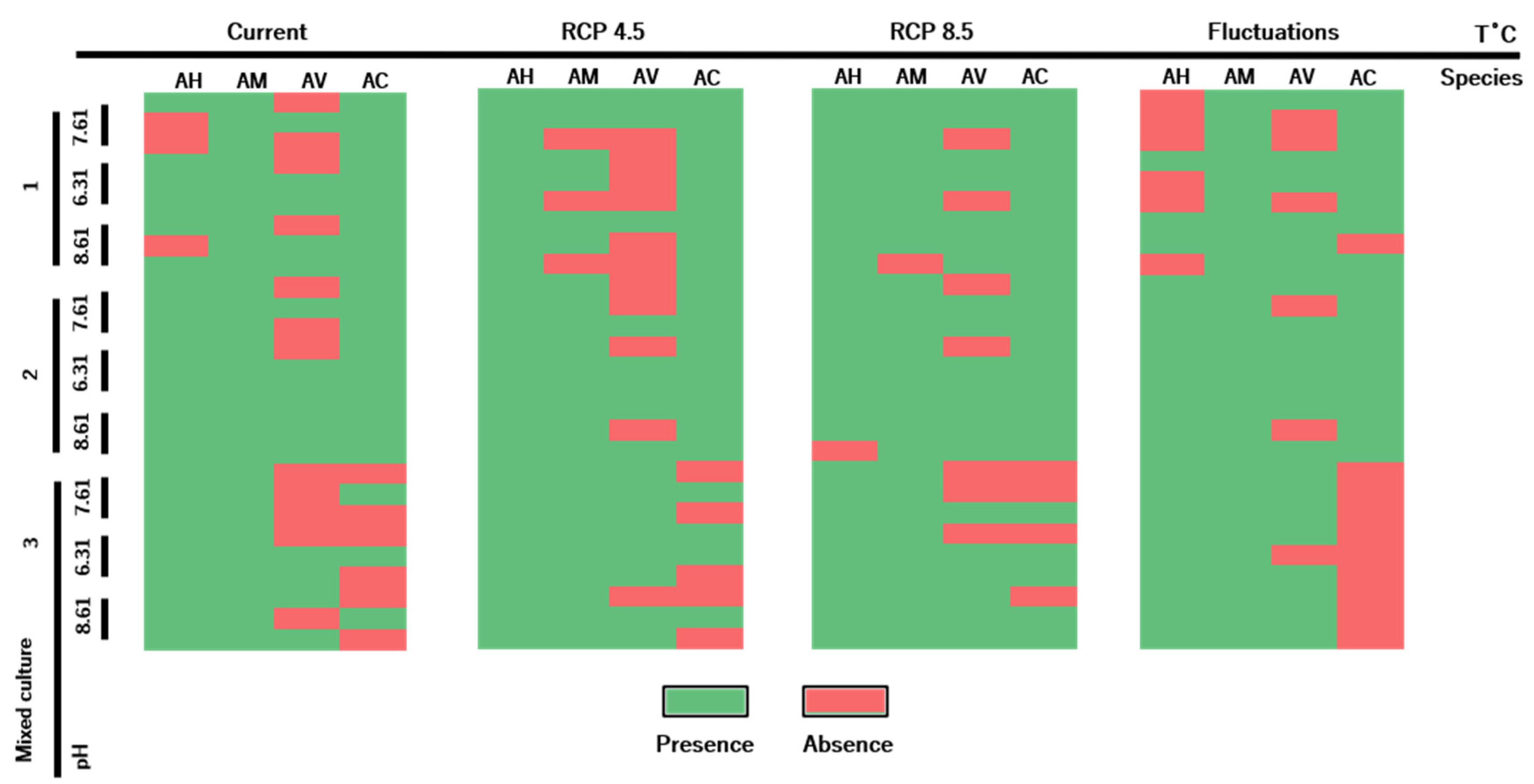
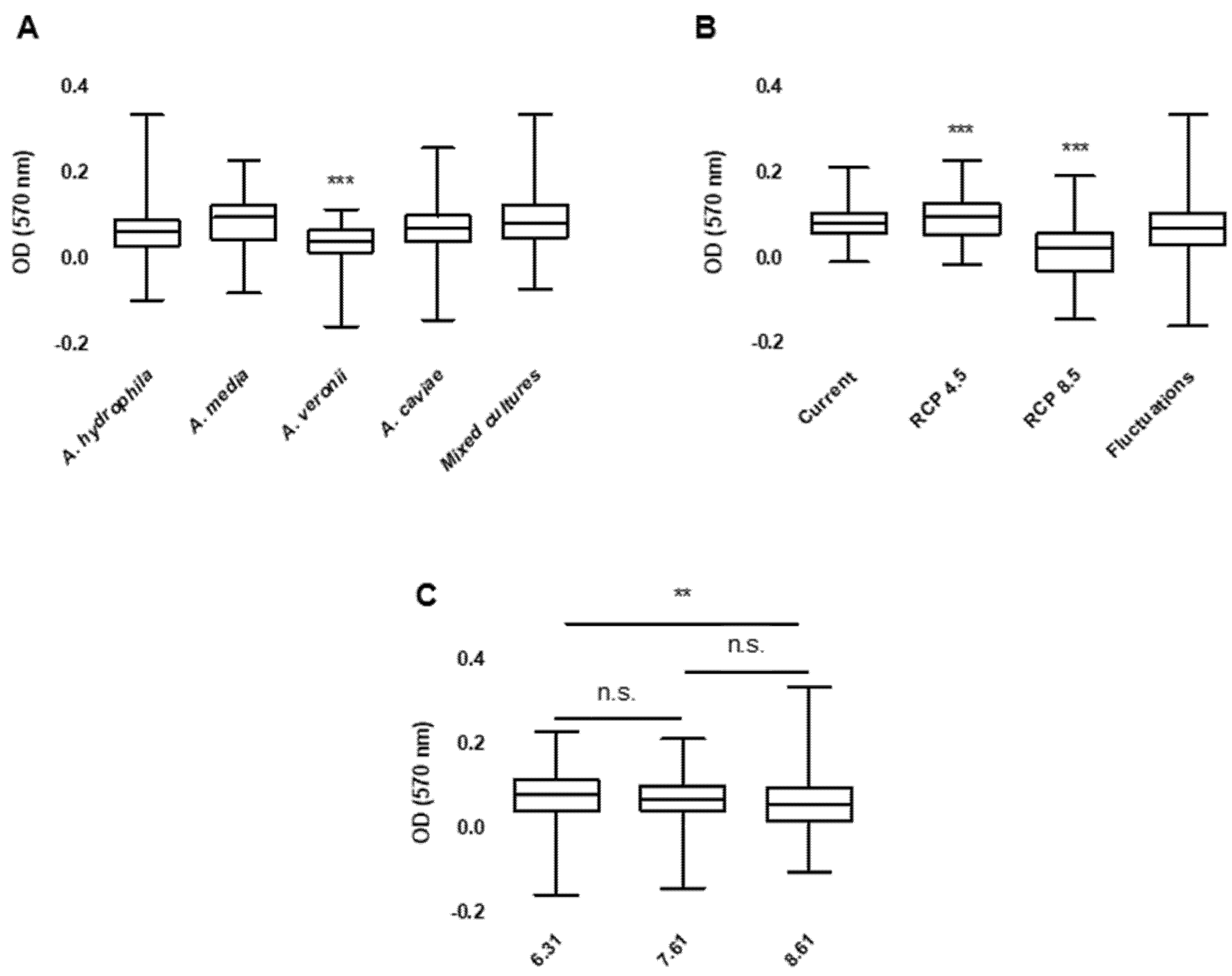
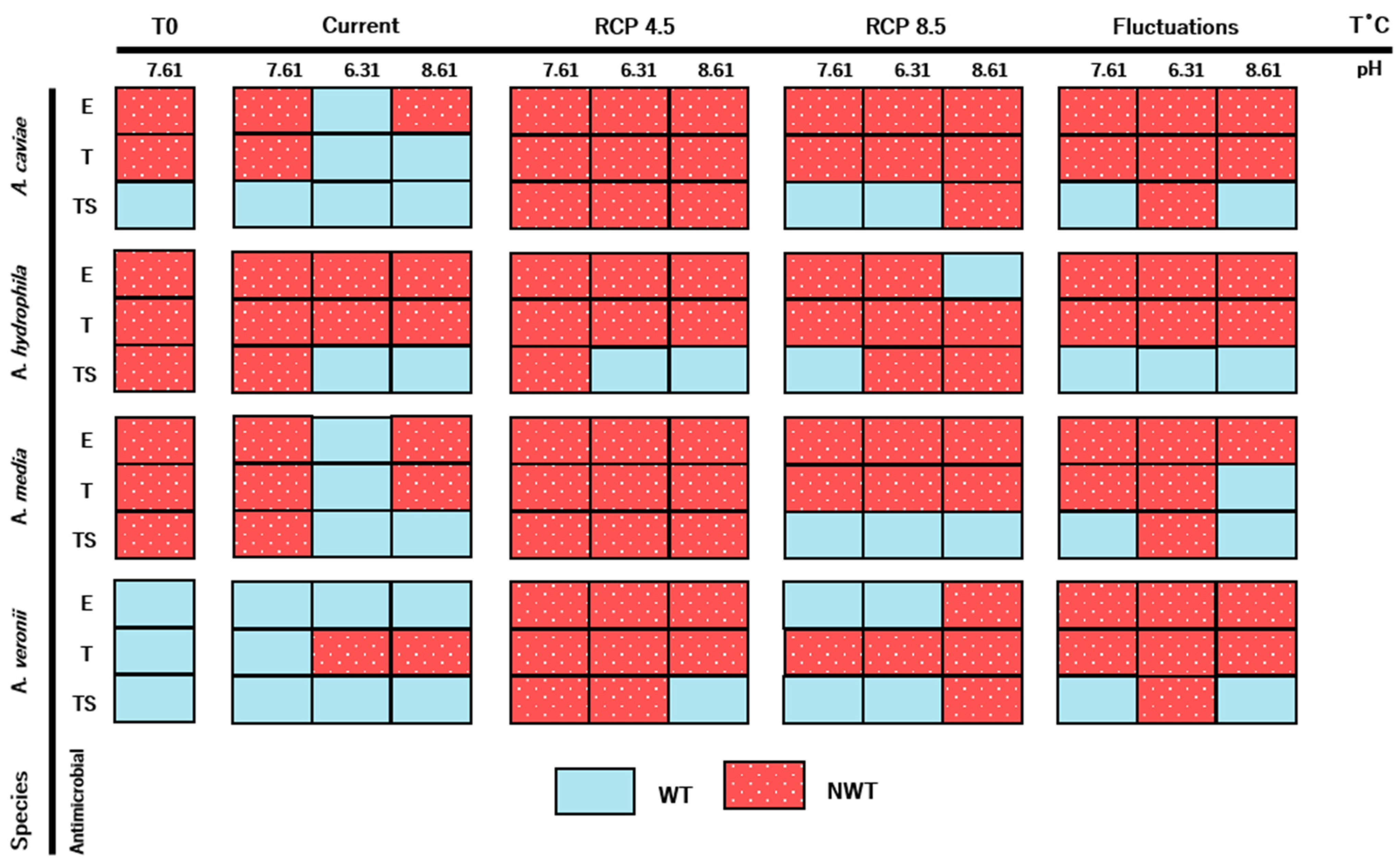
2. Results
3. Discussion
3.1. Biofilm Production
3.2. Bacterial Growth
3.3. Antimicrobial Resistance Profiles
4. Materials and Methods
4.1. Strain Selection
4.2. Biofilm Formation Quantification
4.3. Antimicrobial Susceptibility Testing
4.4. Microcosm Assay
4.5. Aeromonas Species Confirmation in Mixed Culture Wells
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Parter, M.; Kashtan, N.; Alon, U. Environmental variability and modularity of bacterial metabolic networks. BMC Evol. Biol. 2007, 7, 169. [Google Scholar] [CrossRef] [Green Version]
- Ratzke, C.; Gore, J. Modifying and reacting to the environmental pH can drive bacterial interactions. PLoS Biol. 2018, 16, e2004248. [Google Scholar] [CrossRef] [Green Version]
- De Silva, P.M.; Chong, P.; Fernando, D.M.; Westmacott, G.; Kumar, A. Effect of incubation temperature on antibiotic resistance and virulence factors of Acinetobacter baumannii ATCC 17978. Antimicrob. Agents Chemother. 2018, 62, e01514-17. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Liu, W.; Jiang, Q.; Zuo, Y.; Su, Y.; Zhao, L.; Qin, Y.; Yan, Q. Integration of transcriptomic and proteomic approaches reveals the temperature-dependent virulence of Pseudomonas plecoglossicida. Front. Cell. Infect. Microbiol. 2018, 8, 207. [Google Scholar] [CrossRef] [Green Version]
- Pachauri, R.K.; Allen, M.R.; Barros, V.R.; Broome, J.; Cramer, W.; Christ, R.; Church, J.A.; Clarke, L.; Dahe, Q.; Dasgupta, P.; et al. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Pachauri, R., Meyer, L., Eds.; IPCC: Geneva, Switzerland, 2014; p. 151. [Google Scholar]
- Knouft, J.H.; Ficklin, D. The potential impacts of climate change on biodiversity in flowing freshwater systems. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 111–133. [Google Scholar] [CrossRef]
- Van Vliet, M.T.H.; Ludwig, F.; Zwolsman, J.J.G.; Weedon, G.P.; Kabat, P. Global river temperatures and sensitivity to atmospheric warming and changes in river flow. Water Resour. Res. 2011, 47. [Google Scholar] [CrossRef]
- Bucak, T.; Trolle, D.; Andersen, H.E.; Thodsen, H.; Erdoğan, S.; Levi, E.E.; Filiz, N.; Jeppesen, E.; Beklioğlu, M. Future water availability in the largest freshwater Mediterranean lake is at great risk as evidenced from simulations with the SWAT model. Sci. Total Environ. 2017, 581–582, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Griffith, A.W.; Gobler, C.J. Harmful algal blooms: A climate change co-stressor in marine and freshwater ecosystems. Harmful Algae 2020, 91, 101590. [Google Scholar] [CrossRef] [PubMed]
- Kaushal, S.S.; Likens, G.E.; Pace, M.L.; Utz, R.; Haq, S.; Gorman, J.; Grese, M. Freshwater salinization syndrome on a continental scale. Proc. Natl. Acad. Sci. USA 2018, 115, E574–E583. [Google Scholar] [CrossRef] [Green Version]
- Le, T.D.H.; Kattwinkel, M.; Schützenmeister, K.; Olson, J.R.; Hawkins, C.P.; Schäfer, R.B. Predicting current and future background ion concentrations in German surface water under climate change. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dudgeon, D. Multiple threats imperil freshwater biodiversity in the Anthropocene. Curr. Biol. 2019, 29, R960–R967. [Google Scholar] [CrossRef]
- Mosley, L. Drought impacts on the water quality of freshwater systems; review and integration. Earth Sci. Rev. 2015, 140, 203–214. [Google Scholar] [CrossRef]
- Van Vliet, M.T.; Franssen, W.H.; Yearsley, J.R.; Ludwig, F.; Haddeland, I.; Lettenmaier, D.P.; Kabat, P. Global river discharge and water temperature under climate change. Glob. Environ. Chang. 2013, 23, 450–464. [Google Scholar] [CrossRef]
- Pinceel, T.; Buschke, F.; Weckx, M.; Brendonck, L.; Vanschoenwinkel, B. Climate change jeopardizes the persistence of freshwater zooplankton by reducing both habitat suitability and demographic resilience. BMC Ecol. 2018, 18, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodell, M.; Famiglietti, J.; Wiese, D.N.; Reager, J.T.; Beaudoing, H.K.; Landerer, F.W.; Lo, M.-H. Emerging trends in global freshwater availability. Nature 2018, 557, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Jing, L.; Teng, Y.; Wang, J. Characterization of antibiotics in a large-scale river system of China: Occurrence pattern, spatiotemporal distribution and environmental risks. Sci. Total Environ. 2018, 618, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Peterson, E.; Kaur, P. Antibiotic resistance mechanisms in bacteria: Relationships between resistance determinants of antibiotic producers, environmental bacteria, and clinical pathogens. Front. Microbiol. 2018, 9, 2928. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.; Bollmann, A.; Seitz, W.; Schwartz, T. Microbiological characterization of aquatic microbiomes targeting taxonomical marker genes and antibiotic resistance genes of opportunistic bacteria. Sci. Total Environ. 2015, 512–513, 316–325. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.F.; da Rocha Fernandes, G.; de Paiva, M.C.; Salim, A.C.D.M.; Santos, A.B.; Nascimento, A.M.A. Exploring the resistome, virulome and microbiome of drinking water in environmental and clinical settings. Water Res. 2020, 174, 115630. [Google Scholar] [CrossRef]
- Friman, V.-P.; Hiltunen, T.; Jalasvuori, M.; Lindstedt, C.; Laanto, E.; Örmälä, A.-M.; Laakso, J.; Mappes, J.; Bamford, J.K.H. High Temperature and bacteriophages can indirectly select for bacterial pathogenicity in environmental reservoirs. PLoS ONE 2011, 6, e17651. [Google Scholar] [CrossRef] [PubMed]
- Saarinen, K.; Lindström, L.; Ketola, T. Invasion triple trouble: Environmental fluctuations, fluctuation-adapted invaders and fluctuation-mal-adapted communities all govern invasion success. BMC Evol. Biol. 2019, 19, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbott, S.L.; Cheung, W.K.; Janda, J.M. The genus aeromonas: Biochemical characteristics, atypical reactions, and phenotypic identification schemes. J. Clin. Microbiol. 2003, 41, 2348–2357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Gu, J. Influence of temperature, salinity and pH on the growth of environmental Aeromonas and Vibrio species isolated from Mai Po and the Inner Deep Bay Nature Reserve Ramsar Site of Hong Kong. J. Basic Microbiol. 2005, 45, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Awan, F.; Dong, Y.; Wang, N.; Liu, J.; Ma, K.; Liu, Y. The fight for invincibility: Environmental stress response mechanisms and Aeromonas hydrophila. Microb. Pathog. 2018, 116, 135–145. [Google Scholar] [CrossRef]
- Thomas, L.V.; Wimpenny, J.W. Competition between Salmonella and Pseudomonas species growing in and on agar, as affected by pH, sodium chloride concentration and temperature. Int. J. Food Microbiol. 1996, 29, 361–370. [Google Scholar] [CrossRef]
- Lopez-Vazquez, C.M.; Oehmen, A.; Hooijmans, C.M.; Brdjanovic, D.; Gijzen, H.J.; Yuan, Z.; van Loosdrecht, M.C. Modeling the PAO-GAO competition: Effects of carbon source, pH and temperature. Water Res. 2009, 43, 450–462. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Meng, F. Ecological insights into the underlying evolutionary patterns of biofilm formation from biological wastewater treatment systems: Red or black queen hypothesis? Biotechnol. Bioeng. 2020, 117, 1270–1280. [Google Scholar] [CrossRef]
- Delamare, A.P.L.; Costa, S.O.P.; Da Silveira, M.M.; Echeverrigaray, S. Growth of Aeromonas species on increasing concentrations of sodium chloride. Lett. Appl. Microbiol. 2000, 30, 57–60. [Google Scholar] [CrossRef] [Green Version]
- Nana, G.Y.G.; Ripoll, C.; Cabin-Flaman, A.; Gibouin, D.; Delaune, A.; Janniere, L.; Grancher, G.; Chagny, G.; Loutelier-Bourhis, C.; Lentzen, E.; et al. Division-based, growth rate diversity in bacteria. Front. Microbiol. 2018, 9, 849. [Google Scholar] [CrossRef]
- Goelzer, A.; Fromion, V. Bacterial growth rate reflects a bottleneck in resource allocation. Biochim. Biophys. Acta 2011, 1810, 978–988. [Google Scholar] [CrossRef]
- Cavari, B.Z.; Allen, D.; Colwell, R.R. Effect of temperature on growth and activity of Aeromonas spp. and mixed bacterial populations in the Anacostia river. Appl. Environ. Microbiol. 1981, 41, 1052–1054. [Google Scholar] [CrossRef] [Green Version]
- Sautour, M.; Mary, P.; Chihib, N.; Hornez, J. The effects of temperature, water activity and pH on the growth of Aeromonas hydrophila and on its subsequent survival in microcosm water. J. Appl. Microbiol. 2003, 95, 807–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palumbo, S.A.; Morgan, D.R.; Buchanan, R.L. Influence of temperature, NaCI, and pH on the growth of Aeromonas hydrophila. J. Food Sci. 1985, 50, 1417–1421. [Google Scholar] [CrossRef]
- Knøchel, S. Growth characteristics of motile Aeromonas spp. isolated from different environments. Int. J. Food Microbiol. 1990, 10, 235–244. [Google Scholar] [CrossRef]
- Vivekanandhan, G.; Savithamani, K.; Lakshmanaperumalsamy, P. Influence of pH, salt concentration and temperature on the growth of Aeromonas hydrophila. J. Environ. Biol. 2003, 24, 373–379. [Google Scholar] [PubMed]
- Karem, K.L.; Foster, J.W.; Bej, A.K. Adaptive acid tolerance response (ATR) in Aeromonas hydrophila. Microbiology 1994, 140, 1731–1736. [Google Scholar] [CrossRef] [Green Version]
- Buncic, S.; Avery, S.M. Effect of pre-incubation pH on the growth characteristics of Aeromonas hydrophila at 5 °C, as assessed by two methods. Lett. Appl. Microbiol. 1995, 20, 7–10. [Google Scholar] [CrossRef]
- Alvarez-Uria, G.; Midde, M. Trends and factors associated with antimicrobial resistance of Acinetobacter spp. invasive isolates in Europe: A country-level analysis. J. Glob. Antimicrob. Resist. 2018, 14, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Kaba, H.E.; Kuhlmann, E.; Scheithauer, S. Thinking outside the box: Association of antimicrobial resistance with climate warming in Europe—A 30 country observational study. Int. J. Hyg. Environ. Health 2020, 223, 151–158. [Google Scholar] [CrossRef]
- Reverter, M.; Sarter, S.; Caruso, D.; Avarre, J.-C.; Combe, M.; Pepey, E.; Pouyaud, L.; Vega-Heredía, S.; De Verdal, H.; Gozlan, R.E. Aquaculture at the crossroads of global warming and antimicrobial resistance. Nat. Commun. 2020, 11, 70. [Google Scholar] [CrossRef] [Green Version]
- MacFadden, D.R.; McGough, S.F.; Fisman, D.; Santillana, M.; Brownstein, J.S. Antibiotic resistance increases with local temperature. Nat. Clim. Chang. 2018, 8, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Andersson, D.I.; Hughes, D. Antibiotic resistance and its cost: Is it possible to reverse resistance? Nat. Rev. Microbiol. 2010, 8, 260–271. [Google Scholar] [CrossRef]
- Sundqvist, M. Reversibility of antibiotic resistance. Upsala J. Med Sci. 2014, 119, 142–148. [Google Scholar] [CrossRef] [Green Version]
- Melnyk, A.H.; Wong, A.; Kassen, R. The fitness costs of antibiotic resistance mutations. Evol. Appl. 2015, 8, 273–283. [Google Scholar] [CrossRef]
- Hernando-Amado, S.; Sanz-García, F.; Blanco, P.; Martínez, J.L. Fitness costs associated with the acquisition of antibiotic resistance. Essays Biochem. 2017, 61, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Berger, D.; Stångberg, J.; Baur, J.; Walters, R.J. Elevated temperature increases genome-wide selection on de novo mutations. Proc. R. Soc. B 2021, 288, 20203094. [Google Scholar] [CrossRef] [PubMed]
- Freihofer, P.; Akbergenov, R.; Teo, Y.; Juskeviciene, R.; Andersson, D.I.; Böttger, E.C. Nonmutational compensation of the fitness cost of antibiotic resistance in mycobacteria by overexpression of tlyA rRNA methylase. RNA 2016, 22, 1836–1843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fay, A.; Philip, J.; Saha, P.; Hendrickson, R.C.; Glickman, M.S.; Burns-Huang, K. The DnaK chaperone system buffers the fitness cost of antibiotic resistance mutations in mycobacteria. mBio 2021, 12, e00123-21. [Google Scholar] [CrossRef] [PubMed]
- McMahon, M.A.S.; Xu, J.; Moore, J.E.; Blair, I.S.; McDowell, D.A. Environmental stress and antibiotic resistance in food-related pathogens. Appl. Environ. Microbiol. 2007, 73, 211–217. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Verdugo, A.; Gaut, B.S.; Tenaillon, O. Evolution of Escherichia coli rifampicin resistance in an antibiotic-free environment during thermal stress. BMC Evol. Biol. 2013, 13, 50. [Google Scholar] [CrossRef] [Green Version]
- Grilo, M.; Isidoro, S.; Chambel, L.; Marques, C.; Marques, T.; Sousa-Santos, C.; Robalo, J.; Oliveira, M. Molecular epidemiology, virulence traits and antimicrobial resistance signatures of Aeromonas spp. in the critically endangered Iberochondrostoma lusitanicum follow geographical and seasonal patterns. Antibiotics 2021, 10, 759. [Google Scholar] [CrossRef]
- Sousa-Santos, C.; Robalo, J.I.; Pereira, A.M.; Branco, P.; Santos, J.M.; Ferreira, M.T.; Sousa, M.; Doadrio, I. Broad-scale sampling of primary freshwater fish populations reveals the role of intrinsic traits, inter-basin connectivity, drainage area and latitude on shaping contemporary patterns of genetic diversity. PeerJ 2016, 4, e1694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freeman, D.J.; Falkiner, F.R.; Keane, C.T. New method for detecting slime production by coagulase negative staphylococci. J. Clin. Pathol. 1989, 42, 872–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stepanović, S.; Vuković, D.; Dakić, I.; Savić, B.; Švabić-Vlahović, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Stepanović, S.; Vukovic, D.; Hola, V.; Di Bonaventura, G.; Djukić, S.; Ćirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Turck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute—CLSI. Performance Standards for Antimicrobial Susceptibility Testing of Bacteria Isolated From Aquatic Animals; 2nd Informational Supplement; CLSI document VET03/VET04-S2; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2014. [Google Scholar]
- Goller, C.C.; Romeo, T. Environmental influences on biofilm development. In Bacterial biofilms. Current Topics in Microbiology and Immunology; Romeo, T., Ed.; Springer: Berlin, Germany, 2008; p. 322. [Google Scholar]
- Toyofuku, M.; Inaba, T.; Kiyokawa, T.; Obana, N.; Yawata, Y.; Nomura, N. Environmental factors that shape biofilm formation. Biosci. Biotechnol. Biochem. 2016, 80, 7–12. [Google Scholar] [CrossRef]
- Cruz-Loya, M.; Kang, T.M.; Lozano, N.A.; Watanabe, R.; Tekin, E.; Damoiseaux, R.; Savage, V.M.; Yeh, P.J. Stressor interaction networks suggest antibiotic resistance co-opted from stress responses to temperature. ISME J. 2018, 13, 12–23. [Google Scholar] [CrossRef]
- Mueller, E.; Egan, A.J.; Breukink, E.; Vollmer, W.; Levin, P.A. Plasticity of Escherichia coli cell wall metabolism promotes fitness and antibiotic resistance across environmental conditions. eLife 2019, 8, e40754. [Google Scholar] [CrossRef]
- SNIRH. Sistema Nacional de Informação de Recursos Hídricos. 2021. Available online: https://snirh.apambiente.pt/index.php?idMain=2&idItem=1 (accessed on 22 June 2020). (In Portuguese)
- Kaushal, S.S.; Likens, G.E.; Utz, R.M.; Pace, M.L.; Grese, M.; Yepsen, M. Increased river alkalinization in the Eastern U.S. Environ. Sci. Technol. 2013, 47, 10302–10311. [Google Scholar] [CrossRef]
- Oberholster, P.; Botha, A.-M.; Hill, L.; Strydom, W. River catchment responses to anthropogenic acidification in relationship with sewage effluent: An ecotoxicology screening application. Chemosphere 2017, 189, 407–417. [Google Scholar] [CrossRef]
- Zhang, Q.-G.; Buckling, A. Phages limit the evolution of bacterial antibiotic resistance in experimental microcosms. Evol. Appl. 2012, 5, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Cairns, J.; Becks, L.; Jalasvuori, M.; Hiltunen, T. Sublethal streptomycin concentrations and lytic bacteriophage together promote resistance evolution. Philos. Trans. R. Soc. B 2017, 372, 20160040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talon, D.; Mulin, B.; Thouverez, M. Clonal identification of Aeromonas hydrophila strains using randomly amplified polymorphic DNA analysis. Eur. J. Epidemiol. 1998, 14, 305–310. [Google Scholar] [CrossRef]
- Persson, S.; Al-Shuweli, S.; Yapici, S.; Jensen, J.N.; Olsen, K.E.P. Identification of clinical Aeromonas species by rpoB and gyrb sequencing and development of a multiplex PCR method for detection of Aeromonas hydrophila, A. caviae, A. veronii, and A. media. J. Clin. Microbiol. 2014, 53, 653–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Conditions | |||
|---|---|---|---|
| Temperature (°C) | pH | ||
| Current | 21 | Current | 7.61 |
| RCP 4.5 | 23.2 | Acidification | 6.31 |
| RCP 8.5 | 24.5 | Alkalization | 8.61 |
| Fluctuations | 21–24.5 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grilo, M.L.; Pereira, A.; Sousa-Santos, C.; Robalo, J.I.; Oliveira, M. Climatic Alterations Influence Bacterial Growth, Biofilm Production and Antimicrobial Resistance Profiles in Aeromonas spp. Antibiotics 2021, 10, 1008. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10081008
Grilo ML, Pereira A, Sousa-Santos C, Robalo JI, Oliveira M. Climatic Alterations Influence Bacterial Growth, Biofilm Production and Antimicrobial Resistance Profiles in Aeromonas spp. Antibiotics. 2021; 10(8):1008. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10081008
Chicago/Turabian StyleGrilo, Miguel L., Ana Pereira, Carla Sousa-Santos, Joana I. Robalo, and Manuela Oliveira. 2021. "Climatic Alterations Influence Bacterial Growth, Biofilm Production and Antimicrobial Resistance Profiles in Aeromonas spp." Antibiotics 10, no. 8: 1008. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10081008