
Early Response of Antimicrobial Resistance and Virulence Genes Expression in Classical, Hypervirulent, and Hybrid hvKp-MDR Klebsiella pneumoniae on Antimicrobial Stress


 ,
,
Abstract
:1. Introduction
2. Results
2.1. Bacterial Isolates and Clinical Data
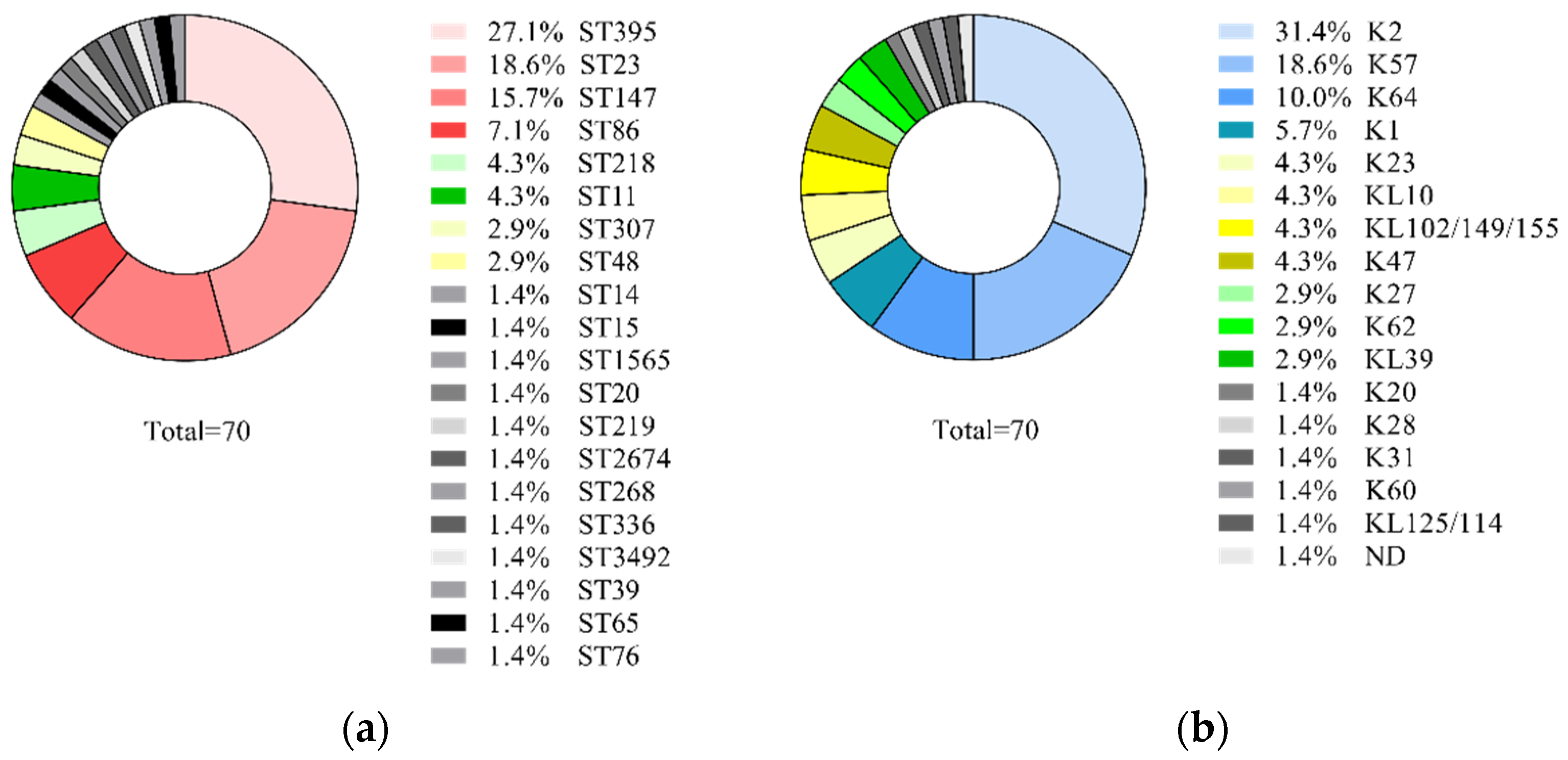
2.2. K. pneumoniae Sequence Types and Capsular Types
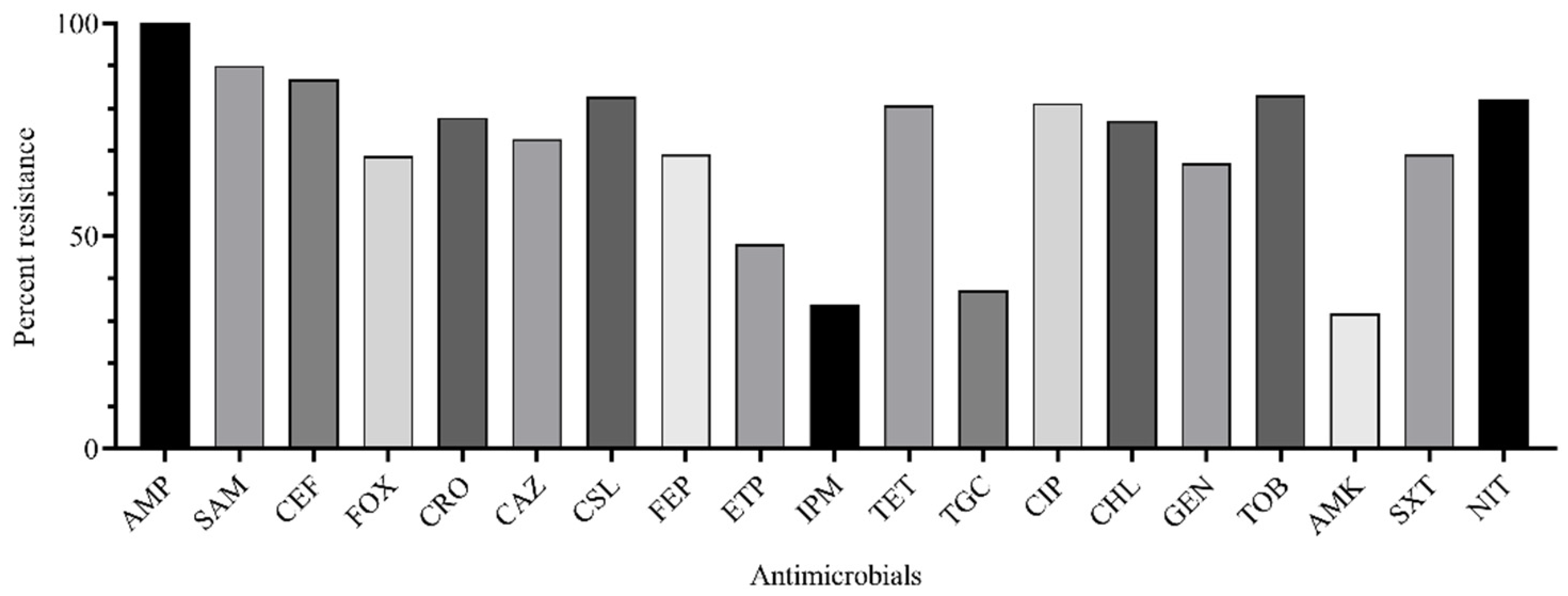
2.3. Susceptibility to Antimicrobials and Resistance Genes
2.4. K. pneumoniae Virulence Genes
2.5. Hypermucoviscousity Phenotype
2.6. Whole-Genome Sequencing
2.7. Relative mRNA Levels of Antimicrobial Resistance and Virulence Genes in K. pneumoniae Cells
2.8. Fold Change of Expression Levels of K. pneumoniae Antimicrobial Resistance and Virulence Genes under AMP and CRO Selective Pressure
3. Discussion
4. Materials and Methods
4.1. Bioethical Requirements and Patients
4.2. Bacterial Isolates, Identification, and Growth Conditions
4.3. Susceptibility to Antimicrobials
4.4. Hypermucoviscousity Testing
4.5. K. pneumoniae Sequence Type and Capsular Type Identification
4.6. Detection of Antimicrobial Resistance and Virulence Genes
4.7. Real-Time PCR Detection of Antimicrobial Resistance and Virulence Genes
4.8. Whole-Genome Sequencing
4.9. K. pneumoniae Growing in Antimicrobial Conditions and RNA Isolation
4.10. cDNA Synthesis and Quantitative Real-Time PCR
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Bengoechea, J.A.; Sa Pessoa, J. Klebsiella Pneumoniae Infection Biology: Living to Counteract Host Defences. FEMS Microbiol. Rev. 2019, 43, 123–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bialek-Davenet, S.; Criscuolo, A.; Ailloud, F.; Passet, V.; Jones, L.; Delannoy-Vieillard, A.S.; Garin, B.; le Hello, S.; Arlet, G.; Nicolas-Chanoine, M.H.; et al. Genomic Definition of Hypervirulent and Multidrug-Resistant Klebsiella Pneumoniae Clonal Groups. Emerg. Infect. Dis. 2014, 20, 1812–1820. [Google Scholar] [CrossRef]
- Baraniak, A.; Izdebski, R.; Fiett, J.; Sadowy, E.; Adler, A.; Kazma, M.; Salomon, J.; Lawrence, C.; Rossini, A.; Salvia, A.; et al. Comparative Population Analysis of Klebsiella Pneumoniae Strains with Extended-Spectrum β-Lactamases Colonizing Patients in Rehabilitation Centers in Four Countries. Antimicrob. Agents Chemother. 2013, 57, 1992–1997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Struve, C.; Roe, C.C.; Stegger, M.; Stahlhut, S.G.; Hansen, D.S.; Engelthaler, D.M.; Andersen, P.S.; Driebe, E.M.; Keim, P.; Krogfelt, K.A. Mapping the Evolution of Hypervirulent Klebsiella Pneumoniae. mBio 2015, 6, e00630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choby, J.E.; Howard-Anderson, J.; Weiss, D.S. Hypervirulent Klebsiella Pneumoniae—Clinical and Molecular Perspectives. J. Intern. Med. 2020, 287, 283–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, M.; Yang, X.; Xu, Q.; Ye, L.; Chen, K.; Zheng, Z.; Dong, N.; Sun, Q.; Shu, L.; Gu, D.; et al. Clinical Evolution of ST11 Carbapenem Resistant and Hypervirulent Klebsiella Pneumoniae. Commun. Biol. 2021, 4, 650. [Google Scholar] [CrossRef]
- Liu, B.T.; Su, W.Q. Whole Genome Sequencing of NDM-1-Producing Serotype K1 St23 Hypervirulent Klebsiella Pneumoniae in China. J. Med. Microbiol. 2019, 68, 866–873. [Google Scholar] [CrossRef] [PubMed]
- Turton, J.; Davies, F.; Turton, J.; Perry, C.; Payne, Z.; Pike, R. Hybrid Resistance and Virulence Plasmids in “High-Risk” Clones of Klebsiella Pneumoniae, Including Those Carrying Blandm-5. Microorganisms 2019, 7, 326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ershova, K.; Savin, I.; Kurdyumova, N.; Wong, D.; Danilov, G.; Shifrin, M.; Alexandrova, I.; Sokolova, E.; Fursova, N.; Zelman, V.; et al. Implementing an Infection Control and Prevention Program Decreases the Incidence of Healthcare-Associated Infections and Antibiotic Resistance in a Russian Neuro-ICU. Antimicrob. Resist. Infect. Control 2018, 7, 94. [Google Scholar] [CrossRef]
- Fursova, N.K.; Astashkin, E.I.; Ershova, O.N.; Aleksandrova, I.A.; Savin, I.A.; Novikova, T.S.; Fedyukina, G.N.; Kislichkina, A.A.; Fursov, M.V.; Kuzina, E.S.; et al. Multidrug-Resistant Klebsiella Pneumoniae Causing Severe Infections in the Neuro-Icu. Antibiotics 2021, 10, 979. [Google Scholar] [CrossRef]
- Jain, N.; Jansone, I.; Obidenova, T.; Sīmanis, R.; Meisters, J.; Straupmane, D.; Reinis, A. Epidemiological Characterization of Clinical Fungal Isolates from Pauls Stradinš Clinical University Hospital, Latvia: A 4-Year Surveillance Report. Life 2021, 11, 1002. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, K.H.; Chen, W.; Yu, Y.; Feng, S.F. Epidemiology and Risk Factors for Nosocomial Infection in the Respiratory Intensive Care Unit of a Teaching Hospital in China: A Prospective Surveillance during 2013 and 2015. BMC Infect. Dis. 2019, 19, 145. [Google Scholar] [CrossRef] [Green Version]
- Fursova, N.K.; Astashkin, E.I.; Knyazeva, A.I.; Kartsev, N.N.; Leonova, E.S.; Ershova, O.N.; Alexandrova, I.A.; Kurdyumova, N.V.; Sazikina, S.Y.; Volozhantsev, N.V.; et al. The spread of bla OXA-48 and bla OXA-244 carbapenemase genes among Klebsiella pneumoniae, Proteus mirabilis and Enterobacter spp. isolated in Moscow, Russia. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 46. [Google Scholar] [CrossRef] [Green Version]
- Izdebski, R.; Baraniak, A.; Zabicka, D.; Machulska, M.; Urbanowicz, P.; Fiett, J.; Literacka, E.; Bojarska, K.; Kozińska, A.; Zieniuk, B.; et al. Enterobacteriaceae Producing OXA-48-like Carbapenemases in Poland, 2013-January 2017. J. Antimicrob. Chemother. 2018, 73, 620–625. [Google Scholar] [CrossRef]
- Muggeo, A.; Guillard, T.; Klein, F.; Reffuveille, F.; François, C.; Babosan, A.; Bajolet, O.; Bertrand, X.; de Champs, C. Spread of Klebsiella Pneumoniae ST395 Non-Susceptible to Carbapenems and Resistant to Fluoroquinolones in North-Eastern France. J. Glob. Antimicrob. Resist. 2018, 13, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Maida, C.M.; Bonura, C.; Geraci, D.M.; Graziano, G.; Carattoli, A.; Rizzo, A.; Torregrossa, M.V.; Vecchio, D.; Giuffrè, M. Outbreak of ST395 KPC-Producing Klebsiella Pneumoniae in a Neonatal Intensive Care Unit in Palermo, Italy. Infect. Control Hosp. Epidemiol. 2018, 39, 496–498. [Google Scholar] [CrossRef] [Green Version]
- Fursova, N.K.; Astashkin, E.I.; Gabrielyan, N.I.; Novikova, T.S.; Fedyukina, G.N.; Kubanova, M.K.; Esenova, N.M.; Sharapchenko, S.O.; Volozhantsev, N.V. Emergence of Five Genetic Lines ST395NDM-1, ST13OXA-48, ST3346OXA-48, ST39CTX-M-14, and Novel ST3551OXA-48of Multidrug-Resistant Clinical Klebsiella Pneumoniae in Russia. Microb. Drug Resist. 2020, 26, 924–933. [Google Scholar] [CrossRef]
- Peirano, G.; Chen, L.; Kreiswirth, B.N.; Pitout, J.D.D. Emerging Antimicrobial-Resistant High-Risk Klebsiella Pneumoniae Clones ST307 and ST147. Antimicrob. Agents Chemother. 2020, 64, e01148-20. [Google Scholar] [CrossRef]
- Shankar, C.; Jacob, J.J.; Vasudevan, K.; Biswas, R.; Manesh, A.; Sethuvel, D.P.M.; Varughese, S.; Biswas, I.; Veeraraghavan, B. Emergence of Multidrug Resistant Hypervirulent ST23 Klebsiella Pneumoniae: Multidrug Resistant Plasmid Acquisition Drives Evolution. Front. Cell. Infect. Microbiol. 2020, 10, 575289. [Google Scholar] [CrossRef] [PubMed]
- Baron, S.A.; Pascale, L.M.; Million, M.; Briantais, A.; Durand, J.M.; Hadjadj, L.; Rolain, J.M. Whole Genome Sequencing to Decipher the Virulence Phenotype of Hypervirulent Klebsiella Pneumoniae Responsible for Liver Abscess, Marseille, France. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 1073–1077. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.H.; Huang, Y.T.; Chang, C.Y.; Hsu, H.S.; Hsueh, P.R. Capsular Serotypes and Multilocus Sequence Types of Bacteremic Klebsiella Pneumoniae Isolates Associated with Different Types of Infections. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 365–369. [Google Scholar] [CrossRef]
- Lev, A.I.; Astashkin, E.I.; Kislichkina, A.A.; Solovieva, E.V.; Kombarova, T.I.; Korobova, O.V.; Ershova, O.N.; Alexandrova, I.A.; Malikov, V.E.; Bogun, A.G.; et al. Comparative Analysis of Klebsiella Pneumoniae Strains Isolated in 2012–2016 That Differ by Antibiotic Resistance Genes and Virulence Genes Profiles. Pathog. Glob. Health 2018, 112, 142–151. [Google Scholar] [CrossRef]
- Shon, A.S.; Bajwa, R.P.S.; Russo, T.A. Hypervirulent (Hypermucoviscous) Klebsiella Pneumoniae: A New and Dangerous Breed. Virulence 2013, 4, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Jin, L.; Ouyang, P.; Wang, Q.; Wang, R.; Wang, J.; Gao, H.; Wang, X.; Wang, H. Evolution of Hypervirulence in Carbapenem-Resistant Klebsiella Pneumoniae in China: A Multicentre, Molecular Epidemiological Analysis. J. Antimicrob. Chemother. 2020, 75, 327–336. [Google Scholar] [CrossRef]
- Wyres, K.L.; Lam, M.M.C.; Holt, K.E. Population Genomics of Klebsiella Pneumoniae. Nat. Rev. Microbiol. 2020, 18, 344–359. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Dong, N.; Chen, K.; Yang, X.; Ye, L.; Chan, E.W.C.; Zhang, R.; Chen, S. A Hybrid Plasmid Formed by Recombination of a Virulence Plasmid and a Resistance Plasmid in Klebsiella Pneumoniae. J. Glob. Antimicrob. Resist. 2020, 23, 466–470. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Li, Y.; Wang, G.; Li, C.; Chang, Y.F.; Chen, W.; Nian, S.; Mao, Y.; Zhang, J.; Zhong, F.; et al. Bla NDM-5 Carried by a Hypervirulent Klebsiella Pneumoniae with Sequence Type 29. Antimicrob. Resist. Infect. Control 2019, 8, 140. [Google Scholar] [CrossRef]
- Mohammad Ali Tabrizi, A.; Badmasti, F.; Shahcheraghi, F.; Azizi, O. Outbreak of Hypervirulent Klebsiella Pneumoniae Harbouring BlaVIM-2 among Mechanically-Ventilated Drug-Poisoning Patients with High Mortality Rate in Iran. J. Glob. Antimicrob. Resist. 2018, 15, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Lazareva, I.; Ageevets, V.; Sopova, J.; Lebedeva, M.; Starkova, P.; Likholetova, D.; Lebedeva, M.; Gostev, V.; Moiseenko, V.; Egorenkov, V.; et al. The Emergence of Hypervirulent BlaNDM-1-Positive Klebsiella Pneumoniae Sequence Type 395 in an Oncology Hospital. Infect. Genet. Evol. 2020, 85, 104527. [Google Scholar] [CrossRef]
- Hou, X.H.; Song, X.Y.; Ma, X.B.; Zhang, S.Y.; Zhang, J.Q. Molecular Characterization of Multidrug-Resistant Klebsiella Pneumoniae Isolates. Braz. J. Microbiol. 2015, 46, 759–768. [Google Scholar] [CrossRef] [Green Version]
- Farhadi, M.; Ahanjan, M.; Goli, H.R.; Haghshenas, M.R.; Gholami, M. High Frequency of Multidrug-Resistant (MDR) Klebsiella Pneumoniae Harboring Several β-Lactamase and Integron Genes Collected from Several Hospitals in the North of Iran. Ann. Clin. Microbiol. Antimicrob. 2021, 20, 70. [Google Scholar] [CrossRef]
- Lin, Z.W.; Zheng, J.X.; Bai, B.; Xu, G.J.; Lin, F.J.; Chen, Z.; Sun, X.; Qu, D.; Yu, Z.J.; Deng, Q.W. Characteristics of Hypervirulent Klebsiella Pneumoniae: Does Low Expression of RmpA Contribute to the Absence of Hypervirulence? Front. Microbiol. 2020, 11, 436. [Google Scholar] [CrossRef]
- Protonotariou, E.; Meletis, G.; Chatzopoulou, F.; Malousi, A.; Chatzidimitriou, D.; Skoura, L. Emergence of Klebsiella Pneumoniae ST11 Co-Producing NDM-1 and OXA-48 Carbapenemases in Greece. J. Glob. Antimicrob. Resist. 2019, 19, 81–82. [Google Scholar] [CrossRef] [PubMed]
- Hernández, M.; López-Urrutia, L.; Abad, D.; Serna, M.D.F.; Ocampo-Sosa, A.A.; Eiros, J.M. First Report of an Extensively Drug-Resistant ST23 Klebsiella Pneumoniae of Capsular Serotype K1 Co-Producing CTX-M-15, OXA-48 and ArmA in Spain. Antibiotics 2021, 10, 157. [Google Scholar] [CrossRef]
- Carvalho, I.; Carvalho, J.A.; Martínez-álvarez, S.; Sadi, M.; Capita, R.; Alonso-Calleja, C.; Rabbi, F.; de Lurdes Nunes Enes Dapkevicius, M.; Igrejas, G.; Torres, C.; et al. Characterization of Esbl-Producing Escherichia Coli and Klebsiella Pneumoniae Isolated from Clinical Samples in a Northern Portuguese Hospital: Predominance of Ctx-m-15 and High Genetic Diversity. Microorganisms 2021, 9, 1914. [Google Scholar] [CrossRef] [PubMed]
- Chaves, J.; Ladona, M.G.; Segura, C.; Coira, A.; Reig, R.; Ampurdanés, C. SHV-1 β-Lactamase Is Mainly a Chromosomally Encoded Species-Specific Enzyme in Klebsiella Pneumoniae. Antimicrob. Agents Chemother. 2001, 45, 2856–2861. [Google Scholar] [CrossRef] [Green Version]
- Awosile, B.B.; Agbaje, M. Genetic Environments of Plasmid-Mediated BlaCTXM-15 Beta-Lactamase Gene in Enterobacteriaceae from Africa. Microbiol. Res. 2021, 12, 26. [Google Scholar] [CrossRef]
- Barguigua, A.; el Otmani, F.; Talmi, M.; Reguig, A.; Jamali, L.; Zerouali, K.; Timinouni, M. Prevalence and Genotypic Analysis of Plasmid-Mediated β-Lactamases among Urinary Klebsiella Pneumoniae Isolates in Moroccan Community. J. Antibiot. 2013, 66, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Shi, W.; Qin, J.; Mi, Z. A Klebsiella pneumoniaesputum culture isolate from China carrying blaOXA-1, blaCTX-M-55 and aac(69)-Ib-cr. J. Med. Microbiol. 2008, 57, 1588–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherchan, J.B.; Tada, T.; Shrestha, S.; Uchida, H.; Hishinuma, T.; Morioka, S.; Shahi, R.K.; Bhandari, S.; Twi, R.T.; Kirikae, T.; et al. Emergence of Clinical Isolates of Highly Carbapenem-Resistant Klebsiella Pneumoniae Co-Harboring BlaNDM-5 and BlaOXA-181 or -232 in Nepal. Int. J. Infect. Dis. 2020, 92, 247–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurya, N.; Jangra, M.; Tambat, R.; Nandanwar, H. Alliance of Efflux Pumps with β-Lactamases in Multidrug-Resistant Klebsiella Pneumoniae Isolates. Microb. Drug Resist. 2019, 25, 1155–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirzaie, A.; Ranjbar, R. Antibiotic Resistance, Virulence-Associated Genes Analysis and Molecular Typing of Klebsiella Pneumoniae Strains Recovered from Clinical Samples. AMB Express 2021, 11, 122. [Google Scholar] [CrossRef]
- Russo, T.A.; Olson, R.; Fang, C.T.; Stoesser, N.; Miller, M.; MacDonald, U.; Hutson, A.; Barker, J.H.; la Hoz, R.M.; Johnson, J.R.; et al. Identification of Biomarkers for Differentiation of Hypervirulent Klebsiella Pneumoniae from Classical K. Pneumoniae. J. Clin. Microbiol. 2018, 56, e00776-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, T.A.; MacDonald, U.; Hassan, S.; Camanzo, E.; LeBreton, F.; Corey, B.; McGann, P. An Assessment of Siderophore Production, Mucoviscosity, and Mouse Infection Models for Defining the Virulence Spectrum of Hypervirulent Klebsiella Pneumoniae. mSphere 2021, 6, e00045-21. [Google Scholar] [CrossRef] [PubMed]
- Shaidullina, E.; Shelenkov, A.; Yanushevich, Y.; Mikhaylova, Y.; Shagin, D.; Alexandrova, I.; Ershova, O.; Akimkin, V.; Kozlov, R.; Edelstein, M. Antimicrobial Resistance and Genomic Characterization of OXA-48-and CTX-M-15-Co-Producing Hypervirulent Klebsiella Pneumoniae St23 Recovered from Nosocomial Outbreak. Antibiotics 2020, 9, 862. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, F.; Qarasnji, B.K.; Abdullah Tawgozy, F.H.; Amin, B.K. Molecular Study of SHV-11 and SHV-12 Genes among Klebsiella Pneumoniae Isolated from UTI Patients in Erbil City. Zanco J. Pure Appl. Sci. 2016, 28, 25–35. [Google Scholar]
- Pal, A.; Dhara, L.; Tripathi, A. Contribution of AcrB Upregulation & OmpC/Ompk36 Loss over the Presence of BlaNDM towards Carbapenem Resistance Development among Pathogenic Escherichia Coli & Klebsiella Spp. Indian J. Med. Res. 2019, 149, 528–538. [Google Scholar] [CrossRef]
- Hennequin, C.; Robin, F. Correlation between Antimicrobial Resistance and Virulence in Klebsiella Pneumoniae. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 333–341. [Google Scholar] [CrossRef]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Diancourt, L.; Passet, V.; Verhoef, J.; Grimont, P.A.D.; Brisse, S. Multilocus Sequence Typing of Klebsiella Pneumoniae Nosocomial Isolates. J. Clin. Microbiol. 2005, 43, 4178–4182. [Google Scholar] [CrossRef] [Green Version]
- Brisse, S.; Fevre, C.; Passet, V.; Issenhuth-Jeanjean, S.; Tournebize, R.; Diancourt, L.; Grimont, P. Virulent Clones of Klebsiella Pneumoniae: Identification and Evolutionary Scenario Based on Genomic and Phenotypic Characterization. PLoS ONE 2009, 4, e4982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, C.T.; Lai, S.Y.; Yi, W.C.; Hsueh, P.R.; Liu, K.L.; Chang, S.C. Klebsiella Pneumoniae Genotype K1: An Emerging Pathogen That Causes Septic Ocular or Central Nervous System Complications from Pyogenic Liver Abscess. Clin. Infect. Dis. 2007, 45, 284–293. [Google Scholar] [CrossRef]
- Brisse, S.; Passet, V.; Haugaard, A.B.; Babosan, A.; Kassis-Chikhani, N.; Struve, C.; Decre, D. Wzi Gene Sequencing, a Rapid Method for Determination of Capsulartype for Klebsiella Strains. J. Clin. Microbiol. 2013, 51, 4073–4078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bikandi, J.; Millán, R.S.; Rementeria, A.; Garaizar, J. In Silico Analysis of Complete Bacterial Genomes: PCR, AFLP-PCR and Endonuclease Restriction. Bioinformatics 2004, 20, 798–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, A.É.I.; Stuchi, L.P.; Siqueira, N.M.G.; Henrique, J.B.; Vicentini, R.; Ribeiro, M.L.; Darrieux, M.; Ferraz, L.F.C. Selection and Validation of Reference Genes for Gene Expression Studies in Klebsiella Pneumoniae Using Reverse Transcription Quantitative Real-Time PCR. Sci. Rep. 2018, 8, 9001. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2-ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feature/strain | B185/19 | B16K/19 | B2523/18 | B2580/14 | B1261/19 |
|---|---|---|---|---|---|
| SCPM-O | SCPM-O-B-9222 | SCPM-O-B-9221 | SCPM-O-B-9220 | SCPM-O-B-7852 | SCPM-O-B-9223 |
| Isolation date | 30-Jan-2019 | 09-Jan-2019 | 30-Aug-2018 | 21-Jul-2014 | 27-May-2019 |
| Source | wound | urine | blood | urine | urine |
| GenBank | JAGVVR000000000.1 | JAGVVS000000000.1 | JAGRZF000000000.1 | PUXF00000000.1 | JAGVVQ000000000.1 |
| Genome size, bp | 5,812,311 | 5,532,115 | 5,430,135 | 5,773,846 | 5,570,452 |
| Number of contigs | 267 | 180 | 125 | 360 | 160 |
| Genetic line | ST395K47 | ST147K64 | ST86K2 | ST23K1 | ST23K57 |
| String-test | Negative | Negative | Positive | Positive | Positive |
| Antimicrobial resistance phenotype | AMP, SAM, CXM, FOX, CRO, CAZ, CSL, FEP, ETP, IPM, TET, CIP, CHL, GEN, TOB, AMK, SXT, NIT | AMP, SAM, CXM, FOX, CRO, CAZ, CSL, FEP, ETP, IPM, CIP, GEN, TOB, AMK, SXT, NIT | AMP | AMP, SAM, CXM, CRO, TET, CIP, CHL, TOB, SXT, NIT | AMP, SAM, CXM, FOX, CRO, CAZ, CSL, FEP, ETP, IPM, TET, TGC, CIP, CHL, GEN, TOB, SXT, NIT |
| Beta-lactamases | blaSHV-12, blaTEM-1B, blaCTX-M-15, blaOXA-1, blaOXA-48, blaNDM-1 | blaSHV-67, blaTEM-1B, blaCTX-M-15, blaOXA-1, blaOXA-48, blaNDM-1 | blaSHV-28 | blaSHV-190, blaTEM-1B, blaCTX-M-15, blaOXA-1, blaOXA-48 | blaSHV-33, blaCTX-M-55, blaOXA-1, blaOXA-48, blaNDM-1 |
| AG-resistance | aadA2, aph(3′)-Ia, armA | aph(3′)-VI, aadA1, aadA2, aadA5, armA | - | aph(6)-Id, aac(6′)-Ib-cr, aph(3″)-Ib | aph(6)-Id, aph(3′)-Ia, aph(3″)-Ib, aac(3)-IIa, aac(3)-IId, aadA16 |
| FOS-resistance | fosA | fosA | fosA | fosA | fosA |
| STR-resistance | msr(E) | msr(E) | - | - | |
| PHE-resistance | catB3 | catB3 | - | catB3 | catB3, floR |
| QNL-resistance | aac(6′)-Ib-cr, oqxA, oqxB | aac(6′)-Ib-cr, oqxA, oqxB | oqxA, oqxB | qnrB1, oqxA, oqxB | aac(6′)-Ib-cr, oqxA, oqxB |
| SUL-resistance | sul1 | sul1 | - | sul2 | sul1, sul2 |
| TET-resistance | tet(A) | - | - | tet(A) | tet(A) |
| TRI-resistance | dfrA1, dfrA12 | dfrA12, dfrA17. | - | dfrA14 | dfrA27 |
| MCR-resistance | mph(A), mph(E) | mph(E) | - | - | mph(A) |
| RIF-resistance | - | - | - | - | ARR-3 |
| HM-resistance | ars, pco, sil | sil | pbr, pco, sil, ter | pbr, pco, sil, ter | pbr, pco, sil, ter |
| Efflux pums | acr, oqx, kpn, kde, kex | acr, oqx, kpn, kde, kex | acr, oqx, kpn, kde, kex | acr, oqx, kpn, kde | acr, oqx, kpn, kde, kex |
| Virulence genes | uge, wabG, fimH, irp, iut, mrk, ybt, fyu,treC, cellB, ureA | uge, wabG, fimH, mrk,treC, cellB, ureA | peg-344, rmpA, iro, ent uge, wabG, fimH, irp, iuc, iut, mrk, ybt, fyu,treC, cellB, ureA | peg-344, rmpA, rmpA2, iro, ent, uge, wabG, kfu, fimH, allR, irp, iuc, iut, mrk, ybt, fyu,treC, cellB, ureA | peg-344, rmpA, ent, wabG, fimH, irp, iuc, iut, mrk, ybt, fyu,treC, cellB, ureA |
| Plasmid | ColRNAI, IncFIB(K), IncFII(K), IncR, IncX3 | IncFIA(HI1), IncFIB(K), IncFIB, IncFII(K), IncHI1B, IncM2 | IncHI1B, repB | IncFII(K), IncHI1B, IncL, repB | ColRNAI, IncC, IncFIA(HI1), IncFII, IncFII(K), IncL, repB |
| Target Gene | Forward Primer (5′→3′) | Reverse Primer (5′→3′) | Size of PCR Product, bp |
|---|---|---|---|
| proC * | GATTGCCGATATCGTCTTCG | GAGACCACCAGCGACTCTTT | 99 |
| recA * | TTAAACAGGCCGAATTCCAG | CCGCTTTCTCAATCAGCTTC | 99 |
| rpoD * | TCCGGTGCATATGATTGAGA | ATACGCTCAGCCAGCTCTTC | 105 |
| blaTEM | GCCAACTTACTTCTGACAACGA | GATCAAGGCGAGTTACATG | 88 |
| blaSHV | GCCATTACCATGAGCGATAA | CAAAAAGGCAGTCAATCC | 81 |
| blaCTX-M | AAGCAGAGTGAAACGCAAAA | GGCAATCGGATTGTAGTTAA | 84 |
| blaOXA-48 | GTGTTTTTGGTGGCATCGATTA | GGTTCGCCCGTTTAAGATTATT | 164 |
| blaNDM | ACTTTGGCCCGCTCAAGGTATT | TGATCAGGCAGCCACCAAAA | 105 |
| ompK36 | CGGTTACGGCCAGTGGGAATA | GAGCCCGCGTCGCCAAATTT | 111 |
| acrA | ATGTGACGATAAACCGGCTC | CTGGCAGTTCGGTGGTTATT | 101 |
| acrB | TGGTGCTGCATGGCCAGGTT | CTTGTCCACCAGCCGCGAAA | 149 |
| oqxA | AGGCGCTTCCGGTGGAGATT | GCAAACAGCCCTGGCGTGAA | 160 |
| oqxB | CGCAACTACGCCACGCTGAA | GCCAGACGCGCATCGCATAT | 103 |
| kpnE | ATTGCTGAAATTACCGGCAC | AAATACCGATCCCTTCCCAC | 172 |
| kpnF | ATAACCCCCGCCCAGCCTTT | CCCTGTCGCAGGCGGTTAAA | 139 |
| kexD | TGAAGTTGCGCCACCCGGTA | TGTCGGTGCCGGGTTTGAAA | 191 |
| kdeA | CAGGCGCTGTTGTTCCCGTT | AAACATGCCGCCTGCCAGAT | 162 |
| rmpA | TGTACCTTTTGCAGCCAA | CTGCAATGCCTACTGAAAA | 121 |
| iroN | TTCCGGCTGGTTGGTGTATA | ATCGAAGTGATCCGTGGCC | 132 |
| iroD | ATAAGTTTGGCCGGCAGG | TTCGTGCCGAACAACCGC | 183 |
| uge | CCCGGATATGGCGCTGTTCA | CGCTTCATTTTGCCGTAGTT | 84 |
| wabG | AACGTATTCCCGGCTGCGATCT | CCGCGCAGGTGTGAGTCTTCAT | 175 |
| fimH | GCAGGATCAGCACCGCGATTAA | TCACGGACCGATAAACCCTGGC | 106 |
| allR | GAGCCTTCCGCGCAAAA | CCGCGCCTTCCAGCACTTTTA | 163 |
| treC | CCATTTCTCTCCGCCGGGATAA | ACCTGATGACCGTCGGCGAGAT | 137 |
| celB | GCGCGACGGCATCATTCT | GCGATGCCGAAGCTGGAAAT | 192 |
| ureA | GAGGGCGTCCCGGAAAT | TGTGAACGGTGACCAGCTT | 79 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fursova, A.D.; Fursov, M.V.; Astashkin, E.I.; Novikova, T.S.; Fedyukina, G.N.; Kislichkina, A.A.; Alexandrova, I.A.; Ershova, O.N.; Dyatlov, I.A.; Fursova, N.K. Early Response of Antimicrobial Resistance and Virulence Genes Expression in Classical, Hypervirulent, and Hybrid hvKp-MDR Klebsiella pneumoniae on Antimicrobial Stress. Antibiotics 2022, 11, 7. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics11010007
Fursova AD, Fursov MV, Astashkin EI, Novikova TS, Fedyukina GN, Kislichkina AA, Alexandrova IA, Ershova ON, Dyatlov IA, Fursova NK. Early Response of Antimicrobial Resistance and Virulence Genes Expression in Classical, Hypervirulent, and Hybrid hvKp-MDR Klebsiella pneumoniae on Antimicrobial Stress. Antibiotics. 2022; 11(1):7. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics11010007
Chicago/Turabian StyleFursova, Anastasiia D., Mikhail V. Fursov, Evgenii I. Astashkin, Tatiana S. Novikova, Galina N. Fedyukina, Angelina A. Kislichkina, Irina A. Alexandrova, Olga N. Ershova, Ivan A. Dyatlov, and Nadezhda K. Fursova. 2022. "Early Response of Antimicrobial Resistance and Virulence Genes Expression in Classical, Hypervirulent, and Hybrid hvKp-MDR Klebsiella pneumoniae on Antimicrobial Stress" Antibiotics 11, no. 1: 7. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics11010007