
Effect of Lactobacillus plantarum Biofilms on the Adhesion of Escherichia coli to Urinary Tract Devices


Abstract
:1. Introduction
2. Results
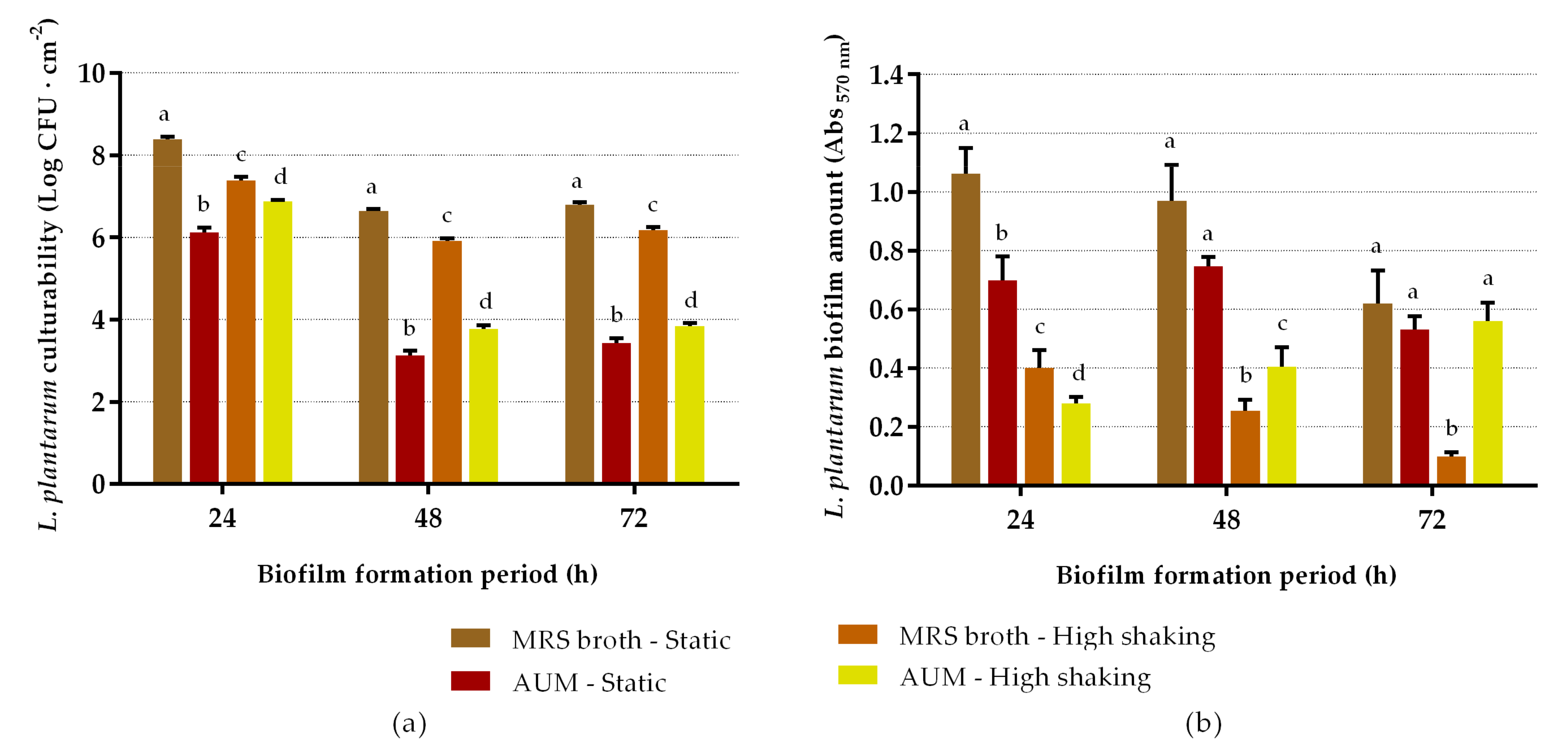
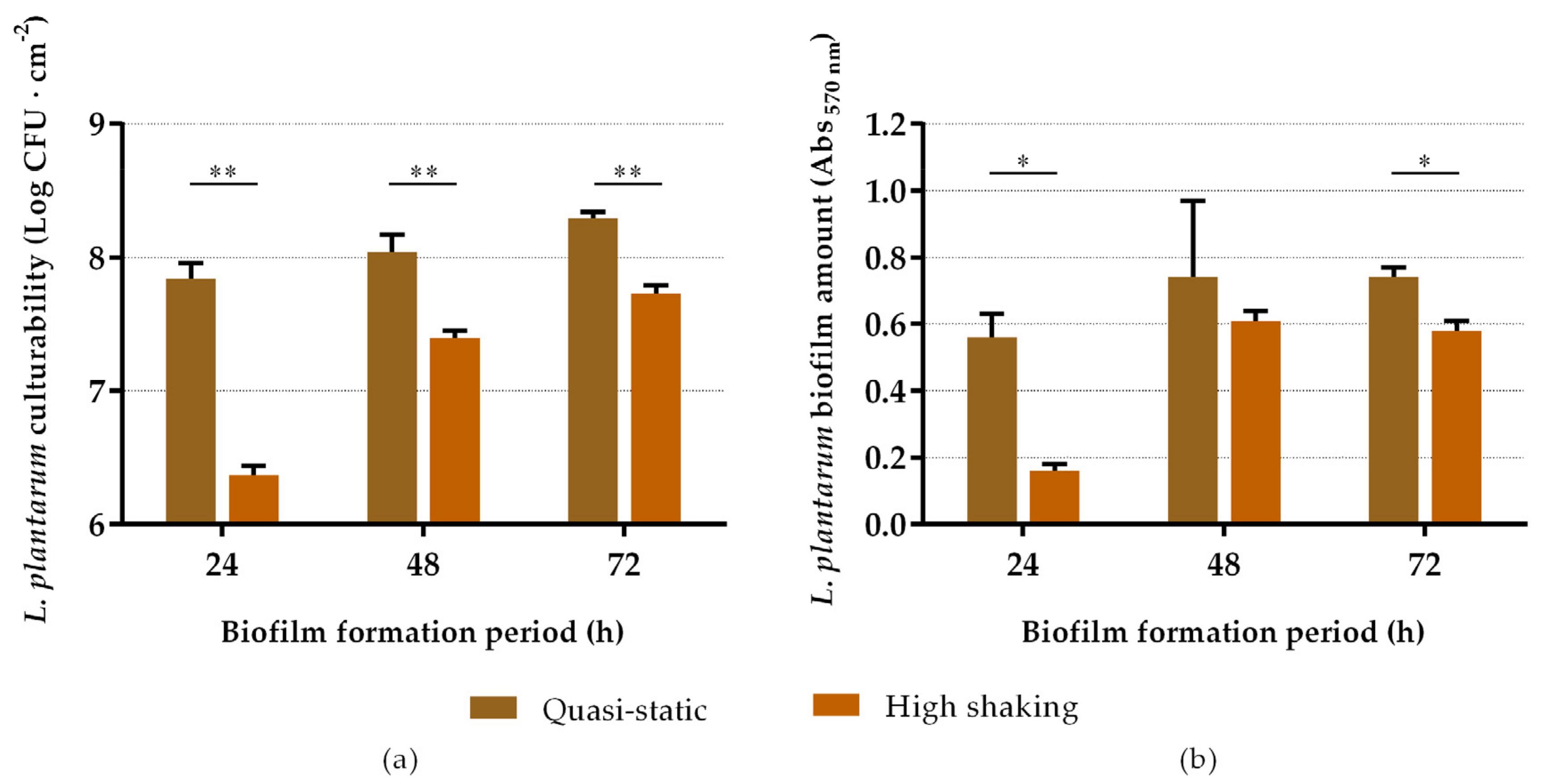
2.1. Biofilm-Forming Capacity of L. plantarum
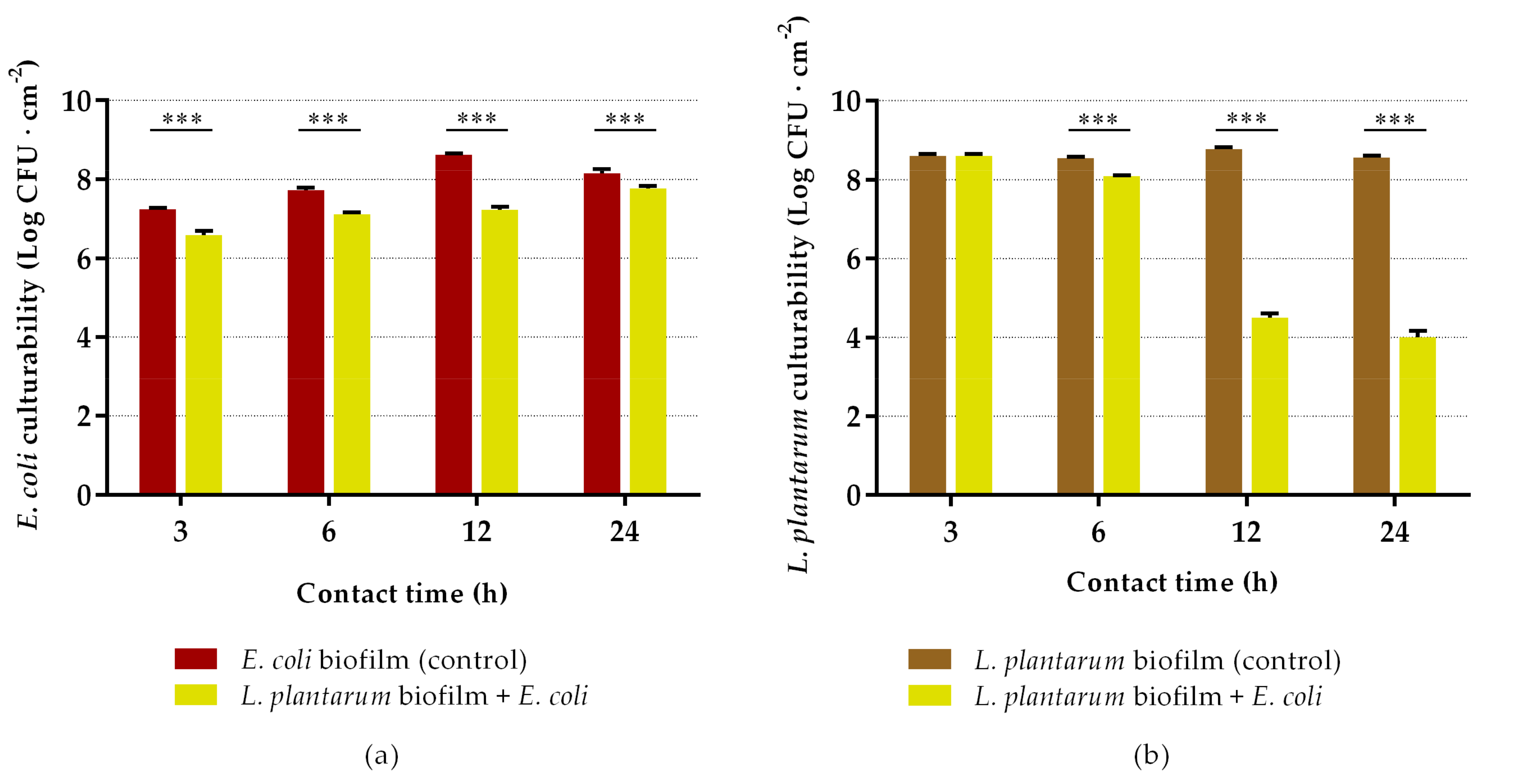
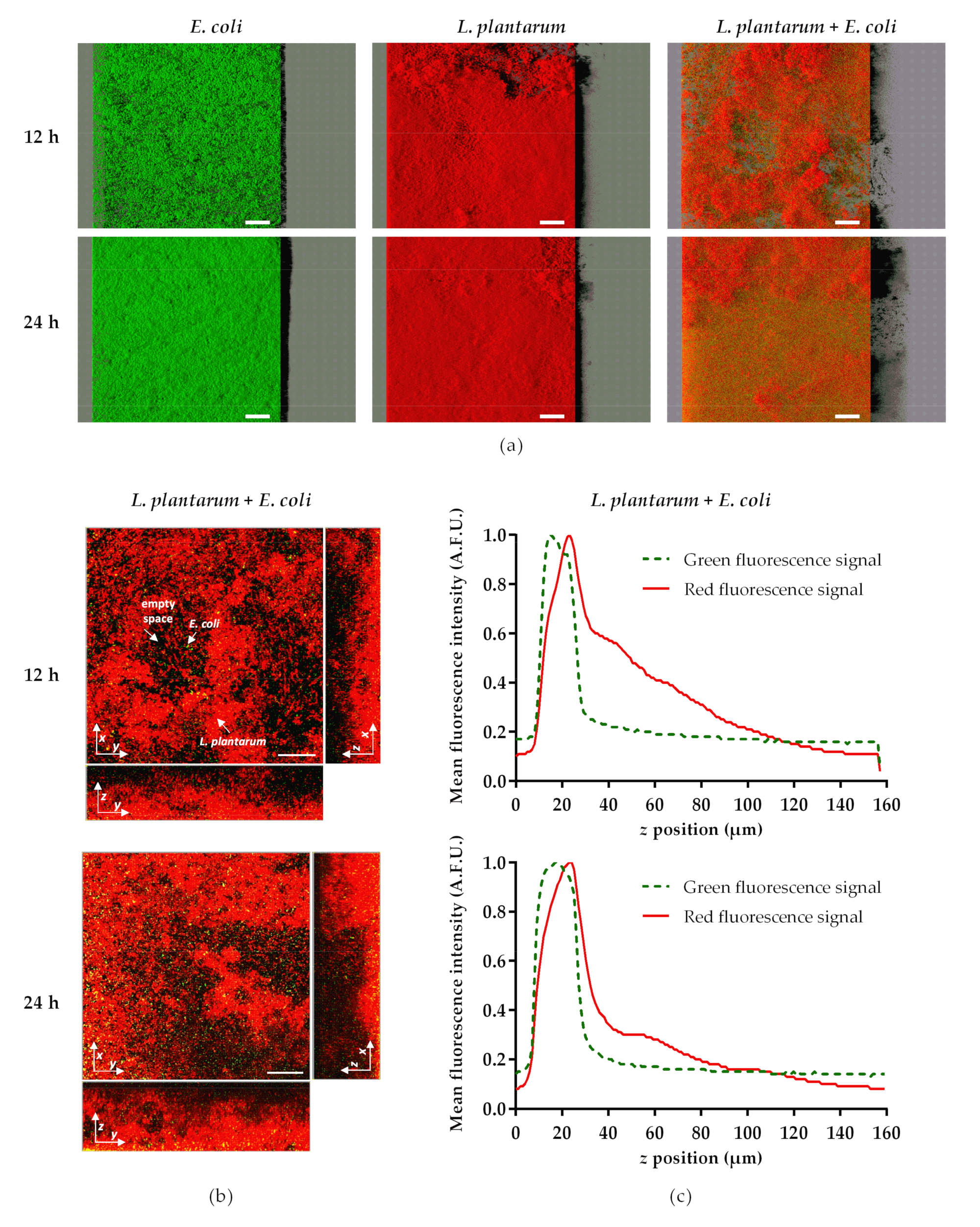
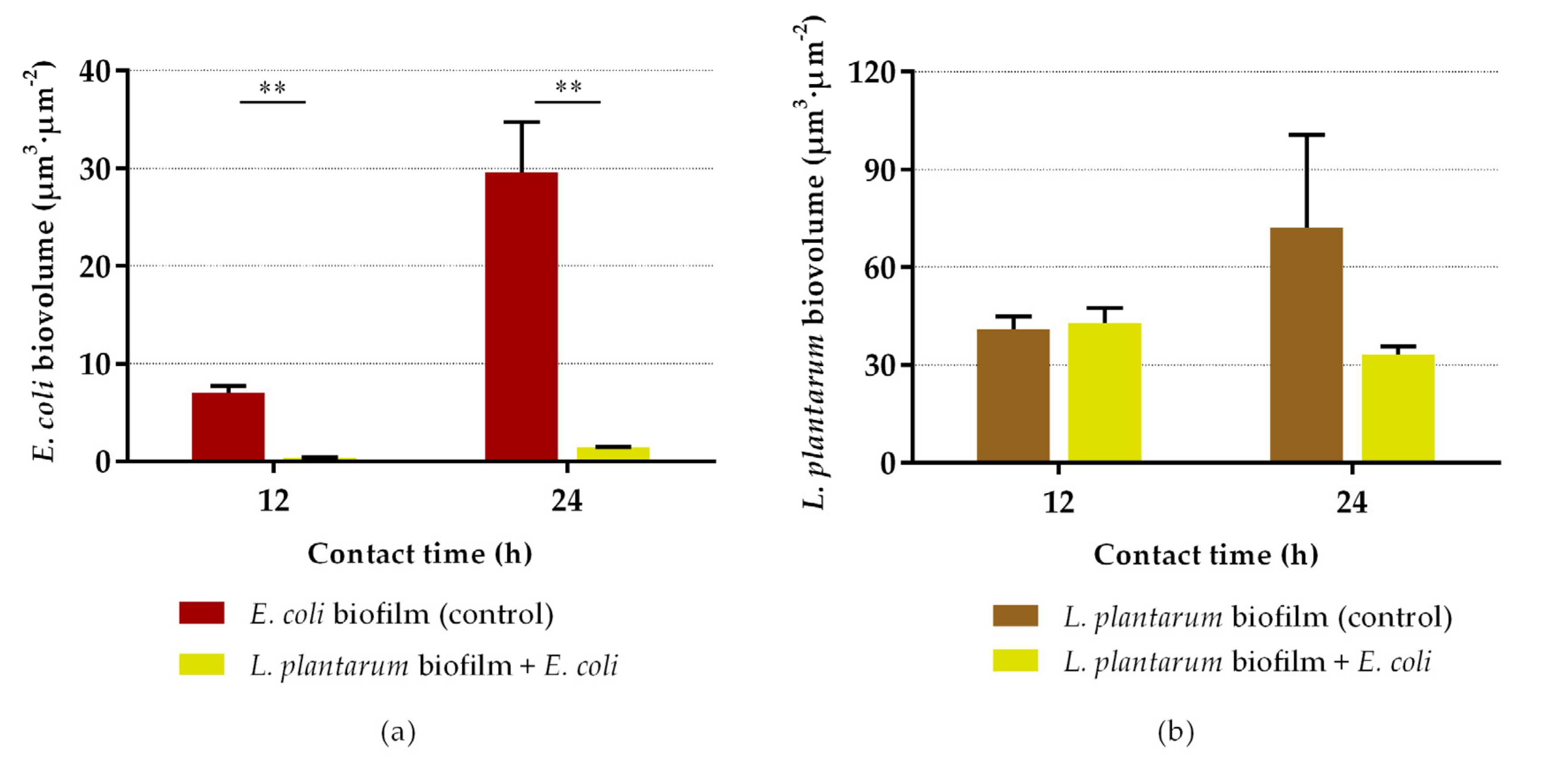
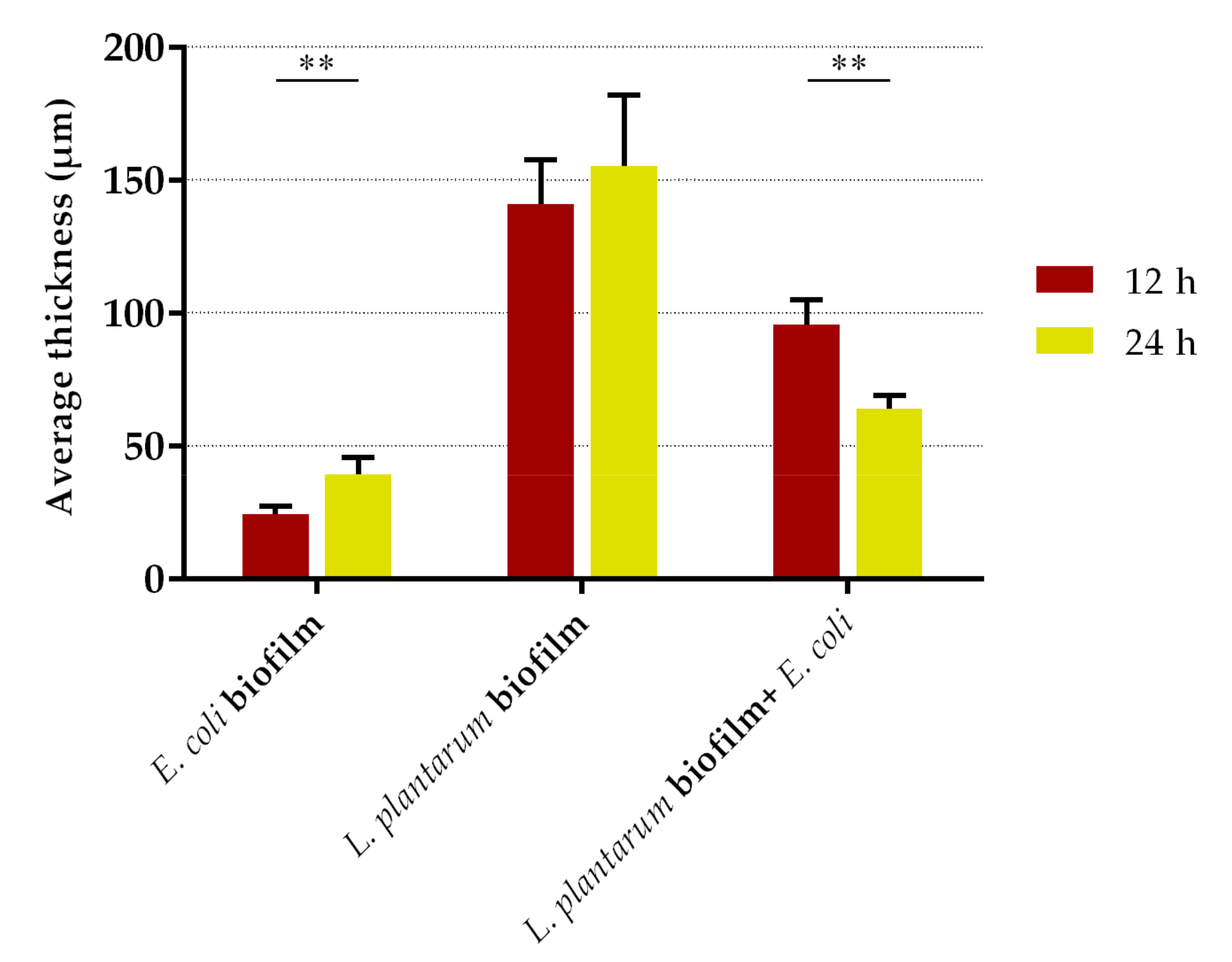
2.2. Antibiofilm Assays
2.3. Thermodynamic Surface Properties
3. Discussion
4. Materials and Methods
4.1. Preparation of Silicone Surfaces
4.2. Bacterial Strains and Culture Conditions
4.3. Biofilm-Forming Capacity of L. plantarum
4.4. Antibiofilm Assays
4.5. Colony-Forming Units (CFU) Enumeration
4.6. Crystal Violet (CV) Staining
4.7. Confocal Laser Scanning Microscopy (CLSM) Analysis
4.8. Contact Angle Measurements, Hydrophobicity, and Free Energy of Adhesion between Bacteria and Substrates
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Corte, L.; Pierantoni, D.C.; Tascini, C.; Roscini, L.; Cardinali, G. Biofilm specific activity: A measure to quantify microbial biofilm. Microorganisms 2019, 7, 73. [Google Scholar] [CrossRef] [Green Version]
- Rabin, N.; Zheng, Y.; Opoku-Temeng, C.; Du, Y.; Bonsu, E.; Sintim, H.O. Biofilm formation mechanisms and targets for developing antibiofilm agents. Future Med. Chem. 2015, 7, 493–512. [Google Scholar] [CrossRef] [PubMed]
- Khatoon, Z.; McTiernan, C.D.; Suuronen, E.J.; Mah, T.F.; Alarcon, E.I. Bacterial biofilm formation on implantable devices and approaches to its treatment and prevention. Heliyon 2018, 4, e01067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azevedo, A.S.; Almeida, C.; Melo, L.F.; Azevedo, N.F. Impact of polymicrobial biofilms in catheter-associated urinary tract infections. Crit. Rev. Microbiol. 2017, 43, 423–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, H.; Zhang, J.; Qu, J.; Liu, J.; Yin, P.; Zhang, G.; Shang, D. Lactobacillus rhamnosus GG microcapsules inhibit Escherichia coli biofilm formation in coculture. Biotechnol. Lett. 2019, 41, 1007–1014. [Google Scholar] [CrossRef]
- Medina, M.; Castillo-Pino, E. An introduction to the epidemiology and burden of urinary tract infections. Ther. Adv. Urol. 2019, 11, 1756287219832172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OECD/EU Health at a Glance: Europe 2018 State of Health in the EU Cycle. Available online: https://0-doi-org.brum.beds.ac.uk/10.1787/health_glance_eur-2018-en (accessed on 3 August 2021).
- Maharjan, G.; Khadka, P.; Siddhi Shilpakar, G.; Chapagain, G.; Dhungana, G.R. Catheter-Associated Urinary Tract Infection and Obstinate Biofilm Producers. Can. J. Infect. Dis. Med. Microbiol. 2018, 2018, 7624857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramstedt, M.; Ribeiro, I.A.C.; Bujdakova, H.; Mergulhão, F.J.M.; Jordao, L.; Thomsen, P.; Alm, M.; Burmølle, M.; Vladkova, T.; Can, F.; et al. Evaluating efficacy of antimicrobial and antifouling materials for urinary tract medical devices: Challenges and recommendations. Macromol. Biosci. 2019, 19, e1800384. [Google Scholar] [CrossRef] [Green Version]
- Feneley, R.C.L.; Hopley, I.B.; Wells, P.N.T. Urinary catheters: History, current status, adverse events and research agenda. J. Med. Eng. Technol. 2015, 39, 459–470. [Google Scholar] [CrossRef] [Green Version]
- Singha, P.; Locklin, J.; Handa, H. A review of the recent advances in antimicrobial coatings for urinary catheters. Acta Biomater. 2017, 50, 20–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centers for Disease Control and Prevention. Catheter-associated Urinary Tract Infections (CAUTI). Available online: https://www.cdc.gov/hai/ca_uti/uti.html (accessed on 4 August 2021).
- Lawrence, E.L.; Turner, I.G. Materials for urinary catheters: A review of their history and development in the UK. Med. Eng. Phys. 2005, 27, 443–453. [Google Scholar] [CrossRef]
- Donlan, R.M. Biofilms and device-associated infections. Emerg. Infect. Dis. 2001, 7, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Seif Eldein, S.S.; El-Temawy, A.-E.-K.A.; Ahmed, E.H. Biofilm Formation by E. coli Causing Catheter Associated Urinary Tract Infection (CAUTI) in Assiut University Hospital. Egypt. J. Med. Microbiol. 2013, 22, 101–110. [Google Scholar] [CrossRef]
- Niveditha, S.; Pramodhini, S.; Umadevi, S.; Kumar, S.; Stephen, S. The isolation and the biofilm formation of uropathogens in the patients with catheter associated urinary tract infections (UTIs). J. Clin. Diagn. Res. 2012, 6, 1478–1482. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.G.; Palermo, J.J.; Schilling, J.D.; Roth, R.; Heuser, J.; Hultgren, S.J. Intracellular bacterial biofilm-like pods in urinary tract infections. Science 2003, 301, 105–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Yu, Q.; Sun, H. Novel strategies for the prevention and treatment of biofilm related infections. Int. J. Mol. Sci. 2013, 14, 18488–18501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Z.; Wang, Z.; Li, S.; Yuan, X. Antimicrobial strategies for urinary catheters. J. Biomed. Mater. Res. Part A 2019, 107, 445–467. [Google Scholar] [CrossRef] [PubMed]
- Walencka, E.; Różalska, S.; Sadowska, B.; Rózalska, B. The influence of Lactobacillus acidophilus-derived surfactants on staphylococcal adhesion and biofilm formation. Folia Microbiol. 2008, 53, 61–66. [Google Scholar] [CrossRef]
- Barzegari, A.; Kheyrolahzadeh, K.; Mahdi, S.; Khatibi, H.; Sharifi, S.; Memar, M.Y.; Vahed, S.Z. The battle of probiotics and their derivatives against biofilms. Infect. Drug Resist. 2020, 13, 659–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, F.M.; Teixeira-Santos, R.; Mergulhão, F.J.M.; Gomes, L.C. The use of probiotics to fight biofilms in medical devices: A systematic review and meta-analysis. Microorganisms 2021, 9, 27. [Google Scholar] [CrossRef]
- Carvalho, F.M.; Teixeira-Santos, R.; Mergulhão, F.J.M.; Gomes, L.C. Targeting biofilms in medical devices using probiotic cells: A systematic review. AIMS Mater. Sci. 2021, 8, 501–523. [Google Scholar] [CrossRef]
- FAO. Probiotics in Food: Health and Nutritional Properties and Guidelines for Evaluation. Food & Agriculture Org: Rome, Italy, 2006; Available online: http://www.fao.org/3/a0512e/a0512e.pdf (accessed on 9 August 2021).
- Reis, J.A.; Paula, A.T.; Casarotti, S.N.; Penna, A.L.B. Lactic Acid Bacteria Antimicrobial Compounds: Characteristics and Applications. Food Eng. Rev. 2012, 4, 124–140. [Google Scholar] [CrossRef]
- Fioramonti, J.; Theodorou, V.; Bueno, L. Probiotics: What are they? What are their effects on gut physiology? Best Pract. Res. Clin. Gastroenterol. 2003, 17, 711–724. [Google Scholar] [CrossRef]
- Gogineni, V.K.; Morrow, L.E. Probiotics: Mechanisms of action and clinical applications. J. Probiotics Health 2013, 1, 101. [Google Scholar] [CrossRef] [Green Version]
- Gomaa, E.Z. Antimicrobial and anti-adhesive properties of biosurfactant produced by lactobacilli isolates, biofilm formation and aggregation ability. J. Gen. Appl. Microbiol. 2013, 59, 425–436. [Google Scholar] [CrossRef] [Green Version]
- Aoudia, N.; Rieu, A.; Briandet, R.; Deschamps, J.; Chluba, J.; Jego, G.; Garrido, C.; Guzzo, J. Biofilms of Lactobacillus plantarum and Lactobacillus fermentum: Effect on stress responses, antagonistic effects on pathogen growth and immunomodulatory properties. Food Microbiol. 2016, 53, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Khalighi, A.; Behdani, R.; Kouhestani, S. Probiotics: A comprehensive review of their classification, mode of action and role in human nutrition. In Probiotics and Prebiotics in Human Nutrition and Health; InTech: London, UK, 2016. [Google Scholar]
- Prabhurajeshwar, C.; Chandrakanth, R.K. Probiotic potential of lactobacilli with antagonistic activity against pathogenic strains: An in vitro validation for the production of inhibitory substances. Biomed. J. 2017, 40, 270–283. [Google Scholar] [CrossRef]
- Muñoz, M.; Mosquera, A.; Alméciga-Díaz, C.J.; Melendez, A.P.; Sánchez, O.F. Fructooligosaccharides metabolism and effect on bacteriocin production in Lactobacillus strains isolated from ensiled corn and molasses. Anaerobe 2012, 18, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Kechagia, M.; Basoulis, D.; Konstantopoulou, S.; Dimitriadi, D.; Gyftopoulou, K.; Skarmoutsou, N.; Fakiri, E.M. Health benefits of probiotics: A review. ISRN Nutr. 2013, 2013, 481651. [Google Scholar] [CrossRef] [Green Version]
- Holzapfel, W.H.; Haberer, P.; Geisen, R.; Björkroth, J.; Schillinger, U. Taxonomy and important features of probiotic microorganisms in food and nutrition. Am. J. Clin. Nutr. 2001, 73, 365–373. [Google Scholar] [CrossRef] [Green Version]
- De Melo Pereira, G.V.; de Oliveira Coelho, B.; Magalhães Júnior, A.I.; Thomaz-Soccol, V.; Soccol, C.R. How to select a probiotic? A review and update of methods and criteria. Biotechnol. Adv. 2018, 36, 2060–2076. [Google Scholar] [CrossRef]
- Ng, S.C.; Hart, A.L.; Kamm, M.A.; Stagg, A.J.; Knight, S.C. Mechanisms of action of probiotics: Recent advances. Inflamm. Bowel Dis. 2009, 15, 300–310. [Google Scholar] [CrossRef]
- Bermudez-Brito, M.; Plaza-Díaz, J.; Muñoz-Quezada, S.; Gómez-Llorente, C.; Gil, A. Probiotic mechanisms of action. Ann. Nutr. Metab. 2012, 61, 160–174. [Google Scholar] [CrossRef]
- Salas-Jara, M.; Ilabaca, A.; Vega, M.; García, A. Biofilm Forming Lactobacillus: New Challenges for the Development of Probiotics. Microorganisms 2016, 4, 35. [Google Scholar] [CrossRef] [PubMed]
- Nemcová, R. Criteria for selection of lactobacilli for probiotic use. Vet. Med. 1997, 42, 19–27. [Google Scholar]
- Chen, Q.; Zhu, Z.; Wang, J.; Lopez, A.I.; Li, S.; Kumar, A.; Yu, F.; Chen, H.; Cai, C.; Zhang, L. Probiotic E. coli Nissle 1917 biofilms on silicone substrates for bacterial interference against pathogen colonization. Acta Biomater. 2017, 50, 353–360. [Google Scholar] [CrossRef] [Green Version]
- Cadieux, P.; Watterson, J.D.; Denstedt, J.; Harbottle, R.R.; Puskas, J.; Howard, J.; Gan, B.S.; Reid, G. Potential application of polyisobutylene-polystyrene and a Lactobacillus protein to reduce the risk of device-associated urinary tract infections. Colloids Surf. B Biointerfaces 2003, 28, 95–105. [Google Scholar] [CrossRef]
- Reid, G.; Tieszer, C. Use of lactobacilli to reduce the adhesion of Staphylococcus aureus to catheters. Int. Biodeterior. Biodegrad. 1994, 34, 73–83. [Google Scholar] [CrossRef]
- Ray Mohapatra, A.; Jeevaratnam, K. Inhibiting bacterial colonization on catheters: Antibacterial and antibiofilm activities of bacteriocins from Lactobacillus plantarum SJ33. J. Glob. Antimicrob. Resist. 2019, 19, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Ifeoma, M.E.; Jennifer, U.K. Inhibition of biofilms on urinary catheters using immobilized Lactobacillus cells. Afr. J. Microbiol. Res. 2016, 10, 920–929. [Google Scholar] [CrossRef] [Green Version]
- Al-Mathkhury, H.J.F.; Ali, A.S.; Ghafil, J.A. Antagonistic effect of bacteriocin against urinary catheter associated Pseudomonas aeruginosa biofilm. N. Am. J. Med. Sci. 2011, 3, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Saharan, B.S. Functional characterization of biomedical potential of biosurfactant produced by Lactobacillus helveticus. Biotechnol. Rep. 2016, 11, 27–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velraeds, M.M.C.; van de Belt-Gritter, B.; Busscher, H.J.; Reid, G.; van Der Mei, H.C. Inhibition of uropathogenic biofilm growth on silicone rubber in human urine by lactobacilli—A teleologic approach. World J. Urol. 2000, 18, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Velraeds, M.M.C.; van De Belt-Gritter, B.; van Der Mei, H.C.; Reid, G.; Busscher, H.J. Interference in initial adhesion of uropathogenic bacteria and yeasts to silicone rubber by a Lactobacillus acidophilus biosurfactant. J. Med. Microbiol. 1998, 47, 1081–1085. [Google Scholar] [CrossRef] [PubMed]
- Alves, P.; Nir, S.; Reches, M.; Mergulhão, F. The effects of fluid composition and shear conditions on bacterial adhesion to an antifouling peptide-coated surface. MRS Commun. 2018, 8, 938–946. [Google Scholar] [CrossRef]
- Moreira, J.M.R.; Ponmozhi, J.; Campos, J.B.L.M.; Miranda, J.M.; Mergulhão, F.J. Micro- and macro-flow systems to study Escherichia coli adhesion to biomedical materials. Chem. Eng. Sci. 2015, 126, 440–445. [Google Scholar] [CrossRef] [Green Version]
- Gomes, M.; Gomes, L.C.; Teixeira-Santos, R.; Pereira, M.F.R.; Soares, O.S.G.P.; Mergulhão, F.J. Optimizing CNT Loading in Antimicrobial Composites for Urinary Tract Application. Appl. Sci. 2021, 11, 4038. [Google Scholar] [CrossRef]
- Kaur, S.; Sharma, P.; Kalia, N.; Singh, J.; Kaur, S. Anti-biofilm properties of the fecal probiotic lactobacilli against Vibrio spp. Front. Cell. Infect. Microbiol. 2018, 8, 120. [Google Scholar] [CrossRef] [Green Version]
- Maldonado, N.C.; Silva De Ruiz, C.; Cecilia, M.; Nader-Macias, M.E. A simple technique to detect Klebsiella biofilm-forming-strains. Inhibitory potential of Lactobacillus fermentum CRL 1058 whole cells and products. In Communicating Current Research and Educational Topics and Trends in Applied Microbiology; Vilas, A.M., Ed.; FORMATEX: Badajoz, Spain, 2007; pp. 52–59. Available online: https://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.597.5733&rep=rep1&type=pdf (accessed on 9 August 2021).
- Leroy, F.; De Vuyst, L. Growth of the Bacteriocin-Producing Lactobacillus sakei Strain CTC 494 in MRS Broth is Strongly Reduced Due to Nutrient Exhaustion: A Nutrient Depletion Model for the Growth of Lactic Acid Bacteria. Appl. Environ. Microbiol. 2001, 67, 4407–4413. [Google Scholar] [CrossRef] [Green Version]
- Sjollema, J.; Busscher, H.J.; Weerkamp, A.H. Deposition of oral streptococci and polystyrene latices onto glass in a parallel plate flow cell. Biofouling 1988, 1, 101–112. [Google Scholar] [CrossRef]
- Millsap, K.W.; Reid, G.; van Der Mei, H.C.; Busscher, H.J. Adhesion of Lactobacillus species in urine and phosphate buffer to silicone rubber and glass under flow. Biomaterials 1997, 18, 87–91. [Google Scholar] [CrossRef]
- Bujňáková, D.; Kmet’, V. Functional properties of Lactobacillus strains isolated from dairy products. Folia Microbiol. 2012, 57, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Kubota, H.; Senda, S.; Nomura, N.; Tokuda, H.; Uchiyama, H. Biofilm Formation by Lactic Acid Bacteria and Resistance to Environmental Stress. J. Biosci. Bioeng. 2008, 106, 381–386. [Google Scholar] [CrossRef]
- Jalilsood, T.; Baradaran, A.; Song, A.A.L.; Foo, H.L.; Mustafa, S.; Saad, W.Z.; Yusoff, K.; Rahim, R.A. Inhibition of pathogenic and spoilage bacteria by a novel biofilm-forming Lactobacillus isolate: A potential host for the expression of heterologous proteins. Microb. Cell Fact. 2015, 14, 96. [Google Scholar] [CrossRef] [Green Version]
- Klimko, A.I.; Cherdyntseva, T.A.; Brioukhanov, A.L.; Netrusov, A.I. In vitro Evaluation of Probiotic Potential of Selected Lactic Acid Bacteria Strains. Probiotics Antimicrob. Proteins 2020, 12, 1139–1148. [Google Scholar] [CrossRef]
- Simões, M.; Simões, L.C.; Vieira, M.J. A review of current and emergent biofilm control strategies. LWT Food Sci. Technol. 2010, 43, 573–583. [Google Scholar] [CrossRef] [Green Version]
- Fernández Ramírez, M.D.; Smid, E.J.; Abee, T.; Nierop Groot, M.N. Characterisation of biofilms formed by Lactobacillus plantarum WCFS1 and food spoilage isolates. Int. J. Food Microbiol. 2015, 207, 23–29. [Google Scholar] [CrossRef]
- Krasowska, A.; Sigler, K. How microorganisms use hydrophobicity and what does this mean for human needs? Front. Cell. Infect. Microbiol. 2014, 4, 112. [Google Scholar] [CrossRef] [Green Version]
- Honey Chandran, C.; Keerthi, T.R. Probiotic potency of Lactobacillus plantarum KX519413 and KX519414 isolated from honey bee gut. FEMS Microbiol. Lett. 2018, 365, fnx285. [Google Scholar] [CrossRef]
- Malik, S.; Petrova, M.I.; Claes, I.J.J.; Verhoeven, T.L.A.; Busschaert, P.; Vaneechoutte, M.; Lievens, B.; Lambrichts, I.; Siezen, R.J.; Balzarini, J.; et al. The highly autoaggregative and adhesive phenotype of the vaginal Lactobacillus plantarum strain cmpg5300 is sortase dependent. Appl. Environ. Microbiol. 2013, 79, 4576–4585. [Google Scholar] [CrossRef] [Green Version]
- Twetman, L.; Larsen, U.; Fiehn, N.E.; Steckésn-Blicks, C.; Twetman, S. Coaggregation between probiotic bacteria and caries-associated strains: An in vitro study. Acta Odontol. Scand. 2009, 67, 284–288. [Google Scholar] [CrossRef] [PubMed]
- Faten, K.; Hamida, K.; Soumya, E.A.; Saad, I.S.K.; Hasna, M.; Hassan, L.; Moktar, H. Lactobacillus plantarum: Effect of a protective biofilm on the surface of olives during storage. Braz. J. Microbiol. 2016, 47, 202–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vertes, A.; Hitchins, V.; Phillips, K.S. Analytical challenges of microbial biofilms on medical devices. Anal. Chem. 2012, 84, 3858–3866. [Google Scholar] [CrossRef] [PubMed]
- Percival, S.L.; Suleman, L.; Vuotto, C.; Donelli, G. Healthcare-associated infections, medical devices and biofilms: Risk, tolerance and control. J. Med. Microbiol. 2015, 64, 323–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, J.; Halami, P.M. Presence of extracellular DNA & protein in biofilm formation by gentamicin-resistant Lactobacillus plantarum. Indian J. Med. Res. 2019, 149, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Muscariello, L.; Marino, C.; Capri, U.; Vastano, V.; Marasco, R.; Sacco, M. CcpA and three newly identified proteins are involved in biofilm development in Lactobacillus plantarum. J. Basic Microbiol. 2013, 53, 62–71. [Google Scholar] [CrossRef]
- Schiøtz, H.; Tanbo, T. Postoperative voiding, bacteriuria and urinary tract infection with Foley catheterization after gynecological surgery. Acta Obstet. Gynecol. Scand. 2006, 85, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Dong, L.; Gu, L. The timing of urinary catheter removal after gynecologic surgery: A meta-analysis of randomized controlled trials. Medicine (United States) 2020, 99, e18710. [Google Scholar] [CrossRef]
- Alessandri, F.; Mistrangelo, E.; Lijoi, D.; Ferrero, S.; Ragni, N. A prospective, randomized trial comparing immediate versus delayed catheter removal following hysterectomy. Acta Obstet. Gynecol. Scand. 2006, 85, 716–720. [Google Scholar] [CrossRef] [PubMed]
- Glavind, K.; Mørup, L.; Madsen, H.; Glavind, J. A prospective, randomised, controlled trial comparing 3 hour and 24 hour postoperative removal of bladder catheter and vaginal pack following vaginal prolapse surgery. Acta Obstet. Gynecol. Scand. 2007, 86, 1122–1125. [Google Scholar] [CrossRef]
- Hendren, S. Urinary catheter management. Clin. Colon Rectal Surg. 2013, 26, 178–181. [Google Scholar] [CrossRef] [Green Version]
- Basbug, A.; Yuksel, A.; Ellibeş Kaya, A. Early versus delayed removal of indwelling catheters in patients after elective cesarean section: A prospective randomized trial. J. Matern. Neonatal Med. 2020, 33, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, R.; Griffiths, R.; Murie, P. Comparison of late night and early morning removal of short-term urethral catheters. JBI Libr. Syst. Rev. 2003, 1, 1–26. [Google Scholar] [CrossRef]
- Ahmed, M.R.; Sayed Ahmed, W.A.; Atwa, K.A.; Metwally, L. Timing of urinary catheter removal after uncomplicated total abdominal hysterectomy: A prospective randomized trial. Eur. J. Obstet. Gynecol. Reprod. Biol. 2014, 176, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Kin, C.; Rhoads, K.F.; Jalali, M.; Shelton, A.A.; Welton, M.L. Predictors of postoperative urinary retention after colorectal surgery. Dis. Colon Rectum 2013, 56, 738–746. [Google Scholar] [CrossRef]
- Benoist, S.; Panis, Y.; Denet, C.; Mauvais, F.; Mariani, P.; Valleur, P. Optimal duration of urinary drainage after rectal resection: A randomized controlled trial. Surgery 1999, 125, 135–141. [Google Scholar] [CrossRef]
- Dodds, L.; Lawson, P.S.; Crosthwaite, A.H.; Wells, G.R. Early catheter removal: A prospective study of 100 consecutive patients undergoing transurethral resection of the prostate. Br. J. Urol. 1995, 75, 755–757. [Google Scholar] [CrossRef]
- Brackmann, M.; Carballo, E.; Uppal, S.; Torski, J.; Reynolds, R.K.; McLean, K. Implementation of a standardized voiding management protocol to reduce unnecessary re-catheterization—A quality improvement project. Gynecol. Oncol. 2020, 157, 487–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gould, C.V.; Umscheid, C.A.; Agarwal, R.K.; Kuntz, G.; Pegues, D.A. Guideline for Prevention of Catheter-Associated Urinary Tract Infections 2009. Infect Control Hosp Epidemiol. 2010, 4, 319–326. [Google Scholar] [CrossRef] [Green Version]
- Tan, Y.; Leonhard, M.; Moser, D.; Ma, S.; Schneider-Stickler, B. Inhibitory effect of probiotic lactobacilli supernatants on single and mixed non-albicans Candida species biofilm. Arch. Oral Biol. 2018, 85, 40–45. [Google Scholar] [CrossRef]
- Alexandre, Y.; Le Berre, R.; Barbier, G.; Le Blay, G. Screening of Lactobacillus spp. for the prevention of Pseudomonas aeruginosa pulmonary infections. BMC Microbiol. 2014, 14, 107. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Chen, X.; Tu, Y.; Wang, S.; Chen, H. Effect of probiotic lactobacilli on the growth of Streptococcus mutans and multispecies biofilms isolated from children with active caries. Med. Sci. Monit. 2017, 23, 4175–4181. [Google Scholar] [CrossRef] [Green Version]
- Teanpaisan, R.; Piwat, S.; Dahlén, G. Inhibitory effect of oral Lactobacillus against oral pathogens. Lett. Appl. Microbiol. 2011, 53, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-Barragán, A.; Caballero-Guerrero, B.; Lucena-Padrós, H.; Ruiz-Barba, J.L. Induction of bacteriocin production by coculture is widespread among plantaricin-producing Lactobacillus plantarum strains with different regulatory operons. Food Microbiol. 2013, 33, 40–47. [Google Scholar] [CrossRef]
- Hasslöf, P.; Hedberg, M.; Twetman, S.; Stecksén-Blicks, C. Growth inhibition of oral mutans streptococci and candida by commercial probiotic lactobacilli—An In Vitro study. BMC Oral Health 2010, 10, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Vahedi Shahandashti, R.; Kasra Kermanshahi, R.; Ghadam, P. The inhibitory effect of bacteriocin produced by Lactobacillus acidophilus ATCC 4356 and Lactobacillus plantarum ATCC 8014 on planktonic cells and biofilms of Serratia marcescens. Turk. J. Med. Sci. 2016, 46, 1188–1196. [Google Scholar] [CrossRef] [PubMed]
- Zalán, Z.; Németh, E.; Baráth, Á.; Halász, A. Influence of growth medium on hydrogen peroxide and bacteriocin production of Lactobacillus strains. Food Technol. Biotechnol. 2005, 43, 219–225. [Google Scholar]
- Sharma, V.; Harjai, K.; Shukla, G. Effect of bacteriocin and exopolysaccharides isolated from probiotic on P. aeruginosa PAO1 biofilm. Folia Microbiol. 2018, 63, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Besser, M.; Terberger, J.; Weber, L.; Ghebremedhin, B.; Naumova, E.A.; Arnold, W.H.; Stuermer, E.K. Impact of probiotics on pathogen survival in an innovative human plasma biofilm model (hpBIOM). J. Transl. Med. 2019, 17, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasfi, R.; Abd El-Rahman, O.A.; Zafer, M.M.; Ashour, H.M. Probiotic Lactobacillus sp. inhibit growth, biofilm formation and gene expression of caries-inducing Streptococcus mutans. J. Cell. Mol. Med. 2018, 22, 1972–1983. [Google Scholar] [CrossRef] [Green Version]
- Kang, D.K.; Oh, H.K.; Ham, J.S.; Kim, J.G.; Yoon, C.H.; Ahn, Y.T.; Kim, H.U. Identification and characterization of hydrogen peroxide-generating Lactobacillus fermentum CS12-1. Asian Australas. J. Anim. Sci. 2005, 18, 90–95. [Google Scholar] [CrossRef]
- Kumar, S.R.; Imlay, J.A. How Escherichia coli tolerates profuse hydrogen peroxide formation by a catabolic pathway. J. Bacteriol. 2013, 195, 4569–4579. [Google Scholar] [CrossRef] [Green Version]
- Imlay, J.A.; Linn, S. Mutagenesis and stress responses induced in Escherichia coli by hydrogen peroxide. J. Bacteriol. 1987, 169, 2967–2976. [Google Scholar] [CrossRef] [Green Version]
- Hegde, A.; Bhat, G.; Mallya, S. Effect of exposure to hydrogen peroxide on the virulence of Escherichia coli. Indian J. Med. Microbiol. 2008, 26, 25–28. [Google Scholar] [CrossRef]
- Matsubara, V.H.; Wang, Y.; Bandara, H.M.H.N.; Mayer, M.P.A.; Samaranayake, L.P. Probiotic lactobacilli inhibit early stages of Candida albicans biofilm development by reducing their growth, cell adhesion, and filamentation. Appl. Microbiol. Biotechnol. 2016, 100, 6415–6426. [Google Scholar] [CrossRef] [Green Version]
- Merghni, A.; Dallel, I.; Noumi, E.; Kadmi, Y.; Hentati, H.; Tobji, S.; Ben Amor, A.; Mastouri, M. Antioxidant and antiproliferative potential of biosurfactants isolated from Lactobacillus casei and their anti-biofilm effect in oral Staphylococcus aureus strains. Microb. Pathog. 2017, 104, 84–89. [Google Scholar] [CrossRef]
- Satpute, S.K.; Mone, N.S.; Das, P.; Banat, I.M.; Banpurkar, A.G. Inhibition of pathogenic bacterial biofilms on PDMS based implants by L. acidophilus derived biosurfactant. BMC Microbiol. 2019, 19, 1–15. [Google Scholar] [CrossRef]
- Sambanthamoorthy, K.; Feng, X.; Patel, R.; Patel, S.; Paranavitana, C. Antimicrobial and antibiofilm potential of biosurfactants isolated from lactobacilli against multi-drug-resistant pathogens. BMC Microbiol. 2014, 14, 197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceresa, C.; Tessarolo, F.; Caola, I.; Nollo, G.; Cavallo, M.; Rinaldi, M.; Fracchia, L. Inhibition of Candida albicans adhesion on medical-grade silicone by a Lactobacillus-derived biosurfactant. J. Appl. Microbiol. 2015, 118, 1116–1125. [Google Scholar] [CrossRef] [PubMed]
- Satpute, S.K.; Kulkarni, G.R.; Banpurkar, A.G.; Banat, I.M.; Mone, N.S.; Patil, R.H.; Cameotra, S.S. Biosurfactant/s from lactobacilli species: Properties, challenges and potential biomedical applications. J. Basic Microbiol. 2016, 56, 1140–1158. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Oh, S.; Kim, S.H. Released exopolysaccharide (r-EPS) produced from probiotic bacteria reduce biofilm formation of enterohemorrhagic Escherichia coli O157:H7. Biochem. Biophys. Res. Commun. 2009, 379, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Mahdhi, A.; Leban, N.; Chakroun, I.; Bayar, S.; Mahdouani, K.; Majdoub, H.; Kouidhi, B. Use of extracellular polysaccharides, secreted by Lactobacillus plantarum and Bacillus spp., as reducing indole production agents to control biofilm formation and efflux pumps inhibitor in Escherichia coli. Microb. Pathog. 2018, 125, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Høiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Ciofu, O. Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents 2010, 35, 322–332. [Google Scholar] [CrossRef] [Green Version]
- Cerqueira, L.; Oliveira, J.A.; Nicolau, A.; Azevedo, N.F.; Vieira, M.J. Biofilm formation with mixed cultures of Pseudomonas aeruginosa/Escherichia coli on silicone using artificial urine to mimic urinary catheters. Biofouling 2013, 29, 829–840. [Google Scholar] [CrossRef]
- Green, M.R.; Sambrook, J. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor: New York, NY, USA, 1989. [Google Scholar]
- Brooks, T.; Keevil, C.W. A simple artificial urine for the growth of urinary pathogens. Lett. Appl. Microbiol. 1997, 24, 203–206. [Google Scholar] [CrossRef]
- Azeredo, J.; Azevedo, N.F.; Briandet, R.; Cerca, N.; Coenye, T.; Costa, A.R.; Desvaux, M.; Di Bonaventura, G.; Hébraud, M.; Jaglic, Z.; et al. Critical review on biofilm methods. Crit. Rev. Microbiol. 2017, 43, 313–351. [Google Scholar] [CrossRef] [Green Version]
- Moreira, J.M.R.; Gomes, L.C.; Whitehead, K.A.; Lynch, S.; Tetlow, L.A.; Mergulhão, F.J. Effect of surface conditioning with cellular extracts on Escherichia coli adhesion and initial biofilm formation. Food Bioprod. Process. 2017, 104, 1–12. [Google Scholar] [CrossRef]
- Gomes, L.C.; Moreira, J.M.R.; Miranda, J.M.; Simões, M.; Melo, L.F.; Mergulhão, F.J. Macroscale versus microscale methods for physiological analysis of biofilms formed in 96-well microtiter plates. J. Microbiol. Methods 2013, 95, 342–349. [Google Scholar] [CrossRef]
- Heydorn, A.; Nielsen, A.T.; Hentzer, M.; Sternberg, C.; Givskov, M.; Ersboll, B.K.; Molin, S. Quantification of biofilm structures by the novel computer program COMSTAT. Microbiology 2000, 146, 2395–2407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janczuk, B.; Chibowski, E.; Bruque, J.M.; Kerkeb, M.L.; Caballero, F.G. On the consistency of surface free energy components as calculated from contact angles of different liquids: An application to the cholesterol surface. J. Colloid Interface Sci. 1993, 159, 421–428. [Google Scholar] [CrossRef]
- Van Oss, C.J.; Chaudhury, M.K.; Good, R.J. Monopolar surfaces. Adv. Colloid Interface Sci. 1987, 28, 35–64. [Google Scholar] [CrossRef]
- Van Oss, C.J.; Good, R.J.; Chaudhury, M.K. Additive and nonadditive surface tension components and the interpretation of contact angles. Langmuir 1988, 4, 884–891. [Google Scholar] [CrossRef]
- Van Oss, C.J.; Ju, L.; Chaudhury, M.K.; Good, R.J. Estimation of the polar parameters of the surface tension of liquids by contact angle measurements on gels. J. Colloid Interface Sci. 1989, 128, 313–319. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Contact Angle (°) | Hydrophobicity (mJ·m−2) | E. coli–Surface Interaction (mJ·m−2) | ||
|---|---|---|---|---|---|
| Surface | |||||
| Bare silicone | 107.1 ± 4.9 | 100.7 ± 2.4 | 85.8 ± 3.7 | −62.4 | 36.1 |
| L. plantarum biofilm | 34.9 ± 2.8 | 39.5 ± 4.9 | 23.1 ± 2.6 | 34.3 | 84.0 |
| Bacteria | |||||
| E. coli CECT 434 GFP | 19.4 ± 2.2 | 78.1 ± 5.5 | 62.2 ± 4.9 | 131.2 | N.A. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carvalho, F.M.; Teixeira-Santos, R.; Mergulhão, F.J.M.; Gomes, L.C. Effect of Lactobacillus plantarum Biofilms on the Adhesion of Escherichia coli to Urinary Tract Devices. Antibiotics 2021, 10, 966. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10080966
Carvalho FM, Teixeira-Santos R, Mergulhão FJM, Gomes LC. Effect of Lactobacillus plantarum Biofilms on the Adhesion of Escherichia coli to Urinary Tract Devices. Antibiotics. 2021; 10(8):966. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10080966
Chicago/Turabian StyleCarvalho, Fábio M., Rita Teixeira-Santos, Filipe J. M. Mergulhão, and Luciana C. Gomes. 2021. "Effect of Lactobacillus plantarum Biofilms on the Adhesion of Escherichia coli to Urinary Tract Devices" Antibiotics 10, no. 8: 966. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics10080966