
Anti-Virulence Activity of the Cell-Free Supernatant of the Antarctic Bacterium Psychrobacter sp. TAE2020 against Pseudomonas aeruginosa Clinical Isolates from Cystic Fibrosis Patients

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Results
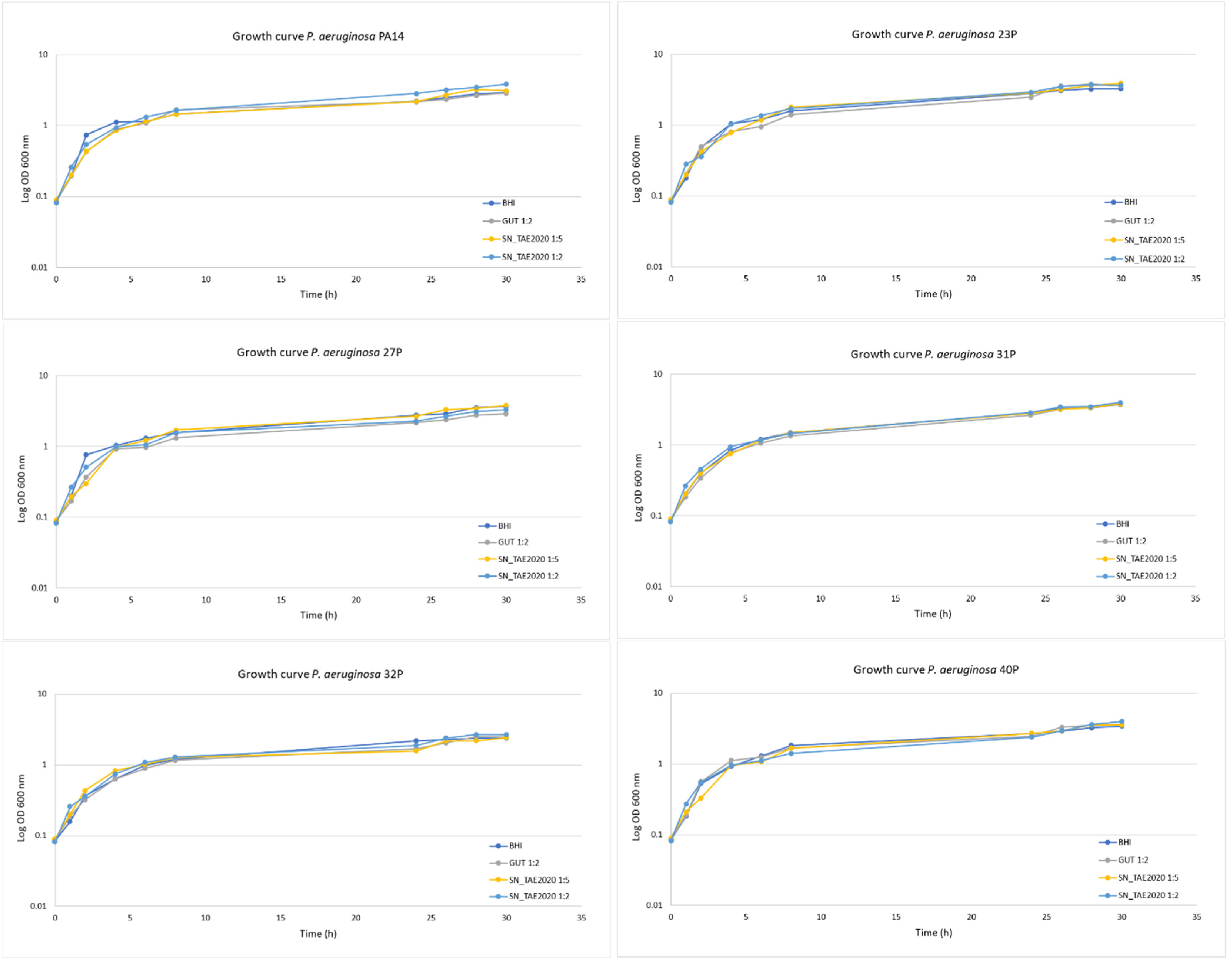
2.1. Phenotypic Characterization of Clinical and PA14 Strains
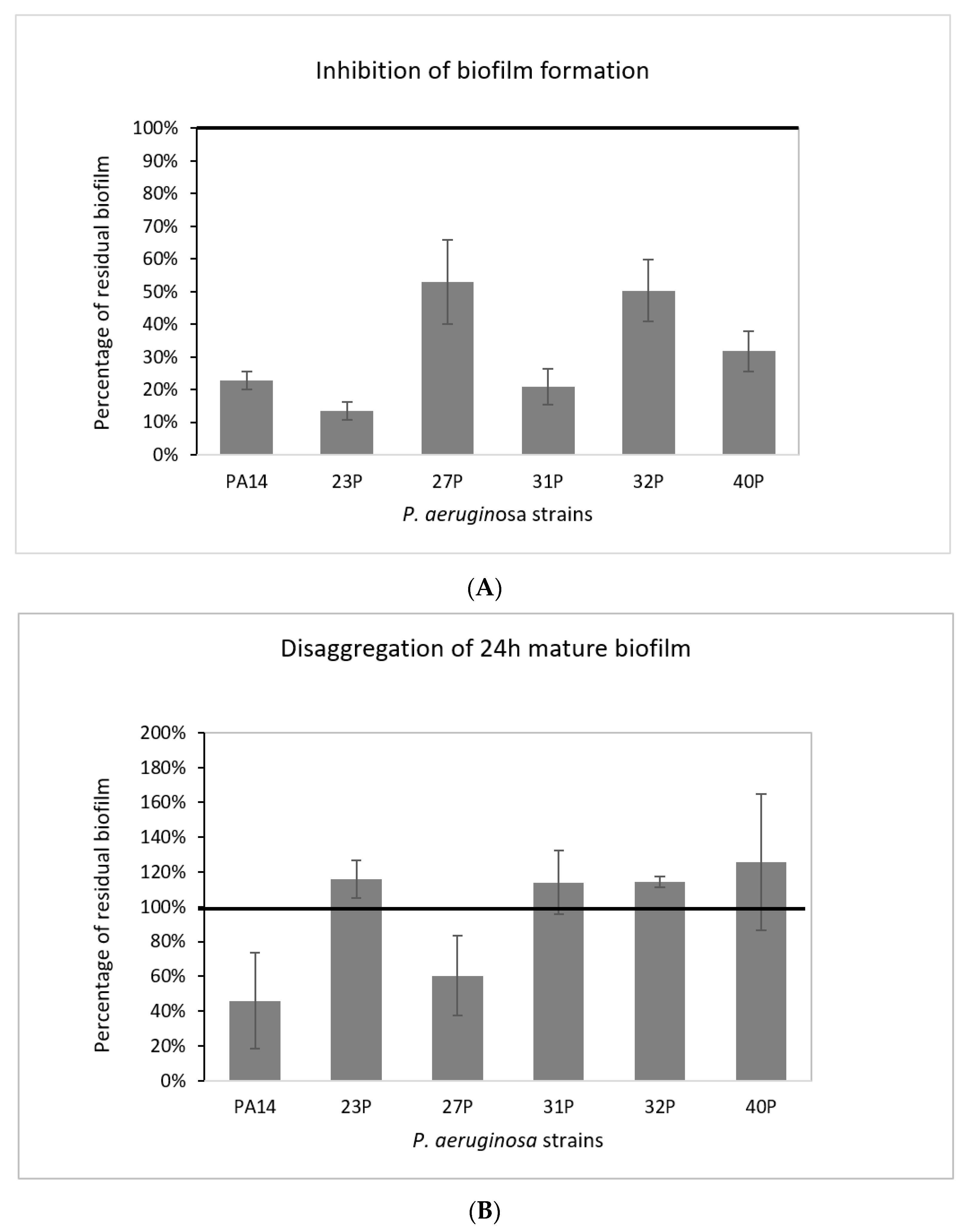
2.2. Effect of SN_TAE2020 on Biofilm Formation
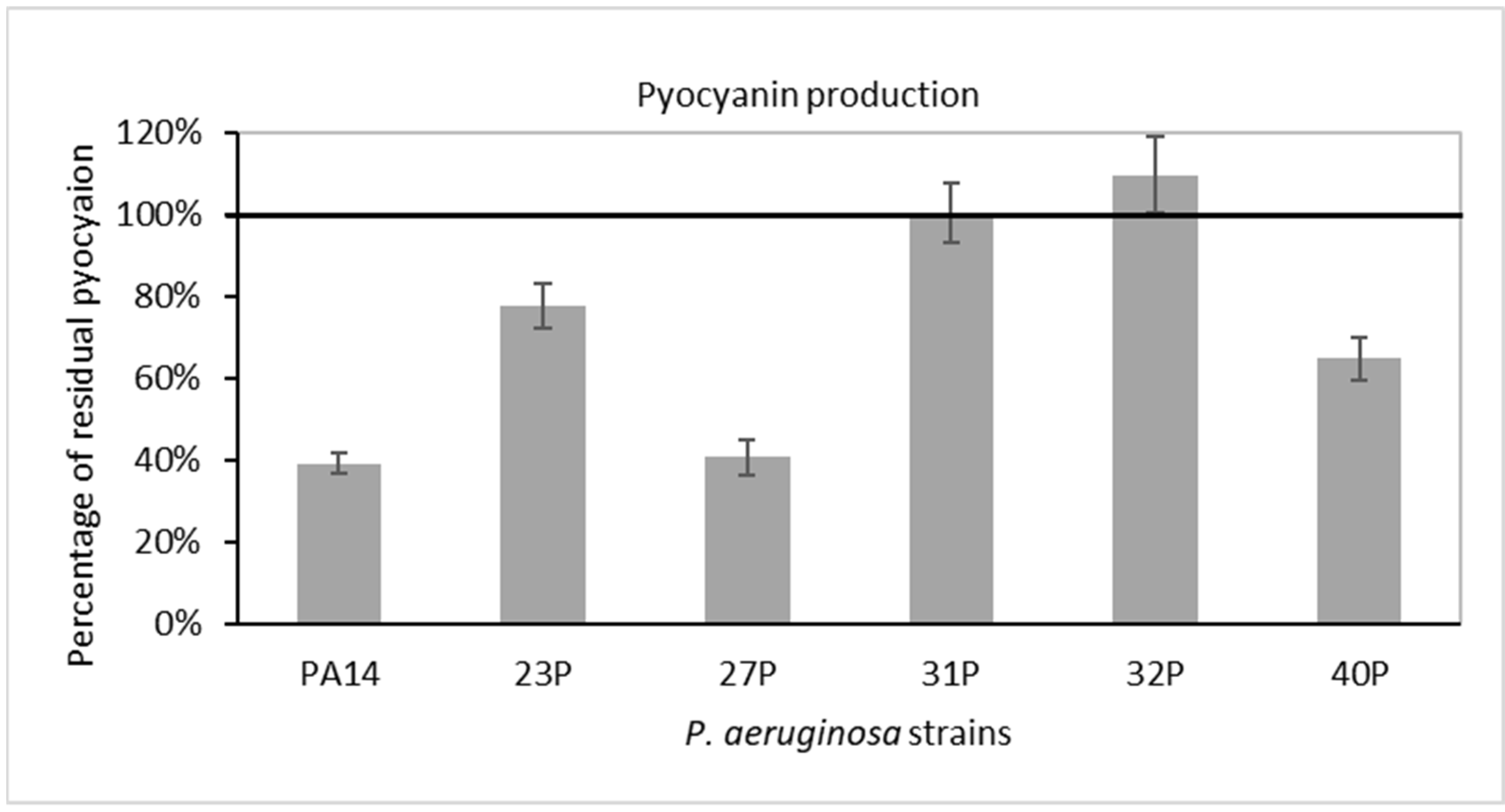
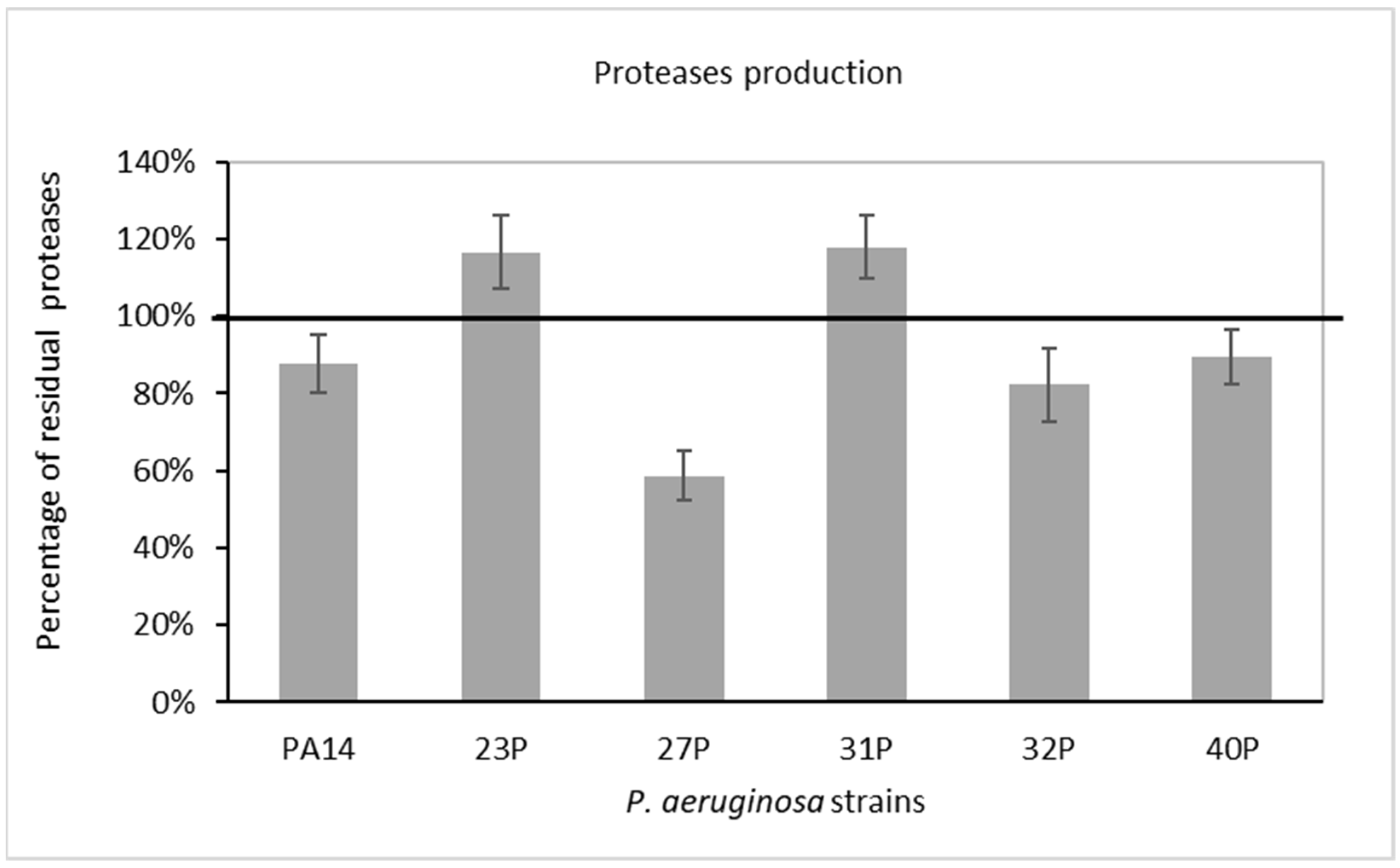
2.3. Effect of SN_TAE2020 on Pyocyanin Production and Protease Activity
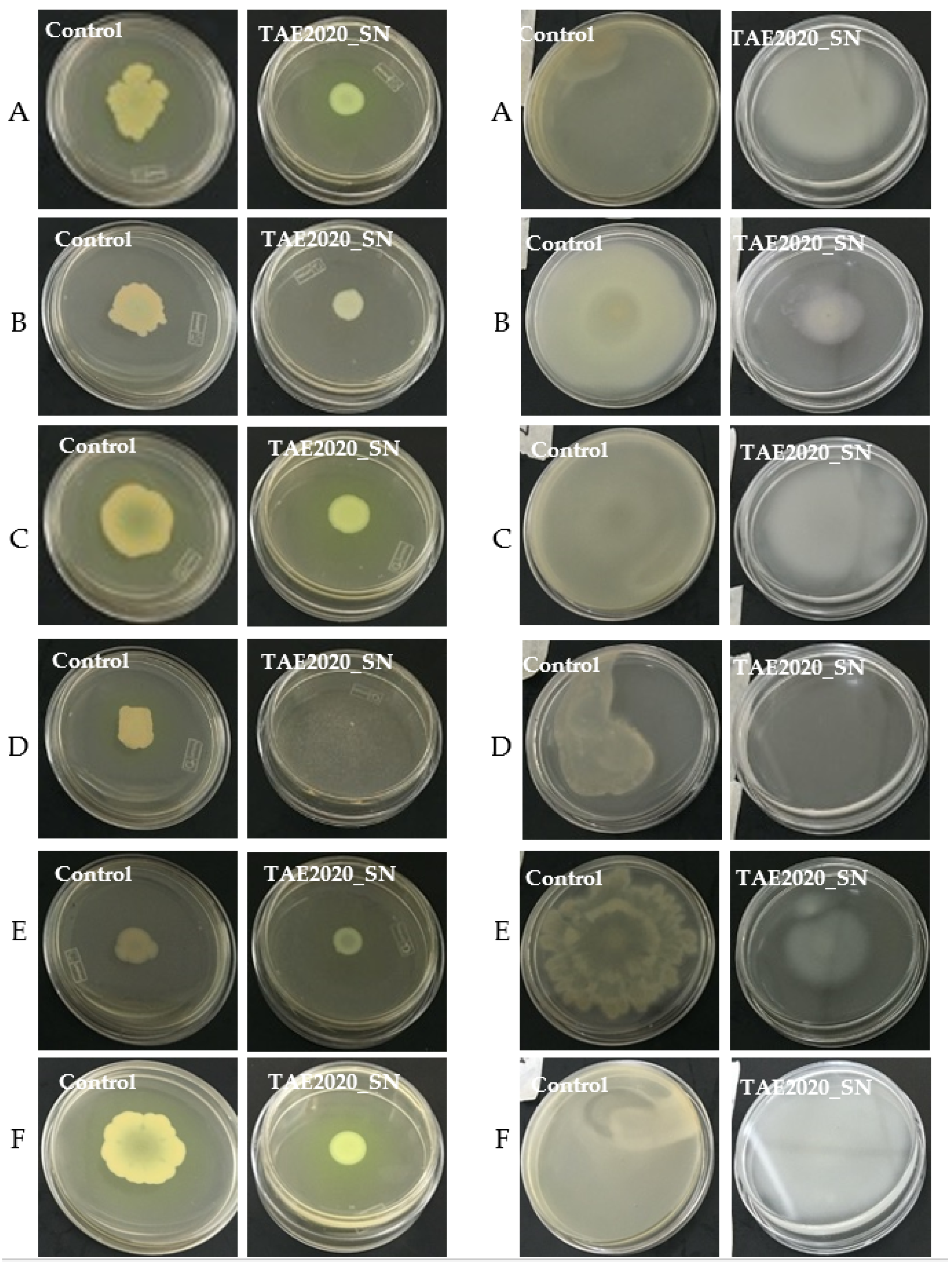
2.4. Effect of SN_TAE2020 on Motility
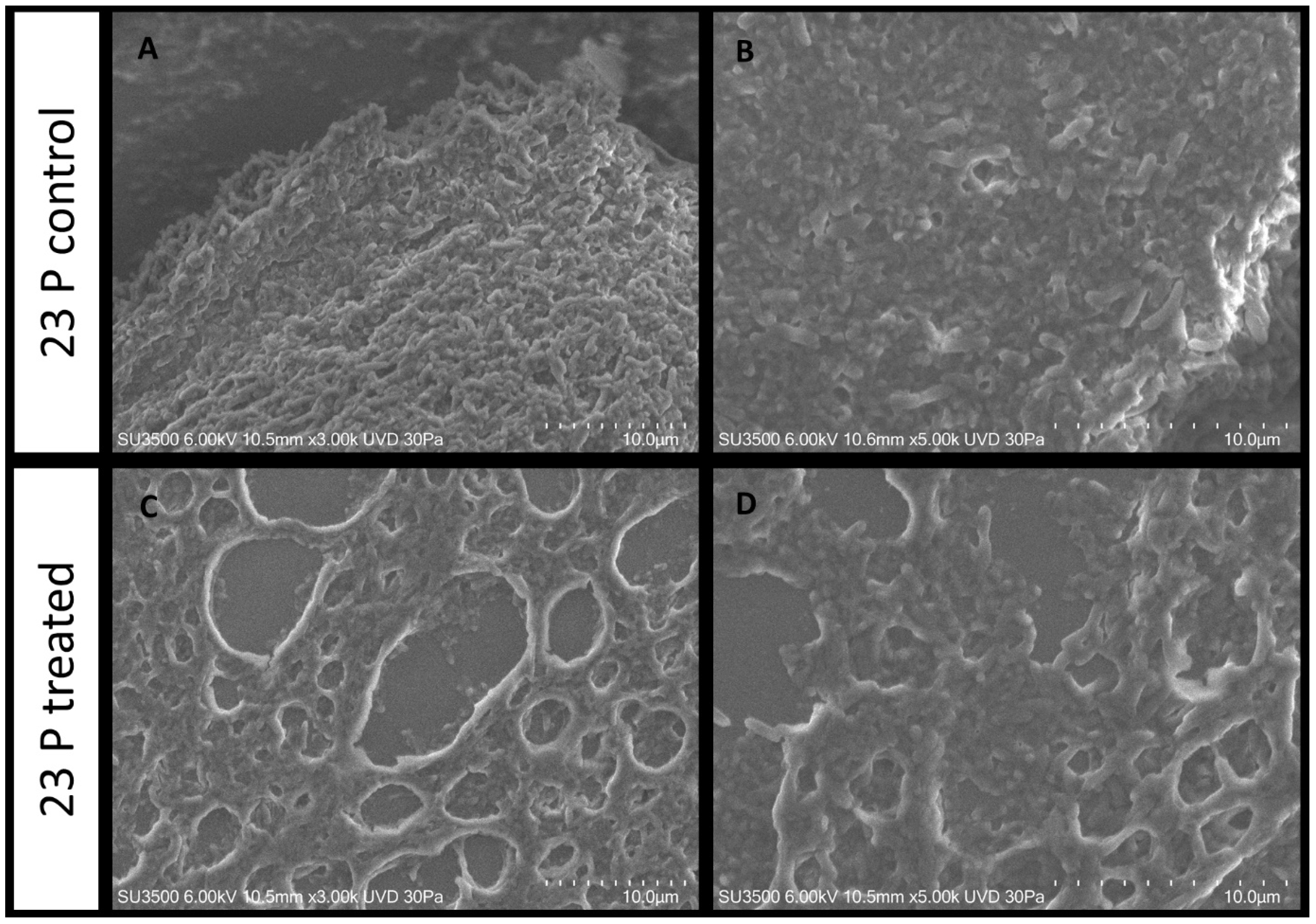
2.5. Morphological Evaluation by SEM Analysis
2.5.1. Evaluation of SN_TAE2020’s Influence on P. aeruginosa PA14
2.5.2. Evaluation of SN_TAE2020 on P. aeruginosa 23P
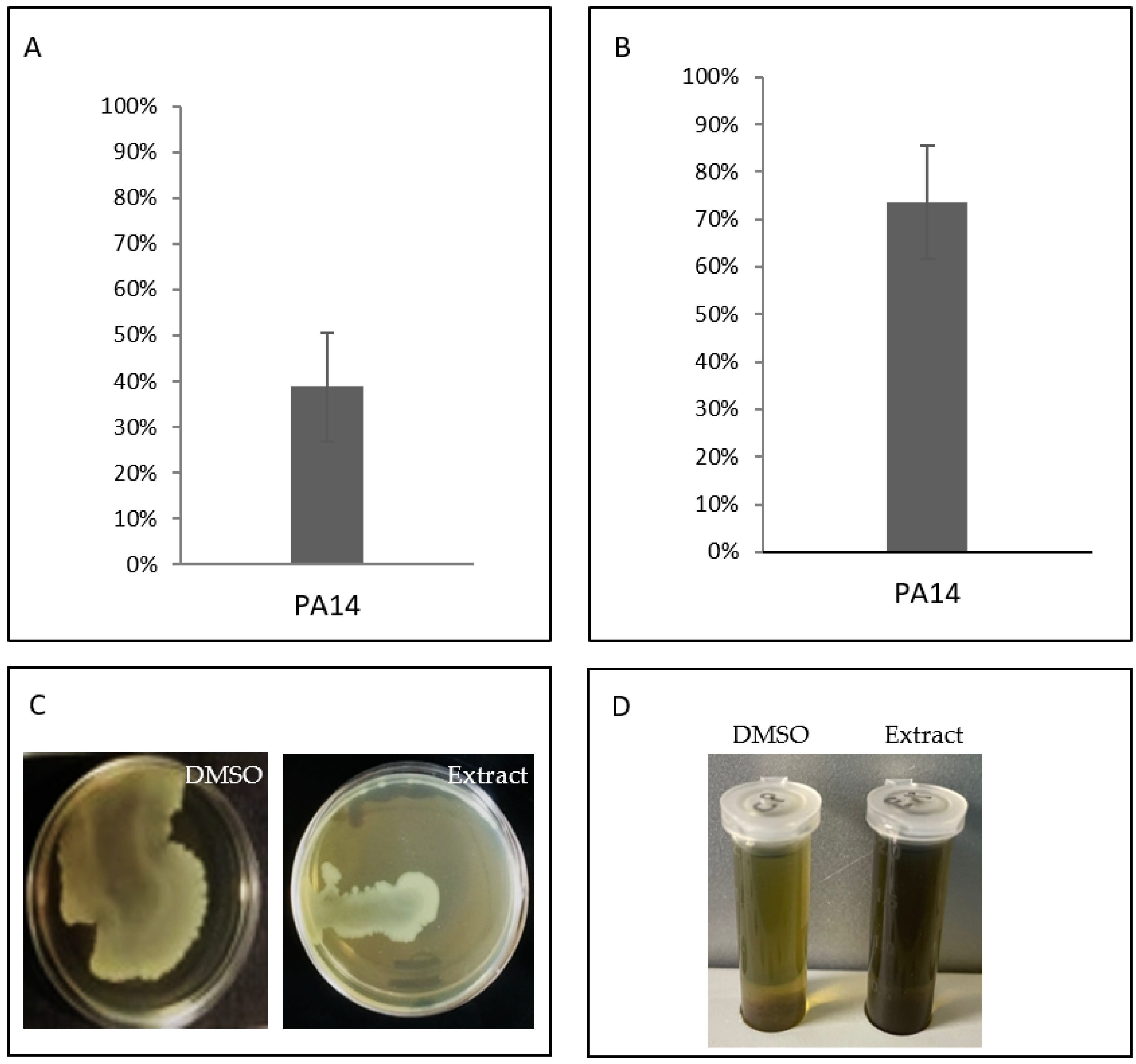
2.6. Anti-Virulence Features of Organic Extract from the Supernatant of TAE2020
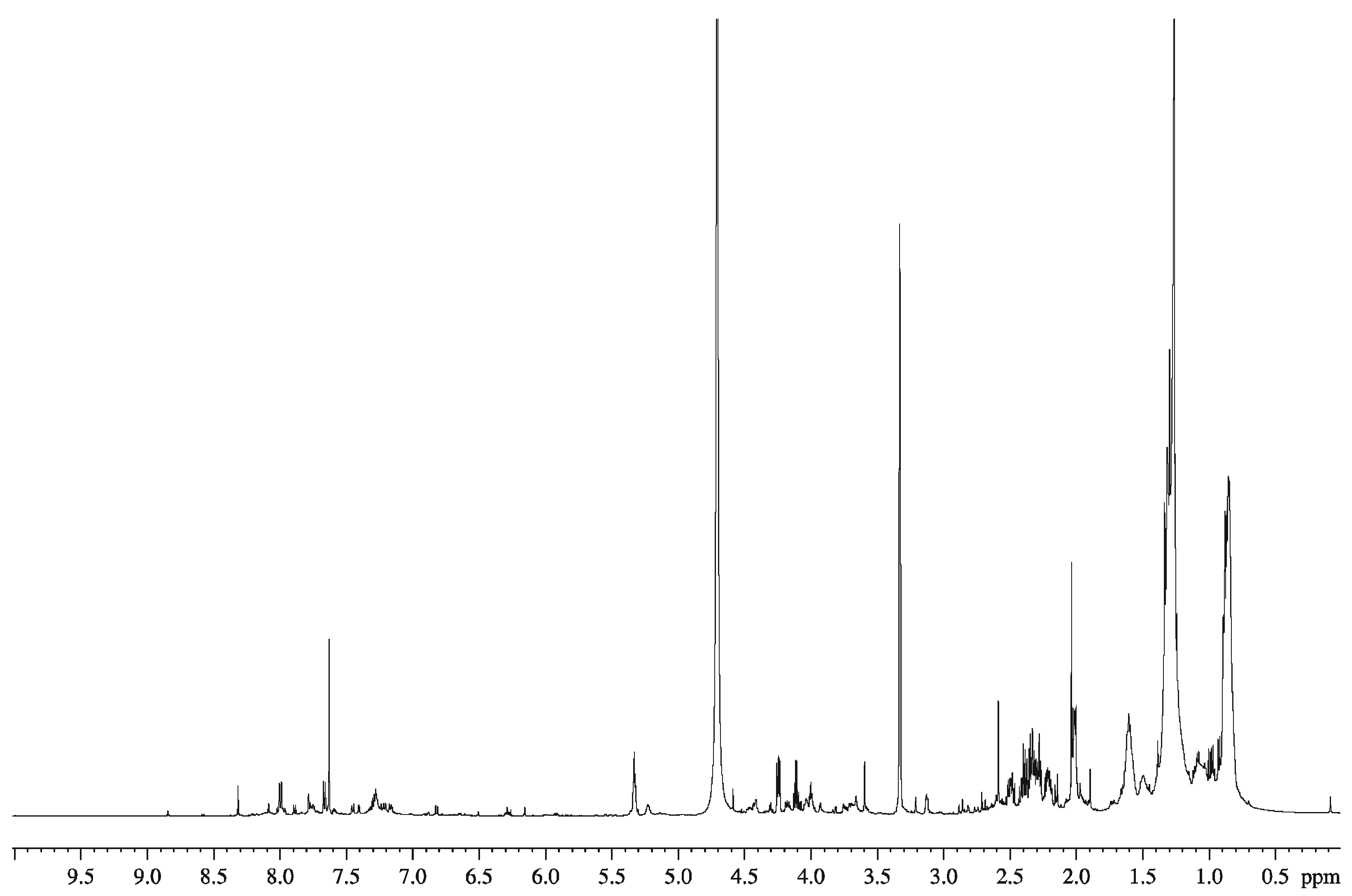
2.7. GC-MS Analysis of the Organic Extract from SN_TAE2020
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
4.2. Pre-Adhesion Period
4.3. Mature Biofilm
4.4. Pyocyanin Assay
4.5. Protease Assay
4.6. Motility Assays
4.6.1. Swarming Assay
4.6.2. Swimming Assay
4.7. SEM Analysis
4.8. SN_TAE2020 Supernatant Preparation and Organic Extraction Protocol
4.9. GC-MS of SN_TAE2020
4.10. NMR Analysis of SN_TAE2020
4.11. Mass Spectrometry Analysis of SN_TAE2020
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rimessi, A.; Vitto, V.A.M.; Patergnani, S.; Pinton, P. Update on Calcium signaling in cystic fibrosis lung disease. Front. Pharm. 2021, 12, 581645. [Google Scholar] [CrossRef]
- Rosales-Reyes, R.; Vargas-Roldán, S.Y.; Lezana-Fernández, J.L.; Santos-Preciado, J.I. Pseudomonas aeruginosa: Genetic adaptation, a strategy for its persistence in cystic fibrosis. Arch. Med. Res. 2020, S0188–4409, 32245–32251. [Google Scholar]
- O’Sullivan, B.P.; Freedman, S.D. Cystic fibrosis. Lancet 2009, 373, 1891–1904. [Google Scholar] [CrossRef]
- Sousa, A.M.; Pereira, M.O. Pseudomonas aeruginosa diversification during infection development in cystic fibrosis Lungs—A review. Pathogens 2014, 3, 680–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gellatly, S.L.; Hancock, R.E. Pseudomonas aeruginosa: New insights into pathogenesis and host defenses. Pathog. Dis. 2013, 67, 159–173. [Google Scholar] [CrossRef] [Green Version]
- Castillo-Juarez, I.; Lopez-Jacome, L.E.; Soberon-Chavez, G.; Tomas, M.; Lee, J.; Castaneda-Tamez, P.; Hernández-Bárragan, I.Á.; Cruz-Muñiz, M.Y.; Maeda, T.; Wood, T.K.; et al. Exploiting quorum sensing inhibition for the control of Pseudomonas aeruginosa and Acinetobacter baumannii biofilms. Curr. Top. Med. Chem. 2017, 17, 1915–1927. [Google Scholar] [CrossRef]
- Pendleton, J.N.; Gorman, S.P.; Gilmore, B.F. Clinical relevance of the ESKAPE pathogens. Expert Rev. Anti Infect. 2013, 11, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Winstanley, C.; O’Brien, S.; Brockhurst, M.A. Pseudomonas aeruginosa evolutionary adaptation and diversification in cystic fibrosis chronic lung infections. Trends Microbiol. 2016, 24, 327–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pejčić, M.; Stojanović-Radić, Z.; Genčić, M.; Dimitrijević, M.; Radulović, N. Anti-virulence potential of basil and sage essential oils: Inhibition of biofilm formation, motility and pyocyanin production of Pseudomonas aeruginosa isolates. Food Chem. Toxicol. 2020, 141, 111431. [Google Scholar] [CrossRef]
- Lee, K.; Yoon, S.S. Pseudomonas aeruginosa biofilm, a programmed bacterial life for fitness. J. Microbiol. Biotechnol. 2017, 27, 1053–1064. [Google Scholar] [CrossRef] [Green Version]
- O’May, C.; Tufenkji, N. The swarming motility of Pseudomonas aeruginosa is blocked by cranberry proanthocyanidins and other tannin-containing materials. Appl. Env. Microbiol. 2011, 77, 3061–3067. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.W.; Yu, S.; Zhang, Z.Y.; Wei, Q.; Yan, L.; Ai, G.M.; Liu, H.; Ma, L.Z. Coordination of swarming motility, biosurfactant synthesis, and biofilm matrix exopolysaccharide production in Pseudomonas aeruginosa. Appl. Env. Microbiol. 2014, 80, 6724–6732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alatraktchi, F.A.; Svendsen, W.E.; Molin, S. Electrochemical detection of pyocyanin as a biomarker for Pseudomonas aeruginosa: A focused review. Sensors 2020, 20, 5218. [Google Scholar] [CrossRef]
- Schauder, S.; Bassler, B.L. The languages of bacteria. Genes Dev. 2001, 15, 1468–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, J.Y.; Chen, T.T.; Tan, X.J.; Chen, T.; Jia, A.Q. The quorum-sensing inhibiting effects of stilbenoids and their potential structure-activity relationship. Bioorg. Med. Chem. Lett. 2015, 25, 5217–5220. [Google Scholar] [CrossRef]
- Rasko, D.A.; Sperandio, V. Anti-virulence strategies to combat bacteria-mediated disease. Nat. Rev. Drug Discov. 2010, 9, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Hentzer, M.; Givskov, M. Pharmacological inhibition of quorum sensing for the treatment of chronic bacterial infections. J. Clin. Investig. 2003, 112, 1300–1307. [Google Scholar] [CrossRef]
- Pattnaik, S.; Ahmed, T.; Ranganathan, S.K.; Ampasala, D.R.; Sarma, V.V.; Busi, S. Aspergillus ochraceopetaliformis SSP13 modulates quorum sensing regulated virulence and biofilm formation in Pseudomonas aeruginosa PAO1. Biofouling 2018, 34, 410–425. [Google Scholar] [CrossRef]
- Artini, M.; Patsilinakos, A.; Papa, R.; Božović, M.; Sabatino, M.; Garzoli, S.; Vrenna, G.; Tilotta, M.; Pepi, F.; Ragno, R.; et al. Antimicrobial and antibiofilm activity and machine learning classification analysis of essential oils from different Mediterranean plants against Pseudomonas aeruginosa. Molecules 2018, 23, 482. [Google Scholar] [CrossRef] [Green Version]
- Papa, R.; Selan, L.; Parrilli, E.; Tilotta, M.; Sannino, F.; Feller, G.; Tutino, M.L.; Artini, M. Anti-biofilm activities from marine cold adapted bacteria against Staphylococci and Pseudomonas aeruginosa. Front. Microbiol. 2015, 6, 1333. [Google Scholar] [CrossRef] [Green Version]
- Elmanama, A.A.; Al-Reefi, M.R. Antimicrobial, anti-biofilm, anti- quorum sensing, antifungal and synergistic effects of some medicinal plants extracts. Iug. J. Nat. Eng. Stud. 2017, 25, 198–207. [Google Scholar]
- Zhou, J.; Bi, S.; Chen, H.; Chen, T.; Yang, R.; Li, M.; Fu, Y.; Jia, A.Q. Anti-biofilm and antivirulence activities of metabolites from Plectosphaerella cucumerina against Pseudomonas aeruginosa. Front. Microbiol 2017, 8, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, T.B.; Cheung, R.C.; Wong, J.H.; Bekhit, A.A.; Bekhit, A.E.D. Antibacterial products of marine organisms. Appl. Microbiol. Biotechnol. 2015, 99, 4145–4173. [Google Scholar] [CrossRef]
- Katz, L.; Baltz, R.H. Natural product discovery: Past, present, and future. J. Ind. Microbiol. Biotechnol. 2016, 43, 155–176. [Google Scholar] [CrossRef] [PubMed]
- Núñez-Montero, K.; Barrientos, L. Advances in Antarctic research for antimicrobial discovery: A comprehensive narrative review of bacteria from Antarctic environments as potential sources of novel antibiotic compounds against human pathogens and microorganisms of industrial importance. Antibiotics 2018, 7, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikkelsen, H.; McMullan, R.; Filloux, A. The Pseudomonas aeruginosa reference strain PA14 displays increased virulence due to a mutation in ladS. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [Green Version]
- Rashiya, N.; Padmini, N.; Ajilda, A.A.K.; Prabakaran, P.; Durgadevi, R.; Veera Ravi, A.; Ghosh, S.; Sivakumar, N.; Selvakumar, G. Inhibition of biofilm formation and quorum sensing mediated virulence in Pseudomonas aeruginosa by marine sponge symbiont Brevibacterium casei strain Alu 1. Microb. Pathog. 2021, 150, 104693. [Google Scholar] [CrossRef] [PubMed]
- Artini, M.; Papa, R.; Vrenna, G.; Lauro, C.; Ricciardelli, A.; Casillo, A.; Corsaro, M.M.; Tutino, M.L.; Parrilli, E.; Selan, L. Cold-adapted bacterial extracts as a source of anti-infective and antimicrobial compounds against Staphylococcus aureus. Future Microbiol. 2019, 14, 1369. [Google Scholar] [CrossRef] [PubMed]
- Starkey, M.; Lepine, F.; Maura, D.; Bandyopadhaya, A.; Lesic, B.; He, J.; Kitao, T.; Righi, V.; Milot, S.; Tzika, A.; et al. Identification of anti-virulence compounds that disrupt quorum-sensing regulated acute and persistent pathogenicity. PloS Pathog. 2014, 10. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.; Zhang, L.; Kirienko, N.V. High-Throughput Approaches for the Identification of Pseudomonas aeruginosa antivirulents. mBio 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Moreau-Marquis, S.; Stanton, B.A.; O’Toole, G.A. Pseudomonas aeruginosa biofilm formation in the cystic fibrosis airway. Pulm. Pharm. 2008, 21, 595–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoodley, P.; Sauer, K.; Davies, D.G.; Costerton, J.W. Biofilms as complex differentiated communities. Annu. Rev. Microbiol. 2002, 56, 187–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muh, U.; Schuster, M.; Heim, R.; Singh, A.; Olson, E.R.; Greenberg, E.P. Novel Pseudomonas aeruginosa quorum-sensing inhibitors identified in an ultra-high-throughput screen. Antimicrob. Agents Chemother. 2006, 50, 3674–3679. [Google Scholar] [CrossRef] [Green Version]
- Jurado-Martín, I.; Sainz-Mejías, M.; McClean, S. Pseudomonas aeruginosa: An audacious pathogen with an adaptable arsenal of virulence factors. Int. J. Mol. Sci. 2021, 22, 3128. [Google Scholar] [CrossRef] [PubMed]
- Papa, R.; Parrilli, E.; Sannino, F.; Barbato, G.; Tutino, M.L.; Artini, M.; Selan, L. Anti-biofilm activity of the Antarctic marine bacterium Pseudoalteromonas haloplanktis TAC125. Res. Microbiol. 2013, 164, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Casillo, A.; Papa, R.; Ricciardelli, A.; Sannino, F.; Ziaco, M.; Tilotta, M.; Selan, L.; Marino, G.; Corsaro, M.M.; Tutino, M.L.; et al. Anti-biofilm activity of a long-chain fatty aldehyde from Antarctic Pseudoalteromonas haloplanktis TAC125 against Staphylococcus epidermidis biofilm. Front. Cell Infect. Microbiol. 2017, 7, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricciardelli, A.; Casillo, A.; Papa, R.; Monti, D.M.; Imbimbo, P.; Vrenna, G.; Artini, M.; Selan, L.; Corsaro, M.M.; Tutino, M.L.; et al. Pentadecanal inspired molecules as new anti-biofilm agents against Staphylococcus epidermidis. Biofouling 2018, 34, 1110–1120. [Google Scholar] [CrossRef]
- Price-Whelan, A.; Dietrich, L.E.; Newman, D.K. Rethinking ‘secondary’ metabolism: Physiological roles for phenazine antibiotics. Nat. Chem. Biol. 2006, 2, 71–78. [Google Scholar] [CrossRef]
- O’Malley, Y.Q.; Reszka, K.J.; Rasmussen, G.T.; Abdalla, M.Y.; Denning, G.M.; Britigan, B.E. The Pseudomonas secretory product pyocyanin inhibits catalase activity in human lung epithelial cells. Am. J. Physiol. Lung Cell Mo. Physiol. 2003, 285, L1077–L1086. [Google Scholar] [CrossRef] [Green Version]
- O’Malley, C.A. Infection control in cystic fibrosis: Cohorting, cross-contamination, and the respiratory therapist. Respir. Care 2009, 54, 641–657. [Google Scholar] [CrossRef]
- Ragno, R.; Papa, R.; Patsilinakos, A.; Vrenna, G.; Garzoli, S.; Tuccio, V.; Fiscarelli, E.; Selan, L.; Artini, M. Essential oils against bacterial isolates from cystic fibrosis patients by means of antimicrobial and unsupervised machine learning approaches. Sci. Rep. 2020, 10, 2653. [Google Scholar] [CrossRef]
- Papa, R.; Garzoli, S.; Vrenna, G.; Sabatino, M.; Sapienza, F.; Relucenti, M.; Donfrancesco, O.; Fiscarelli, E.V.; Artini, M.; Selan, L.; et al. Essential oils biofilm modulation activity, chemical and machine learning analysis. Application on Staphylococcus aureus isolates from cystic fibrosis patients. Int. J. Mol. Sci. 2020, 21, 9258. [Google Scholar] [CrossRef]
- Yang, R.; Guan, Y.; Zhou, J.; Sun, B.; Wang, Z.; Chen, H.; He, Z.; Jia, A. Phytochemicals from Camellia nitidissima chi flowers reduce the pyocyanin production and motility of Pseudomonas aeruginosa PAO1. Front. Microbiol. 2018, 8, 2640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bossù, M.; Selan, L.; Artini, M.; Relucenti, M.; Familiari, G.; Papa, R.; Vrenna, G.; Spigaglia, P.; Barbanti, F.; Salucci, A.; et al. Characterization of Scardovia wiggsiae biofilm by original scanning electron microscopy protocol. Microorganisms 2020, 8, 807. [Google Scholar] [CrossRef] [PubMed]
- Relucenti, M.; Miglietta, S.; Bove, G.; Donfrancesco, O.; Battaglione, E.; Familiari, P.; Barbaranelli, C.; Covelli, E.; Barbara, M.; Familiari, G. SEM BSE 3D image analysis of human incus bone affected by cholesteatoma ascribes to osteoclasts the bone erosion and VpSEM dEDX analysis reveals new bone formation. Scanning 2020, 2020, 9371516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Relucenti, M.; Familiari, G.; Donfrancesco, O.; Taurino, M.; Li, X.; Chen, R.; Artini, M.; Papa, R.; Selan, L. Microscopy methods for biofilm imaging: Focus on SEM and VP-SEM pros and cons. Biology 2021, 10, 51. [Google Scholar] [CrossRef]
- Restaino, O.F.; Finamore, R.; Diana, P.; Marseglia, M.; Vitiello, M.; Casillo, A.; Bedini, E.; Parrilli, M.; Corsaro, M.M.; Trifuoggi, M.; et al. A multi-analytical approach to better assess the keratan sulfate contamination in animal origin chondroitin sulfate. Anal. Chim. Acta 2017, 958, 59–70. [Google Scholar] [CrossRef]
- Carillo, S.; Pieretti, G.; Parrilli, E.; Tutino, M.L.; Gemma, S.; Molteni, M.; Lanzetta, R.; Parrilli, M.; Corsaro, M.M. Structural investigation and biological activity of the lipooligosaccharide from the psychrophilic bacterium Pseudoalteromonas haloplanktis TAB 23. Chem. Eur. J. 2011, 17, 7053–7060. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain | Pyocyanin (OD 520 nm) | Protease Activity (OD 400 nm) | Biofilm 24 h a (OD 590 nm) | Biofilm 48 h b (OD 590 nm) | Swarming Motility c | Swimming Motility c |
|---|---|---|---|---|---|---|
| PA14 | 0.178 ± 0.042 | 2.037 ± 0.020 | 3.561 ± 0.357 | 13.470 ± 1.403 | +++ | +++ |
| 23P | 0.148 ± 0.024 | 1.988 ± 0.057 | 3.175 ± 0.851 | 0.738 ± 0.373 | +++ | +++ |
| 27P | 0.102 ± 0.005 | 2.175 ± 0.036 | 1.429 ± 0.643 | 3.049 ± 0.796 | +++ | +++ |
| 31P | 0.063 ± 0.031 | 1.915 ± 0.001 | 1.741 ± 0.154 | 5.133 ± 0.946 | +++ | +++ |
| 32P | 0.121 ± 0.037 | 2.439 ± 0.078 | 1.117 ± 0.163 | 5.597 ± 1.390 | +++ | +++ |
| 40P | 0.160 ± 0.041 | 2.064 ± 0.058 | 0.970 ± 0.201 | 2.172 ± 0.194 | +++ | +++ |
| ID pt | ID | SAMPLE | Date | Phenotype | 1st Infection | Early Infection | Late Infection |
|---|---|---|---|---|---|---|---|
| 22 | 23P | ESP | 6/24/2005 | sm | X | ||
| 24 | 27P | AT | 1/31/2017 | sm | X | ||
| 9 | 31P | ESP | 1/11/2017 | M | X | ||
| 26 | 32P | AT | 12/5/2006 | sm | X | ||
| 30 | 40P | AT | 7/1/2013 | i | X |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papa, R.; Vrenna, G.; D’Angelo, C.; Casillo, A.; Relucenti, M.; Donfrancesco, O.; Corsaro, M.M.; Fiscarelli, E.V.; Tuccio Guarna Assanti, V.; Tutino, M.L.; et al. Anti-Virulence Activity of the Cell-Free Supernatant of the Antarctic Bacterium Psychrobacter sp. TAE2020 against Pseudomonas aeruginosa Clinical Isolates from Cystic Fibrosis Patients. Antibiotics 2021, 10, 944. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10080944
Papa R, Vrenna G, D’Angelo C, Casillo A, Relucenti M, Donfrancesco O, Corsaro MM, Fiscarelli EV, Tuccio Guarna Assanti V, Tutino ML, et al. Anti-Virulence Activity of the Cell-Free Supernatant of the Antarctic Bacterium Psychrobacter sp. TAE2020 against Pseudomonas aeruginosa Clinical Isolates from Cystic Fibrosis Patients. Antibiotics. 2021; 10(8):944. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10080944
Chicago/Turabian StylePapa, Rosanna, Gianluca Vrenna, Caterina D’Angelo, Angela Casillo, Michela Relucenti, Orlando Donfrancesco, Maria Michela Corsaro, Ersilia Vita Fiscarelli, Vanessa Tuccio Guarna Assanti, Maria Luisa Tutino, and et al. 2021. "Anti-Virulence Activity of the Cell-Free Supernatant of the Antarctic Bacterium Psychrobacter sp. TAE2020 against Pseudomonas aeruginosa Clinical Isolates from Cystic Fibrosis Patients" Antibiotics 10, no. 8: 944. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10080944