

Effects of Coleus amboinicus L. Essential Oil and Ethanolic Extracts on Planktonic Cells and Biofilm Formation of Microsporum canis Isolated from Feline Dermatophytosis

 , ,
, ,
Abstract
:1. Introduction
2. Results
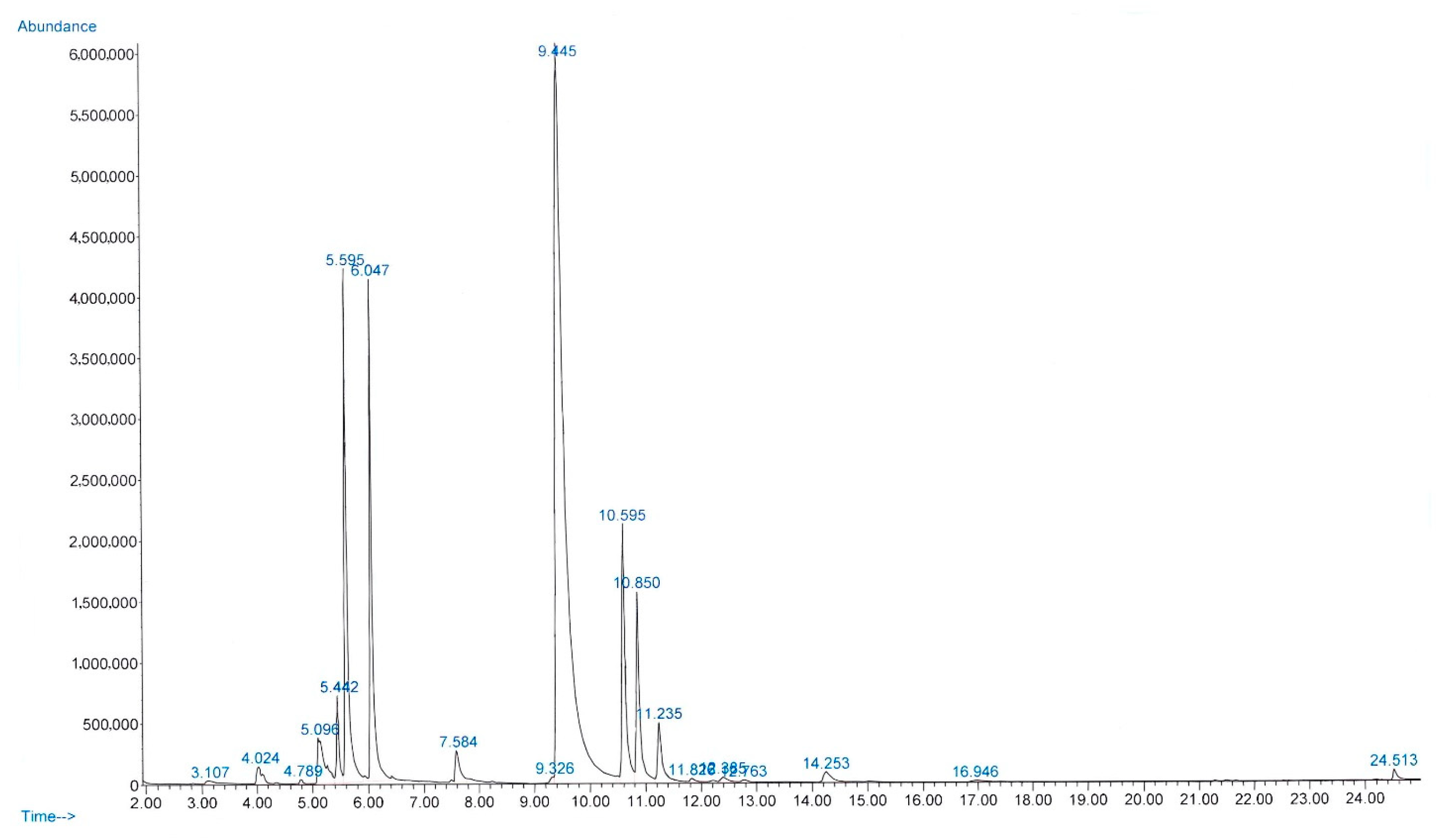
2.1. Chemical Composition of CEO and CEE
2.2. Fungal Isolation and Biofilm Formation
2.3. Effect of CEO and CEE on Planktonic Cells
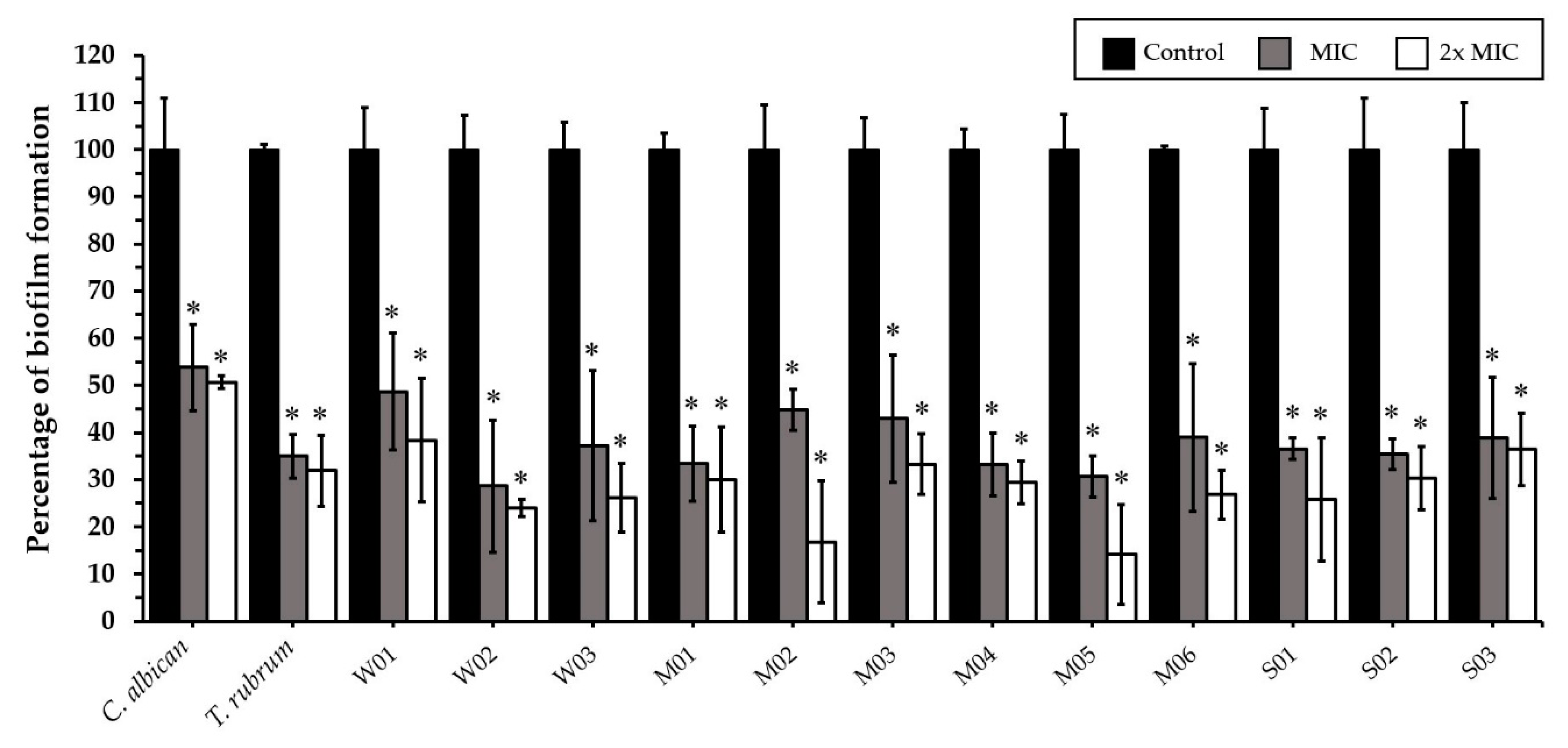
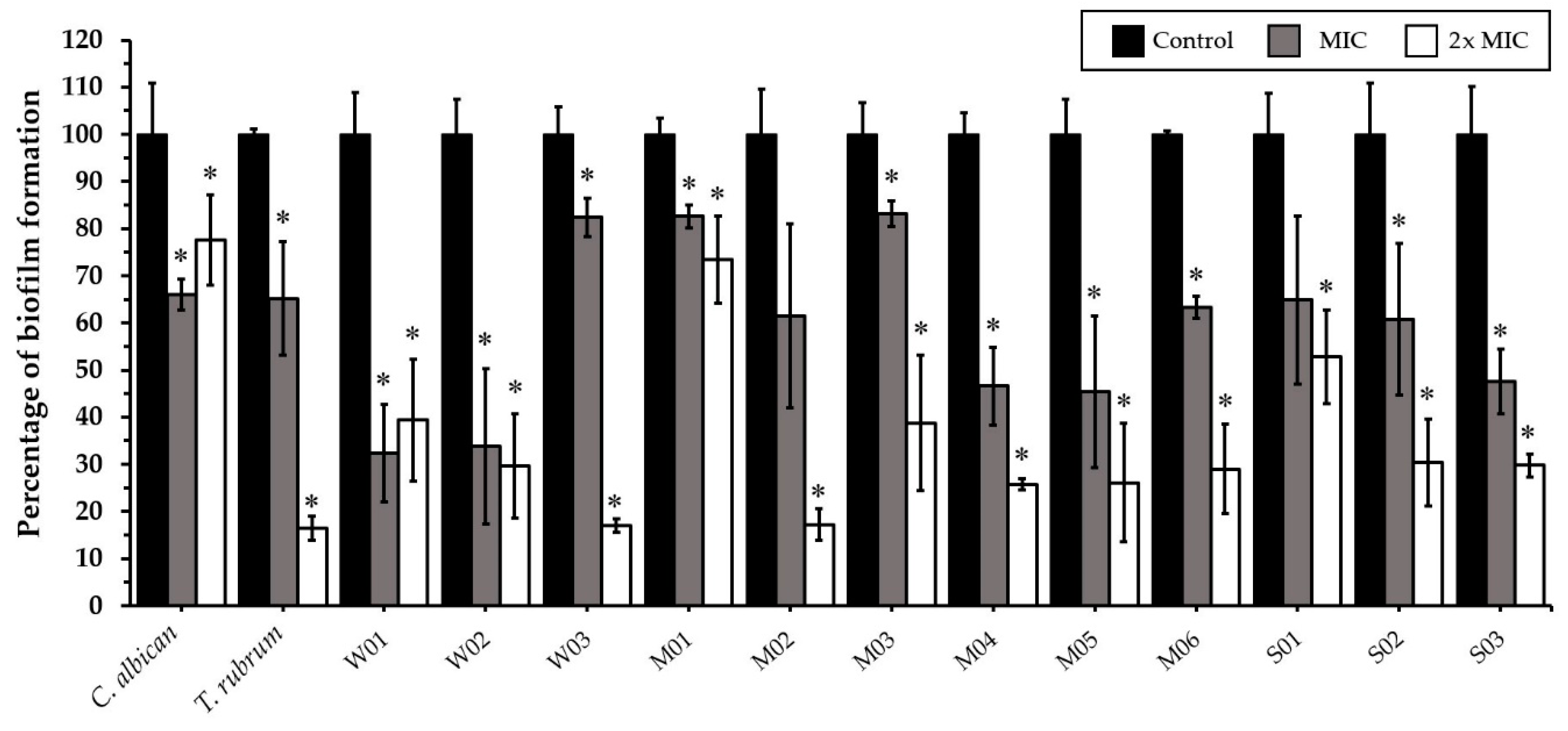
2.4. Effect of CEO and CEE on Biofilm Formation
3. Discussion
4. Materials and Methods
4.1. Plant Preparation and Extractions
4.2. GC-MS
4.3. Determination of Total Phenolic Content
4.4. Determination of Flavonoid Content
4.5. HPLC
4.6. Sample Collection and Fungal Identification
4.7. Confirmation of M. canis Using Molecular Techniques
4.8. Bioinformatic and Phylogenetic Analysis
4.9. Biofilm Formation Classification
4.10. Effects of CEO and CEE on Planktonic Cells
4.11. Effects of CEO and CEE on the Biofilm Formation of M. canis
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moriello, K.A.; Coyner, K.; Paterson, S.; Mignon, B. Diagnosis and treatment of dermatophytosis in dogs and cats.: Clinical Consensus Guidelines of the World Association for Veterinary Dermatology. Vet. Dermatol. 2017, 28, 266-e68. [Google Scholar]
- Gnat, S.; Nowakiewicz, A.; Łagowski, D.; Zięba, P. Host- and pathogen-dependent susceptibility and predisposition to dermatophytosis. J. Med. Microbiol. 2019, 68, 823–836. [Google Scholar]
- Ginter-Hanselmayer, G.; Weger, W.; Ilkit, M.; Smolle, J. Epidemiology of tinea capitis in Europe: Current state and changing patterns. Mycoses 2007, 50, 6–13. [Google Scholar]
- Uhrlaß, S.; Krüger, C.; Nenoff, P. Microsporum canis: Current data on the prevalence of the zoophilic dermatophyte in central Germany. Hautarzt 2015, 66, 855–862. [Google Scholar]
- Pasquetti, M.; Min, A.R.M.; Scacchetti, S.; Dogliero, A.; Peano, A. Infection by Microsporum canis in paediatric patients: A veterinary perspective. Vet. Sci. 2017, 4, 46. [Google Scholar]
- Bond, R. Superficial veterinary mycoses. Clin. Dermatol. 2010, 28, 226–236. [Google Scholar]
- Chupia, V.; Ninsuwon, J.; Piyarungsri, K.; Sodarat, C.; Prachasilchai, W.; Suriyasathaporn, W.; Pikulkaew, S. Prevalence of Microsporum canis from pet cats in Small Animal Hospitals, Chiang Mai, Thailand. Vet. Sci. 2022, 9, 21. [Google Scholar]
- Cafarchia, C.; Romito, D.; Capelli, G.; Guillot, J.; Otranto, D. Isolation of Microsporum canis from the hair coat of pet dogs and cats belonging to owners diagnosed with M. canis tinea corporis. Vet. Dermatol. 2006, 17, 327–331. [Google Scholar]
- Brosh-Nissimov, T.; Ben-Ami, R.; Astman, N.; Malin, A.; Baruch, Y.; Galor, I. An outbreak of Microsporum canis infection at a military base associated with stray cat exposure and person-to-person transmission. Mycoses 2018, 61, 472–476. [Google Scholar]
- Yamada, S.; Aizawa, K.; Mochizuki, T. An epidemiological study of feline and canine dermatophytoses in Japan. Med. Mycol. J. 2019, 60, 39–44. [Google Scholar]
- Uma, K.; Huang, X.; Kumar, B.A. Antifungal effect of plant extract and essential oil. Chin. J. Integr. Med. 2017, 23, 233–239. [Google Scholar]
- Aneke, C.I.; Otranto, D.; Cafarchia, C. Therapy and antifungal susceptibility profile of Microsporum canis. J. Fungi 2018, 4, 107. [Google Scholar]
- Hsiao, Y.H.; Chen, C.; Han, H.S.; Kano, R. The first report of terbinafine resistance Microsporum canis from a cat. J. Vet. Med. Sci. 2018, 80, 898–900. [Google Scholar]
- Cowen, L.E.; Sanglard, D.; Howard, S.J.; Rogers, P.D.; Perlin, D.S. Mechanisms of antifungal drug resistance. Cold Spring Harb. Perspect. Med. 2014, 5, a019752. [Google Scholar]
- Martinez-Rossi, N.M.; Bitencourt, T.A.; Peres, N.T.A.; Lang, E.A.S.; Gomes, E.V.; Quaresemin, N.R.; Martins, M.P.; Lopes, L.; Rossi, A. Dermatophyte resistance to antifungal drugs: Mechanisms and prospectus. Front. Microbiol. 2018, 9, 1108. [Google Scholar]
- Danielli, L.J.; Lopes, W.; Vainstein, M.H.; Fuentefria, A.M.; Apel, M.A. Biofilm formation by Microsporum canis. Clin. Microbiol. Infect. 2017, 23, 941–942. [Google Scholar]
- Merghni, A.; Haddaji, N.; Bouali, N.; Alabbosh, K.F.; Adnan, M.; Snoussi, M.; Noumi, E. Comparative study of antibacterial, antibiofilm, antiswarming and antiquorum sensing activities of Origanum vulgare essential oil and terpinene-4-ol against pathogenic bacteria. Life 2022, 12, 1616. [Google Scholar]
- Saadabi, A. Antifungal activity of some Saudi plants used in traditional medicine. Asian J. Plant Sci. 2006, 5, 907–909. [Google Scholar]
- Avijgan, M.; Hafizi, M.; Mehdi, S.; Nilforoushzadeh, M.A. Antifungal effect of Echinophora Platyloba’s extract against Candida albicans. IJPR 2006, 4, 285–289. [Google Scholar]
- Benoutman, A.; Erbiai, E.H.; Edderdaki, F.Z.; Cherif, E.K.; Saidi, R.; Lamrani, Z.; Pintado, M.; Pinto, E.; da Silva, J.C.G.E.; Maouni, A. Phytochemical composition, antioxidant and antifungal activity of Thymus capitatus, a medicinal plant collected from Northern Morocco. Antibiotics 2022, 11, 681. [Google Scholar]
- Silva, M.R.R.; Oliveira, J.G., Jr.; Fernandes, O.F.L.; Passos, X.S.; Costa, C.R.; Souza, L.K.H.; Lemos, J.A.; Paula, J.R. Antifungal activity of Ocimum gratissimum towards dermatophytes. Mycoses 2005, 48, 172–175. [Google Scholar]
- Falahati, M.; Tabrizib, N.O.; Jahaniani, F. Antidermatophyte activities of Eucalyptus camadulensis in comparison with griseofulvin. IJPT 2005, 4, 80–83. [Google Scholar]
- Arumugam, G.; Swamy, M.K.; Sinniah, U.R. Plectranthus amboinicus (Lour.) Spreng: Botanical, phytochemical, pharmacological, and nutritional significance. Molecules 2016, 21, 369. [Google Scholar]
- Lukhoba, C.W.; Simmonds, M.S.; Paton, A.J. Plectranthus: A review of ethnobotanical uses. J. Ethnopharmacol. 2006, 103, 1–24. [Google Scholar]
- Ślusarczyk, S.; Cieślak, A.; Yanza, Y.R.; Szumacher-Strabel, M.; Varadyova, Z.; Stafiniak, M.; Wojnicz, D.; Matkowski, A. Phytochemical profile and antioxidant activities of Coleus amboinicus Lour. cultivated in Indonesia and Poland. Molecules 2021, 26, 2915. [Google Scholar]
- Cheng, C.Y.; Kao, C.L.; Li, H.T.; Yeh, H.C.; Fang, Z.Y.; Li, W.J.; Wu, H.M. A new flavonoid from Plectranthus amboinicus. Chem. Nat. Compd. 2021, 57, 28–29. [Google Scholar]
- Ruan, T.Z.; Kao, C.L.; Hsieh, Y.L.; Li, H.T.; Chen, C.Y. Chemical constituents of the leaves of Plectranthus amboinicus. Chem. Nat. Compd. 2019, 55, 124–126. [Google Scholar]
- El-hawary, S.S.; El-sofany, R.H.; Abdel-Monem, A.R.; Ashour, R.S.; Sleem, A.A. Polyphenolics content and biological activity of Plectranthus amboinicus (Lour.) spreng growing in Egypt (Lamiaceae). Pharmacogn. J. 2012, 32, 45–54. [Google Scholar]
- Selvakumar, P.; Naveena, B.E.; Prakash, S.D. Studies on the antidandruff activity of the essential oil of Coleus amboinicus and Eucalyptus globulus. Asian Pacific. J. Trop. Dis. 2012, 2, S715–S719. [Google Scholar]
- Laila, F.; Fardiaz, D.; Yuliana, N.D.; Damanik, M.R.M.; Dewi, F.N.A. Methanol extract of Coleus amboinicus (Lour) exhibited antiproliferative activity and induced programmed cell death in Colon Cancer Cell WiDr. Int. J. Food Sci. 2020, 24, 9068326. [Google Scholar]
- Chiu, Y.J.; Huang, T.H.; Chiu, C.S.; Lu, T.C.; Chen, Y.W.; Peng, W.H.; Chen, C.Y. Analgesic and antiinflammatory activities of the aqueous extract from Plectranthus amboinicus (Lour.) Spreng. both in vitro and in vivo. Evid. Based Complement. Altern. Med. 2012, 2012, 508137. [Google Scholar]
- Chang, J.M.; Cheng, C.M.; Hung, L.M.; Chung, Y.S.; Wu, R.Y. Potential use of Plectranthus amboinicus in the treatment of rheumatoid arthritis. Evid. Based Complement. Altern. Med. 2010, 7, 115–120. [Google Scholar]
- Hsu, Y.C.; Cheng, C.P.; Chang, D.M. Plectranthus amboinicus attenuates inflammatory bone erosion in mice with collagen-induced arthritis by downregulation of RANKL-induced NFATc1 expression. J. Rheumatol. 2011, 38, 1844–1857. [Google Scholar]
- de Oliveira, F.F.; Torres, A.F.; Gonçalves, T.B.; Santiago, G.M.; de Carvalho, C.B.; Aguiar, M.B.; Camara, L.M.; Rabenhorst, S.H.; Martins, A.M.; Valença Junior, J.T.; et al. Efficacy of Plectranthus amboinicus (Lour.) Spreng in a murine model of methicillin-resistant Staphylococcus aureus skin abscesses. Evid. Based Complementary Altern. Med. 2013, 2013, 291592. [Google Scholar]
- Arjunan, N.; Murugan, K.; Madhiyazhagan, P.; Kovendan, K.; Prasannakumar, K.; Thangamani, S.; Barnard, D.R. Mosquitocidal and water purification properties of Cynodon dactylon, Aloe vera, Hemidesmus indicus and Coleus amboinicus leaf extracts against the mosquito vectors. Parasitol. Res. 2012, 110, 1435–1443. [Google Scholar]
- Pouët, C.; Deletre, E.; Rhino, B. Repellency of wild oregano plant volatiles, Plectranthus amboinicus, and their essential oils to the silverleaf whitefly, Bemisia Tabaci, on tomato. Neotrop. Entomol. 2022, 51, 133–142. [Google Scholar]
- Leesombun, A.; Sungpradit, S.; Boonmasawai, S.; Weluwanarak, T.; Klinsrithong, S.; Ruangsittichai, J.; Ampawong, S.; Masmeatathip, R.; Changbunjong, T. Insecticidal activity of Plectranthus amboinicus essential oil against the stable fly Stomoxys calcitrans (Diptera: Muscidae) and the horse fly Tabanus megalops (Diptera: Tabanidae). Insects 2022, 13, 255. [Google Scholar]
- Hsu, K.-P.; Ho, C.-L. Antimildew effects of Plectranthus amboinicus leaf essential oil on paper. Nat. Prod. Commun. 2019, 14, 1934578X19862903. [Google Scholar]
- Ramage, G.; Rajendran, R.; Sherry, L.; Williams, C. Fungal biofilm resistance. Int. J. Microbiol. 2012, 2012, 528521. [Google Scholar]
- Roudbary, M.; Vahedi-Shahandashti, R.; Santos, A.L.S.D.; Roudbar, M.S.; Aslani, P.; Lass-Flörl, C.; Rodrigues, C.F. Biofilm formation in clinically relevant filamentous fungi: A therapeutic challenge. Crit. Rev. Microbiol. 2022, 48, 197–221. [Google Scholar]
- Brilhante, R.S.N.; Correia, E.E.M.; Guedes, G.M.M.; de Oliveira, J.S.; Castelo-Branco, D.; Cordeiro, R.A.; Pinheiro, A.Q.; Chaves, L.J.Q.; Pereira Neto, W.A.; Sidrim, J.J.C.; et al. In vitro activity of azole derivatives and griseofulvin against planktonic and biofilm growth of clinical isolates of dermatophytes. Mycoses 2018, 61, 449–454. [Google Scholar]
- Wu, S.; Wang, Y.; Liu, N.; Dong, G.; Sheng, C. Tackling fungal resistance by biofilm inhibitors. J. Med. Chem. 2017, 23, 2193–2211. [Google Scholar]
- Baldo, A.; Monod, M.; Mathy, A.; Cambier, L.; Bagut, E.T.; Defaweux, V.; Symoen, S.F.; Antoine, N.; Mignon, B. Mechanisms of skin adherence and invasion by dermatophytes. Mycoses 2012, 55, 218–223. [Google Scholar]
- Brilhante, R.S.N.; Aguiar, L.; Sales, J.A.; Araújo, G.D.S.; Pereira, V.S.; Pereira-Neto, W.A.; Pinheiro, A.Q.; Paixão, G.C.; Cordeiro, R.A.; Sidrim, J.J.C.; et al. Ex Vivo biofilm-forming ability of dermatophytes using dog and cat hair: An ethically viable approach for an infection model. Biofouling 2019, 35, 392–400. [Google Scholar]
- Al-Fattani, M.A.; Douglas, L.J. Penetration of Candida biofilms by antifungal agents. Antimicrob. Agents Chemother. 2004, 48, 3291–3297. [Google Scholar]
- Shi, C.; Liu, J.; Li, W.; Zhao, Y.; Meng, L.; Xiang, M. Expression of fluconazole resistance-associated genes in biofilm from 23 clinical isolates of Candida albicans. Braz. J. Microbiol. 2019, 50, 157–163. [Google Scholar]
- Jabra-Rizk, M.A.; Falkler, W.A.; Meiller, T.F. Fungal biofilms and drug resistance. Emerg. Infect. Dis. 2004, 10, 14–19. [Google Scholar]
- Bila, N.M.; Costa-Orlandi, C.B.; Vaso, C.O.; Bonatti, J.L.C.; de Assis, L.R.; Regasini, L.O.; Fontana, C.R.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J.S. 2-Hydroxychalcone as a potent compound and photosensitizer against dermatophyte biofilms. Front. Cell Infect. Microbiol. 2021, 11, 679470. [Google Scholar]
- Girish, K. Antimicrobial activities of Coleus aromaticus Benth. J. Pharm. Res. 2016, 10, 635–646. [Google Scholar]
- Umayal, S.; Geetha, R.V. Comparative evaluation of antibiofilm formation activity of Plectranthus amboinicus extract against Streptococcus mutans. Drug Invent. Today 2019, 12, 2253–2255. [Google Scholar]
- Vasconcelos, S.E.C.B.; Melo, H.M.; Cavalcante, T.T.A.; Júnior, F.E.A.C.; de Carvalho, M.G.; Menezes, F.G.R.; de Sousa, O.V.; Costa, R.A. Plectranthus amboinicus essential oil and carvacrol bioactive against planktonic and biofilm of oxacillin- and vancomycin-resistant Staphylococcus aureus. BMC Complement. Altern. Med. 2017, 17, 462. [Google Scholar]
- Manimekalai, K.; Srinivasan, P.; Dineshbabu, J.; Guna, G.; Darsini, D. Anti-biofilm efficacy of Plectranthus amboinicus against Streptococcus pyogenes isolated from pharyngitis patients. Asian J. Pharm. Clin. Res. 2016, 9, 348–354. [Google Scholar]
- Wadikar, D.D.; Patki, P.E. Coleus aromaticus: A therapeutic herb with multiple potentials. J. Food Sci. Technol. 2016, 53, 2895–2901. [Google Scholar]
- da Costa, J.G.M.; Pereira, C.K.B.; Rodrigues, F.F.G.; de Lima, S.G. Chemical composition, antibacterial and fungicidal activities of leaf oil of Plectranthus amboinicus (Lour.) Spreng. JEOR 2010, 22, 183–185. [Google Scholar]
- Dhifi, W.; Bellili, S.; Jazi, S.; Bahloul, N.; Mnif, W. Essential oils’ chemical characterization and investigation of some biological activities: A critical review. Medicines 2016, 3, 25. [Google Scholar]
- Wang, F.; Wei, F.; Song, C.; Jiang, B.; Tian, S.; Yi, J.; Yu, C.; Song, Z.; Sun, L.; Bao, Y.; et al. Dodartia orientalis L. essential oil exerts antibacterial activity by mechanisms of disrupting cell structure and resisting biofilm. Ind. Crops Prod. 2017, 109, 358–366. [Google Scholar]
- Kordali, S.; Cakir, A.; Ozer, H.; Cakmakci, R.; Kesdek, M.; Mete, E. Antifungal, phytotoxic and insecticidal properties of essential oil isolated from Turkish Origanum acutidens and its three components, carvacrol, thymol and p-cymene. Bioresour. Technol. 2008, 99, 8788–8795. [Google Scholar]
- Rao, A.; Zhang, Y.; Muend, S.; Rao, R. Mechanism of antifungal activity of terpenoid phenols resembles calcium stress and inhibition of the TOR pathway. Antimicrob. Agents Chemother. 2010, 54, 5062–5069. [Google Scholar]
- Zuzarte, M.; Vale-Silva, L.; Gonçalves, M.J.; Cavaleiro, C.; Vaz, S.; Canhoto, J.; Pinto, E.; Salgueiro, L. Antifungal activity of phenolic-rich Lavandula multifida L. essential oil. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 1359–1366. [Google Scholar]
- Abbaszadeh, S.; Sharifzadeh, A.; Shokri, H.; Khosravi, A.R.; Abbaszadeh, A. Antifungal efficacy of thymol, carvacrol, eugenol and menthol as alternative agents to control the growth of food-relevant fungi. J. Mycol. Med. 2014, 24, e51–e56. [Google Scholar]
- Tian, F.; Woo, S.Y.; Lee, S.Y.; Chun, H.S. p-Cymene and its derivatives exhibit antiaflatoxigenic activities against Aspergillus flavus through multiple modes of action. Appl. Biol. Chem. 2018, 61, 489–497. [Google Scholar]
- Niu, C.; Wang, C.; Yang, Y.; Chen, R.; Zhang, J.; Chen, H.; Zhuge, Y.; Li, J.; Cheng, J.; Xu, K.; et al. Carvacrol induces Candida albicans apoptosis associated with Ca2+/calcineurin pathway. Front. Cell. Infect. Microbiol. 2020, 10, 192. [Google Scholar]
- Alves Resende, J.; Soares Toneto, D.; Cruz Albuquerque, M.C.; Areas Bastos, K.; Fontes Pinheiro, P.; Drummond Costa Ignacchiti, M. Antibacterial and anti-biofilm potential of Plectranthus amboinicus (Lour.) Spreng essential oil and carvacrol against Staphylococcus aureus and Escherichia coli. Rev. Ciênc. Méd. Biol. 2022, 21, 11–17. [Google Scholar]
- Marchese, A.; Arciola, C.R.; Barbieri, R.; Silva, A.S.; Nabavi, S.F.; Tsetegho Sokeng, A.J.; Morteza Izadi, M.; Jonaidi Jafari, N.; Suntar, I.; Daglia, M.; et al. Update on monoterpenes as antimicrobial agents: A particular focus on p-Cymene. Materials 2017, 10, 947. [Google Scholar]
- Couto, C.; Raposo, N.; Rozental, S.; Borba-Santos, L.; Bezerra, L.; Almeida, P.; Brandão, M. Chemical composition and antifungal properties of essential oil of Origanum vulgare Linnaeus (Lamiaceae) against Sporothrix schenckii and Sporothrix brasiliensis. Trop. J. Pharm. Res. 2015, 14, 1207–1212. [Google Scholar]
- Baba, S.A.; Malik, S.A. Determination of total phenolic and flavonoid content, antimicrobial and antioxidant activity of a root extract of Arisaema jacquemontii Blume. J. Taibah Univ. Med. Sci. 2015, 9, 449–454. [Google Scholar]
- Nguyen, N.Q.; Minh, L.V.; Trieu, L.H.; Bui, L.M.; Lam, T.D.; Hieu, V.Q.; Khang, T.V.; Trung, L.N.Y. Evaluation of total polyphenol content, total flavonoid content, and antioxidant activity of Plectranthus amboinicus leaves. IOP Conf. Ser. Mater. Sci. Eng. 2020, 736, 062017. [Google Scholar]
- Bhatt, P.; Joseph, G.S.; Negi, P.S.; Varadaraj, M.C. Chemical composition and nutraceutical potential of Indian Borage (Plectranthus amboinicus) stem extract. J. Chem. 2013, 2013, 320329. [Google Scholar]
- Harborne, J.B.; Williams, C.A. Advances in flavonoid research since 1992. Phytochemistry 2000, 55, 481–504. [Google Scholar]
- Cushnie, T.P.T.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356. [Google Scholar]
- Ahmad, H.; Matsubara, Y.I. Effect of lemon balm water extract on Fusarium Wilt control in strawberry and antifungal properties of secondary metabolites. Hort. J. 2020, 89, 175–181. [Google Scholar]
- Swari, D.A.M.A.; Santika, I.W.M.; Aman, I.G.M. Antifungal activities of ethanolic extract of rosemary leaf (Rosemarinus officinalis L.) against Candida albicans. JPSA 2020, 2, 28–35. [Google Scholar]
- Aboody, M.S.A.; Mickymaray, S. Anti-fungal efficacy and mechanisms of flavonoids. Antibiotics 2020, 9, 45. [Google Scholar]
- Lee, H.; Woo, E.R.; Lee, D.G. Apigenin induces cell shrinkage in Candida albicans by membrane perturbation. FEMS Yeast Res. 2018, 18, foy003. [Google Scholar]
- Singh, G.; Kumar, P.; Joshi, S.C. Treatment of dermatophytosis by a new antifungal agent ‘apigenin’. Mycoses 2014, 57, 497–506. [Google Scholar]
- Alfarrayeh, I.; Pollák, E.; Czéh, Á.; Vida, A.; Das, S.; Papp, G. Antifungal and anti-biofilm effects of caffeic acid phenethyl ester on different Candida Species. Antibiotics 2021, 10, 1359. [Google Scholar]
- Sun, L.; Liao, K.; Hang, C. Caffeic acid phenethyl ester synergistically enhances the antifungal activity of fluconazole against resistant Candida albicans. Phytomedicine 2018, 40, 55–58. [Google Scholar]
- Cantelli, B.A.M.; Bitencourt, T.A.; Komoto, T.T.; Beleboni, R.O.; Marins, M.; Fachin, A.L. Caffeic acid and licochalcone A interfere with the glyoxylate cycle of Trichophyton rubrum. Biomed. Pharmacother. 2017, 96, 1389–1394. [Google Scholar]
- Wolfe, K.; Wu, X.; Liu, R.H. Antioxidant activity of apple peels. J. Agric. Food Chem. 2003, 51, 609–614. [Google Scholar]
- Matić, P.; Sabljić, M.; Jakobek, L. Validation of spectrophotometric methods for the determination of total polyphenol and total flavonoid content. J. AOAC Int. 2019, 100, 1795–1803. [Google Scholar]
- Petrucelli, M.F.; de Abreu, M.H.; Cantelli, B.A.M.; Segura, G.G.; Nishimura, F.G.; Bitencourt, T.A.; Marins, M.; Fachin, A.L. Epidemiology and diagnostic perspectives of dermatophytoses. J. Fungi 2020, 6, 310. [Google Scholar]
- Sugita, T.; Nishikawa, A.; Shinoda, T. Identification of Trichosporon asahii by PCR based on sequences of the internal transcribed spacer regions. J. Clin. Microbiol. 1998, 36, 2742–2744. [Google Scholar]
- Turin, L.; Riva, F.; Galbiati, G.; Cainelli, T. Fast, simple and highly sensitive double-rounded polymerase chain reaction assay to detect medically relevant fungi in dermatological specimens. Eur. J. Clin. Investig. 2000, 30, 511–518. [Google Scholar]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.fr: Robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 2008, 1, 465–469. [Google Scholar]
- Sen, S.; Borah, S.N.; Bora, A.; Deka, S. Rhamnolipid exhibits anti-biofilm activity against the dermatophytic fungi Trichophyton rubrum and Trichophyton mentagrophytes. Biotechnol. Rep. 2020, 27, e00516. [Google Scholar]
- Sales, G.; Medeiros, S.; Soares, I.; Sampaio, T.; Bandeira, M.; Nogueira, N.; Queiroz, M. Antifungal and modulatory activity of Lemon Balm (Lippia alba (MILL.) N. E. BROWN) essential oil. Sci. Pharm. 2022, 90, 31. [Google Scholar]
- M38-A2; Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi. Clinical and Laboratory Standards Institute (CLSI): Wayne, PA, USA, 2008.
- Kim, D.J.; Lee, M.W.; Choi, J.S.; Lee, S.G.; Park, J.Y.; Kim, S.W. Inhibitory activity of hinokitiol against biofilm formation in fluconazole-resistant Candida species. PLoS ONE 2017, 12, e0171244. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Retention Time (min) | Classes | Compounds | Formula | Chemical Structure | Peak Area (%) | Similarity Index (%) |
|---|---|---|---|---|---|---|---|
| 1 | 3.10 | Aldehydes | Hexenal | C6H10O |  | 0.17 | 94 |
| 2 | 4.02 | Monoterpene | α-Thujene | C10H16 |  | 0.71 | 94 |
| 3 | 4.78 | Monoterpene | Sabinene | C10H16 |  | 0.12 | 91 |
| 4 | 5.09 | Monoterpene | β-Myrcene | C10H16 |  | 2.57 | 97 |
| 5 | 5.44 | Monoterpene | α-Terpinene | C10H16 |  | 1.62 | 98 |
| 6 | 5.59 | Monoterpene | p-Cymene | C10H14 |  | 10.89 | 97 |
| 7 | 6.04 | Monoterpene | γ-Terpinene | C10H16 |  | 9.33 | 97 |
| 8 | 7.58 | Monoterpene | 4-Terpineol | C10H18O |  | 1.26 | 96 |
| 9 | 9.32 | Monoterpene | Thymol | C10H14O |  | 0.17 | 90 |
| 10 | 9.44 | Monoterpene | Carvacrol | C10H14O |  | 56.65 | 97 |
| 11 | 10.59 | Sesquiterpene | β-Caryophyllene | C15H24 |  | 7.12 | 99 |
| 12 | 10.85 | Sesquiterpene | α-Farnesene | C15H24 |  | 5.41 | 91 |
| 13 | 11.23 | Sesquiterpene | α-Humulene | C15H24 |  | 2.28 | 97 |
| 14 | 11.82 | Sesquiterpene | β-Farnesene | C15H24 |  | 0.19 | 92 |
| 15 | 12.38 | Sesquiterpene | β-Bisabolene | C15H24 |  | 0.26 | 95 |
| 16 | 12.76 | Sesquiterpene | β-Sesquiphellandrene | C15H24 |  | 0.11 | 95 |
| 17 | 14.25 | Sesquiterpene | (-)-Caryophyllene oxide | C15H24O |  | 0.69 | 74 |
| 18 | 24.51 | Phthalates | Bis(2-ethylhexyl) phthalate | C24H38O4 |  | 0.29 | 91 |
| Total | 99.84 |
| No. | Retention Time (min) | Compounds | Peak Area | Concentration (mg/g Sample) |
|---|---|---|---|---|
| 1 | 1.52 | Caffeic acid | 96.78 | 0.73 |
| 2 | 3.70 | Rosmarinic acid | 67.43 | 1.25 |
| 3 | 4.55 | Apigenin | 105.93 | 1.17 |
| No. | Fungi | Biofilm Classification | MIC (µg/mL) | ||
|---|---|---|---|---|---|
| Fluconazole | CEO | CEE | |||
| 1 | Candida albicans | strong | 8 | 128 | 64 |
| 2 | Trichophyton rubrum | moderate | 4 | 32 | 256 |
| 3 | W01 | weak | 8 | 32 | 128 |
| 4 | W02 | weak | 8 | 128 | 128 |
| 5 | W03 | weak | 16 | 128 | 128 |
| 6 | M01 | moderate | 4 | 32 | 128 |
| 7 | M02 | moderate | 4 | 256 | 128 |
| 8 | M03 | moderate | 8 | 64 | 128 |
| 9 | M04 | moderate | 8 | 128 | 128 |
| 10 | M05 | moderate | 8 | 128 | 256 |
| 11 | M06 | moderate | 16 | 128 | 32 |
| 12 | S01 | strong | 4 | 64 | 256 |
| 13 | S02 | strong | 4 | 64 | 256 |
| 14 | S03 | strong | 8 | 128 | 64 |
| Treatment | Minimum Inhibitory Concentration (MIC) (µg/mL) | |||
|---|---|---|---|---|
| MIC50 | MIC90 | GM of MIC | MIC Range | |
| Fluconazole | 8 | 16 | 8 | 4–16 |
| CEO | 128 | 128 | 90.51 | 32–256 |
| CEE | 128 | 256 | 128 | 32–256 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leesombun, A.; Thanapakdeechaikul, K.; Suwannawiang, J.; Mukto, P.; Sungpradit, S.; Bangphoomi, N.; Changbunjong, T.; Thongjuy, O.; Weluwanarak, T.; Boonmasawai, S. Effects of Coleus amboinicus L. Essential Oil and Ethanolic Extracts on Planktonic Cells and Biofilm Formation of Microsporum canis Isolated from Feline Dermatophytosis. Antibiotics 2022, 11, 1734. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics11121734
Leesombun A, Thanapakdeechaikul K, Suwannawiang J, Mukto P, Sungpradit S, Bangphoomi N, Changbunjong T, Thongjuy O, Weluwanarak T, Boonmasawai S. Effects of Coleus amboinicus L. Essential Oil and Ethanolic Extracts on Planktonic Cells and Biofilm Formation of Microsporum canis Isolated from Feline Dermatophytosis. Antibiotics. 2022; 11(12):1734. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics11121734
Chicago/Turabian StyleLeesombun, Arpron, Karnchanarat Thanapakdeechaikul, Jiraporn Suwannawiang, Pipada Mukto, Sivapong Sungpradit, Norasuthi Bangphoomi, Tanasak Changbunjong, Orathai Thongjuy, Thekhawet Weluwanarak, and Sookruetai Boonmasawai. 2022. "Effects of Coleus amboinicus L. Essential Oil and Ethanolic Extracts on Planktonic Cells and Biofilm Formation of Microsporum canis Isolated from Feline Dermatophytosis" Antibiotics 11, no. 12: 1734. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics11121734