Clonal Structure, Virulence Factor-encoding Genes and Antibiotic Resistance of Escherichia coli, Causing Urinary Tract Infections and Other Extraintestinal Infections in Humans in Spain and France during 2016

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Phylogenetic Groups
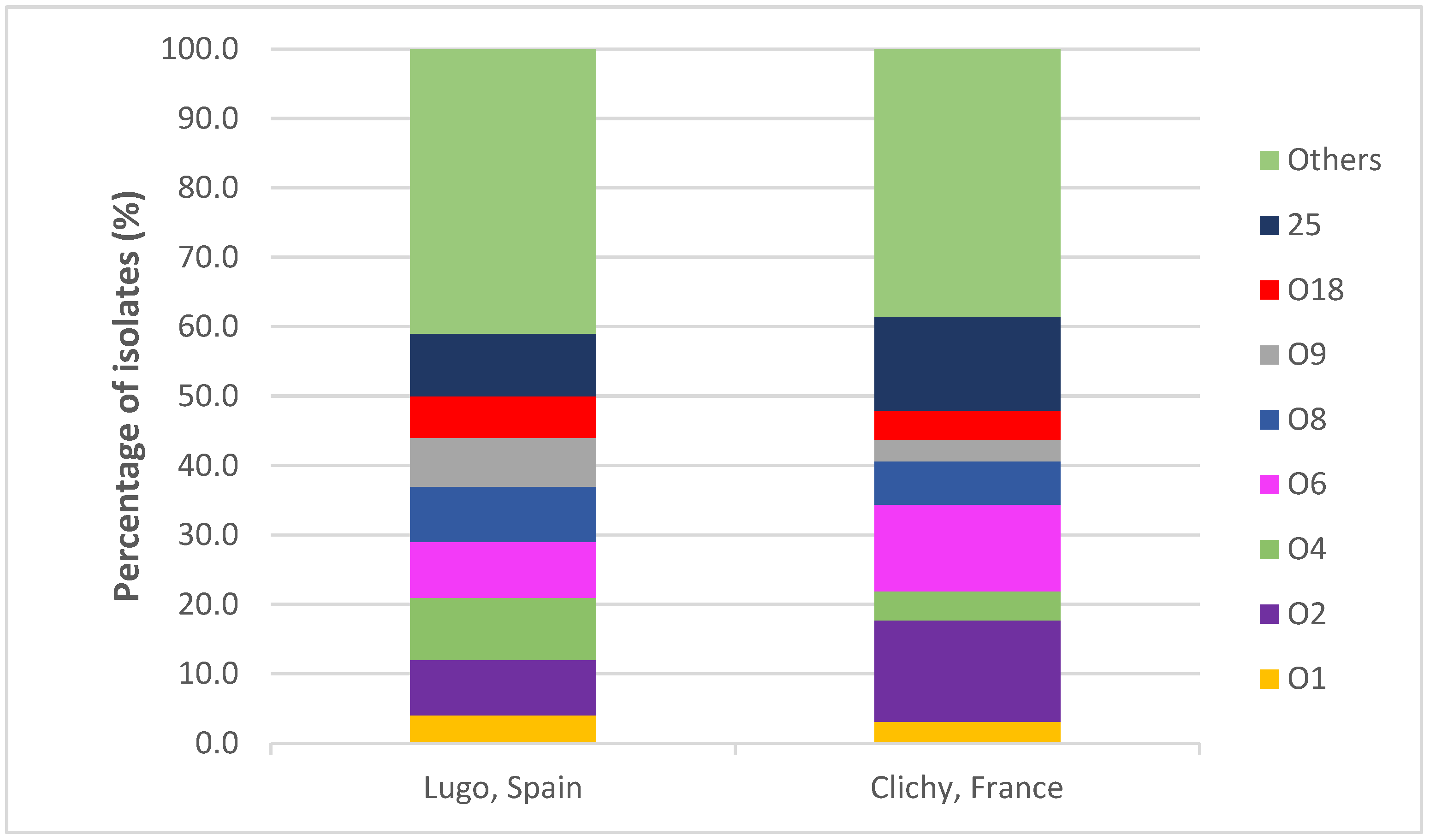
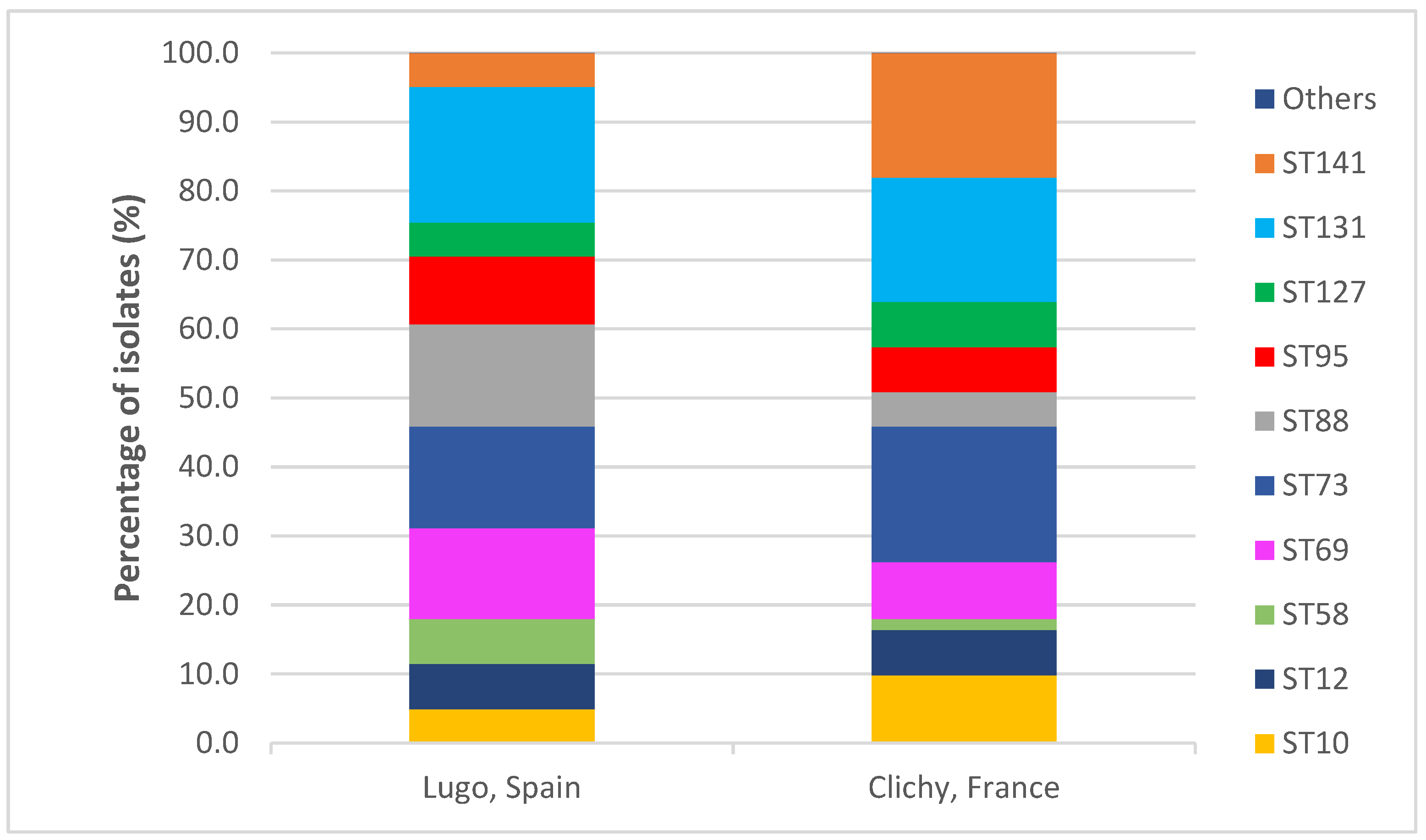
2.2. Serotypes and Sequence Types
2.3. Clones
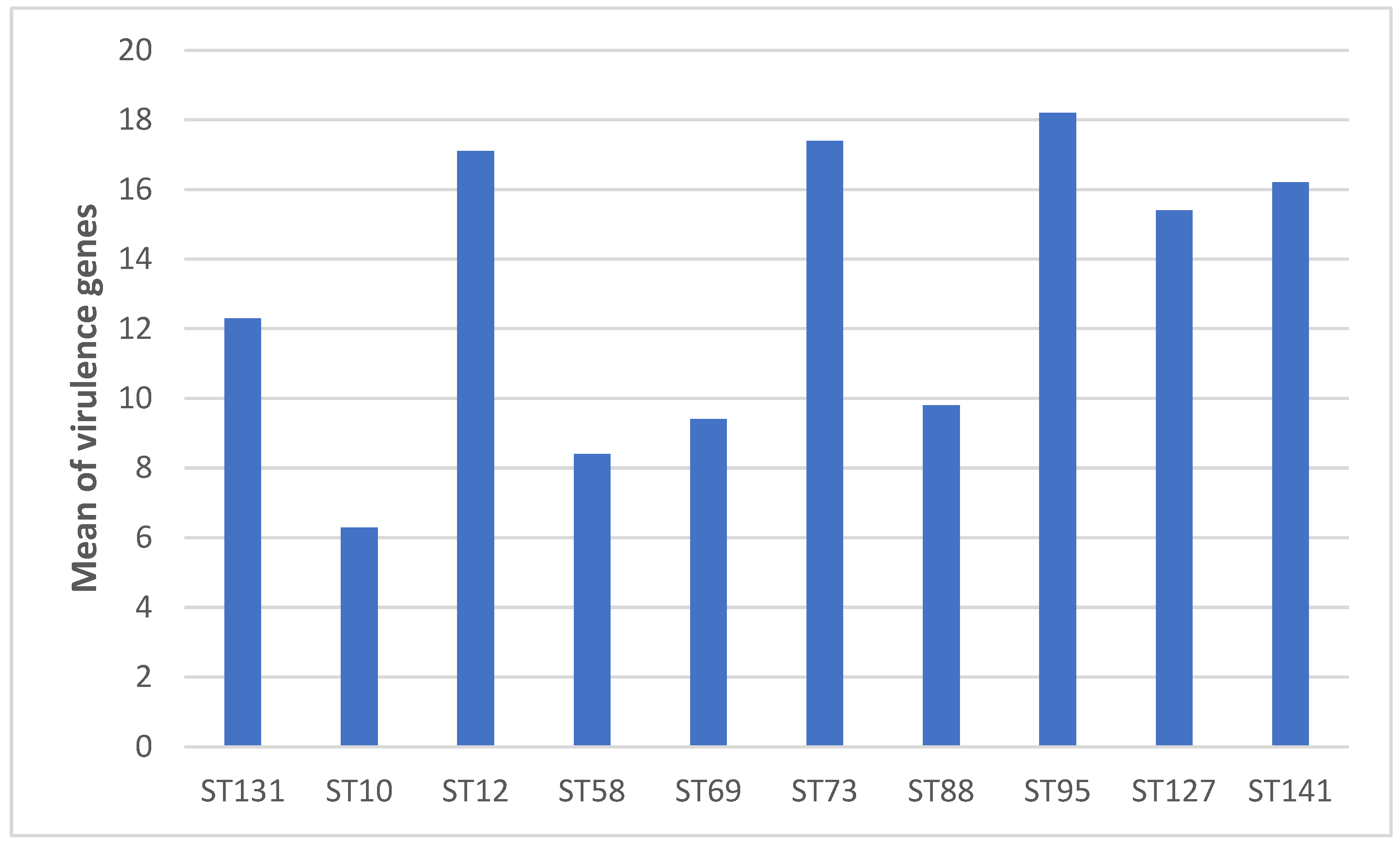
2.4. Virulence Factor (VF)-Encoding Genes
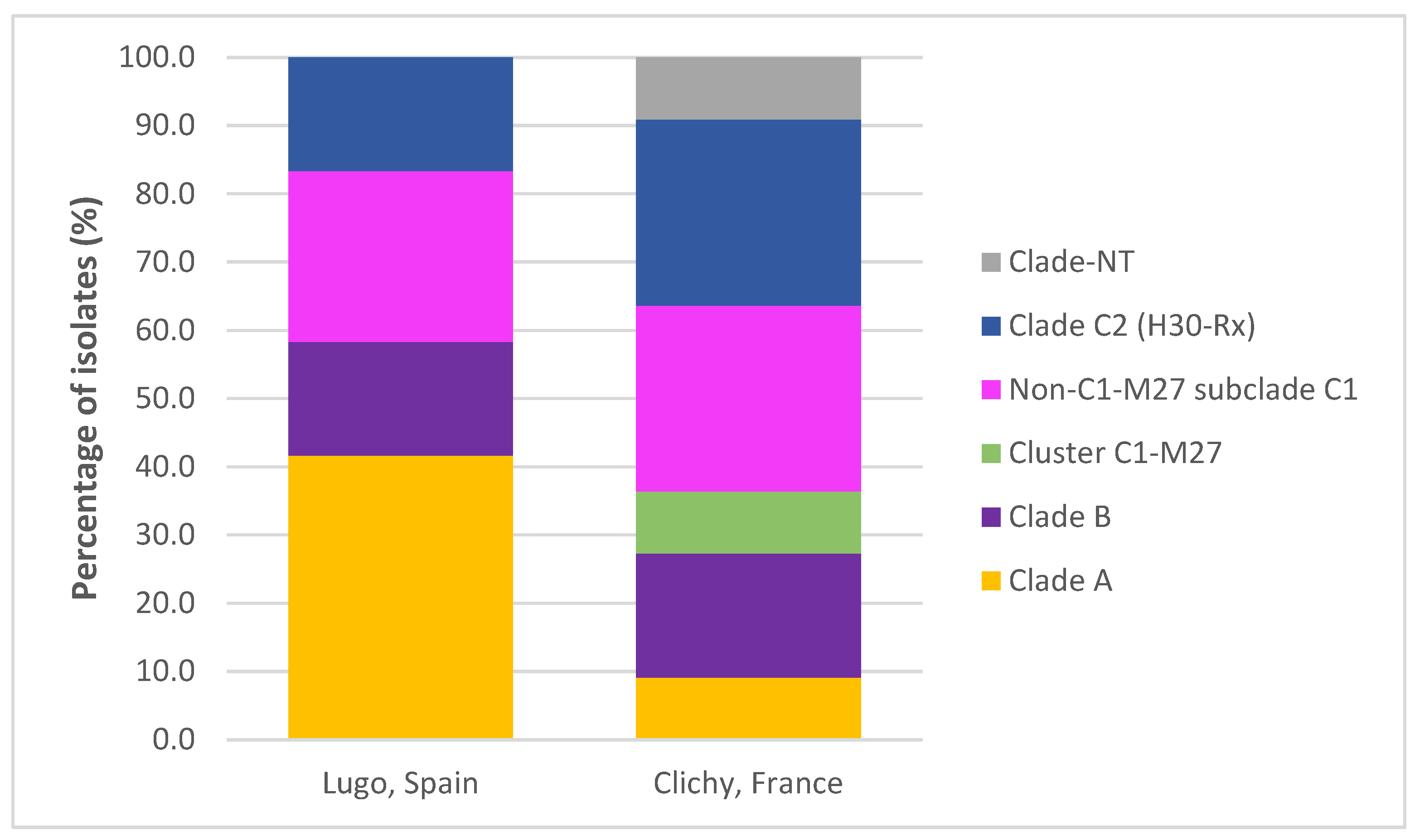
2.5. Clonotypes, Clades, Subclades, Clusters and virotypes of ST131 Isolates
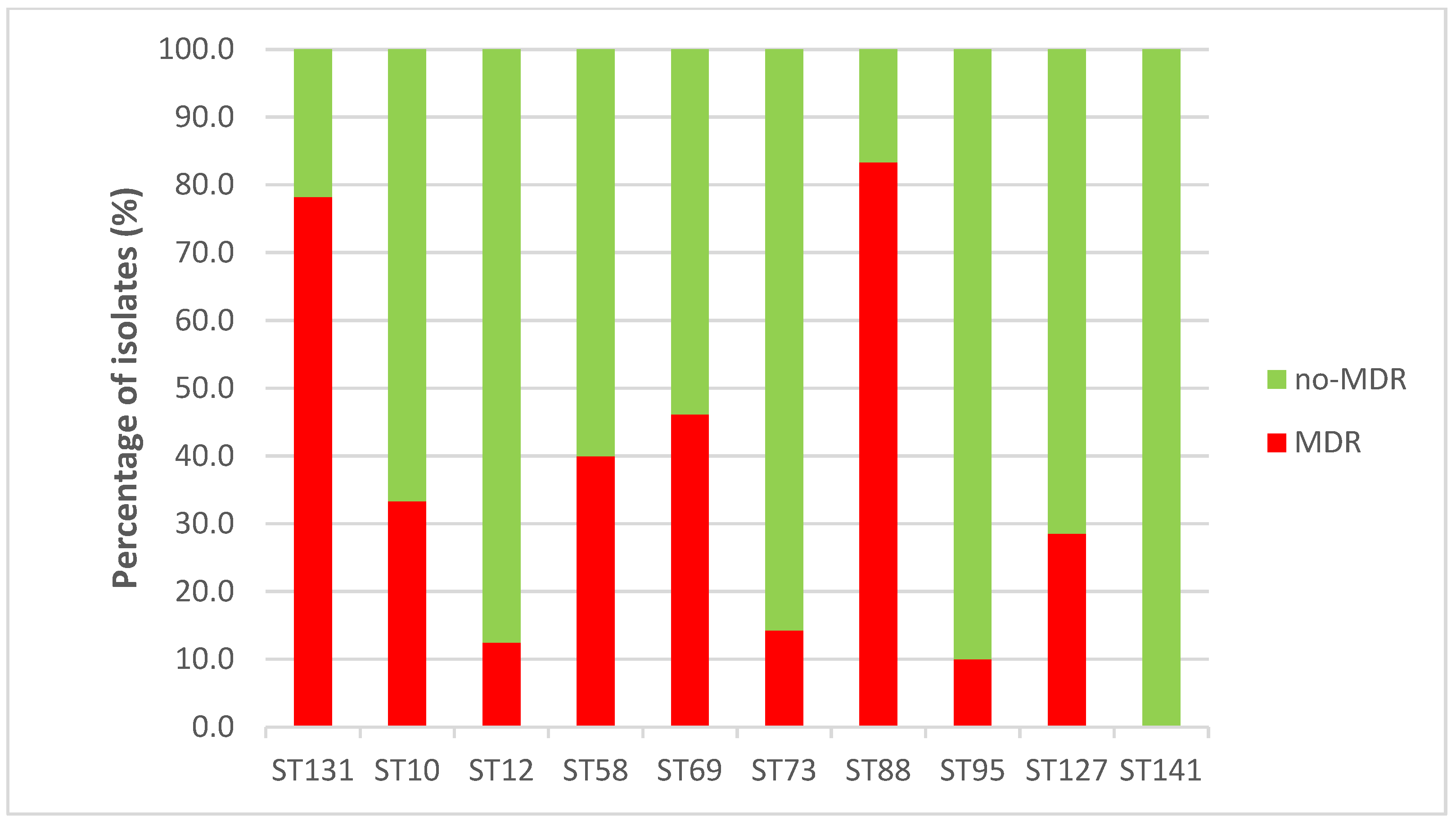
2.6. Antimicrobial Resistance
2.7. Hybrid Pathotypes
3. Discussion
4. Materials and Methods
4.1. E. coli isolates
4.2. Phylogenetic Grouping, Serotyping, MLST, CH Typing and Identification of ST131 Clades and Subclades
- Assignment to the main phylogroups (A, B1, B2, C, D, E, F) was based on the protocol of Clermont et al. [37]
- The determination of O and H antigens was carried out using the method previously described by Guinée et al. [38], with all available O (O1 to O181) and H (H1 to H56) antisera. Isolates that did not react with any antisera were classified as O non-typeable (ONT) or HNT and those that were non-motile were denoted as HNM.
- The sequence types (STs) were established following the multilocus sequence typing (MLST) scheme of Achtman by gene amplification and sequencing of the seven housekeeping genes (adk, fumC, gyrB, icd, mdh, purA, and recA), according to the protocol and primers specified at the E. coli MLST web site (http://mlst.warwick.ac.uk/mlst/dbs/Ecoli) [39].
- The ST131 clades (A, B, C), subclade C2 (also known as subclone H30Rx) and subclade C1 and cluster C1-M27 were established by PCR [41].
4.3. Virulence Genotyping
4.4. Antimicrobial Susceptibility and ESBL Typing
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Spurbeck, R.R.; Dinh, P.C.; Walk, S.T.; Stapleton, A.E.; Hooton, T.M.; Nolan, L.K.; Kim, K.S.; Johnson, J.R.; Mobley, H.L.T. Escherichia coli isolates that carry vat, fyuA, chuA, and yfcV efficiently colonize the urinary tract. Infect. Immun. 2012, 80, 4115–4122. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.R.; Porter, S.; Johnston, B.; Kuskowski, M.A.; Spurbeck, R.R.; Mobley, H.L.T.; Williamson, D.A. Host characteristics and bacterial traits predict experimental virulence for Escherichia coli bloodstream isolates from patients with urosepsis. Open Forum Infect. Dis. 2015, 2, 83. [Google Scholar] [CrossRef]
- Nicolas-Chanoine, M.H.; Blanco, J.; Leflon-Guibout, V.; Demarty, R.; Alonso, M.P.; Caniça, M.M.; Park, Y.J.; Lavigne, J.P.; Pitout, J.; Johnson, J.R. Intercontinental emergence of Escherichia coli clone O25:H4-ST131 producing CTX-M-15. J. Antimicrob. Chemother. 2008, 61, 273–281. [Google Scholar] [CrossRef] [Green Version]
- De Toro, M.; Fernández, J.; García, V.; Mora, A.; Blanco, J.; de la Cruz, F.; Rodicio, M.R. Whole genome sequencing, molecular typing and in vivo virulence of OXA-48-producing Escherichia coli isolates including ST131 H30-Rx, H22 and H41 subclones. Sci. Rep. 2017, 7, 12103. [Google Scholar] [CrossRef] [Green Version]
- Mamani, R.; Flament-Simon, S.C.; García, V.; Mora, A.; Alonso, M.P.; López, C.; García-Meniño, I.; Díaz-Jiménez, D.; Blanco, J.E.; Blanco, M.; et al. Sequence types, clonotypes, serotypes, and virotypes of extended-spectrum β-lactamase-producing Escherichia coli causing bacteraemia in a Spanish hospital over a 12-year period (2000 to 2011). Front. Microbiol. 2019, 10, 1530. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.R.; Orskov, I.; Orskov, F.; Goullet, P.; Picard, B.; Moseley, S.L.; Roberts, P.L.; Stamm, W.E. O, K, and H antigens predict virulence factors, carboxylesterase B pattern, antimicrobial resistance, and host compromise among Escherichia coli strains causing urosepsis. J. Infect. Dis. 1994, 169, 119–126. [Google Scholar] [CrossRef]
- Riley, L.W. Pandemic lineages of extraintestinal pathogenic Escherichia coli. Clin. Microbiol. Infect. 2014, 20, 380–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manges, A.R.; Geum, H.M.; Guo, A.; Edens, T.J.; Fibke, C.D.; Pitout, J.D.D. Global extraintestinal pathogenic Escherichia coli (ExPEC) lineages. Clin. Microbiol. Rev. 2019, 32, e00135-18. [Google Scholar] [CrossRef] [PubMed]
- Tchesnokova, V.L.; Rechkina, E.; Larson, L.; Ferrier, K.; Weaver, J.L.; Schroeder, D.W.; She, R.; Butler-Wu, S.M.; Aguero-Rosenfeld, M.E.; Zerr, D.; et al. Rapid and extensive expansion in the United States of a new multidrug-resistant Escherichia coli clonal group, sequence type 1193. Clin. Infect. Dis. 2019, 68, 334–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.R.; Johnston, B.D.; Porter, S.B.; Clabots, C.; Bender, T.L.; Thuras, P.; Trott, D.J.; Cobbold, R.; Mollinger, J.; Ferrieri, P.; et al. Rapid emergence, subsidence, and molecular detection of Escherichia coli sequence type 1193- FimH64, a new disseminated multidrug-resistant commensal and extraintestinal pathogen. J. Clin. Microbiol. 2019, 57, e01664-18. [Google Scholar] [CrossRef] [Green Version]
- Blanco, J.; Mora, A.; Mamani, R.; Lopez, C.; Blanco, M.; Dahbi, G.; Herrera, A.; Blanco, J.E.; Alonso, M.P.; Garcia-Garrote, F.; et al. National survey of Escherichia coli causing extraintestinal infections reveals the spread of drug-resistant clonal groups O25b:H4-B2-ST131, O15:H1-D-ST393 and CGA-D-ST69 with high virulence gene content in Spain. J. Antimicrob. Chemother. 2011, 66, 2011–2021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oteo, J.; González-López, J.J.; Ortega, A.; Quintero-Zárate, J.N.; Bou, G.; Cercenado, E.; Conejo, M.C.; Martínez-Martínez, L.; Navarro, F.; Oliver, A.; et al. Inhibitor-resistant TEM- and OXA-1-producing Escherichia coli isolates resistant to amoxicillin-clavulanate are more clonal and possess lower virulence gene content than susceptible clinical isolates. Antimicrob. Agents Chemother. 2014, 58, 3874–3881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Mee-Marquet, N.L.; Blanc, D.S.; Gbaguidi-Haore, H.; Dos Santos Borges, S.; Viboud, Q.; Bertrand, X.; Quentin, R. Marked increase in incidence for bloodstream infections due to Escherichia coli, a side effect of previous antibiotic therapy in the elderly. Front. Microbiol. 2015, 6, 646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clermont, O.; Couffignal, C.; Blanco, J.; Mentré, F.; Picard, B.; Denamur, E. Two levels of specialization in bacteraemic Escherichia coli strains revealed by their comparison with commensal strains. Epidemiol. Infect. 2017, 145, 872–882. [Google Scholar] [CrossRef] [Green Version]
- La Combe, B.; Clermont, O.; Messika, J.; Eveillard, M.; Kouatchet, A.; Lasocki, S.; Corvec, S.; Lakhal, K.; Billard-Pomares, T.; Fernandes, R.; et al. Pneumonia-specific Escherichia coli with distinct phylogenetic and virulence profiles, France, 2012–2014. Emerg. Infect. Dis. 2019, 25, 710–718. [Google Scholar] [CrossRef] [Green Version]
- Brisse, S.; Diancourt, L.; Laouenan, C.; Vigan, M.; Caro, V.; Arlet, G.; Drieux, L.; Leflon-Guibout, V.; Mentre, F.; Jarlier, V.; et al. Phylogenetic distribution of CTX-M- and non-extended-spectrum-β-lactamase-producing Escherichia coli isolates: Group B2 isolates, except clone ST131, rarely produce CTX-M enzymes. J. Clin. Microbiol. 2012, 50, 2974–2981. [Google Scholar] [CrossRef] [Green Version]
- Gati, N.S.; Middendorf-Bauchart, B.; Bletz, S.; Dobrindt, U.; Mellmann, A. Origin and evolution of hybrid Shiga toxin-producing and uropathogenic Escherichia coli strains of sequence type 141. J. Clin. Microbiol. 2019, 58, e01309-19. [Google Scholar] [CrossRef]
- Gibreel, T.M.; Dodgson, A.R.; Cheesbrough, J.; Fox, A.J.; Bolton, F.J.; Upton, M. Population structure, virulence potential and antibiotic ausceptibility of uropathogenic Escherichia coli from Northwest England. J. Antimicrob. Chemother. 2012, 67, 346–356. [Google Scholar] [CrossRef] [Green Version]
- Horner, C.; Fawley, W.; Morris, K.; Parnell, P.; Denton, M.; Wilcox, M. Escherichia coli bacteraemia: 2 years of prospective regional surveillance (2010-12). J. Antimicrob. Chemother. 2014, 69, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Day, M.J.; Doumith, M.; Abernethy, J.; Hope, R.; Reynolds, R.; Wain, J.; Livermore, D.M.; Woodford, N. Population structure of Escherichia coli causing bacteraemia in the UK and Ireland between 2001 and 2010. J. Antimicrob. Chemother. 2016, 71, 2139–2142. [Google Scholar] [CrossRef] [Green Version]
- Kallonen, T.; Brodrick, H.J.; Harris, S.R.; Corander, J.; Brown, N.M.; Martin, V.; Peacock, S.J.; Parkhill, J. Systematic longitudinal survey of invasive Escherichia coli in England demonstrates a stable population structure only transiently disturbed by the emergence of ST131. Genome Res. 2017, 27, 1437–1449. [Google Scholar] [CrossRef] [Green Version]
- Toval, F.; Kohler, C.-D.; Vogel, U.; Wagenlehner, F.; Mellmann, A.; Fruth, A.; Schmidt, M.A.; Karch, H.; Bielaszewska, M.; Dobrindt, U.; et al. Characterization of Escherichia coli isolates from hospital inpatients or outpatients with urinary tract infection. J. Clin. Microbiol. 2014, 52, 407–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nüesch-Inderbinen, M.T.; Baschera, M.; Zurfluh, K.; Hächler, H.; Nüesch, H.; Stephan, R. Clonal diversity, virulence potential and antimicrobial resistance of Escherichia coli causing community acquired urinary tract infection in Switzerland. Front. Microbiol. 2017, 8, 2334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fibke, C.D.; Croxen, M.A.; Geum, H.M.; Glass, M.; Wong, E.; Avery, B.P.; Daignault, D.; Mulvey, M.R.; Reid-Smith, R.J.; Parmley, E.J.; et al. Genomic epidemiology of major extraintestinal pathogenic Escherichia coli lineages causing urinary tract infections in young women across Canada. Open Forum Infect. Dis. 2019, 6. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, R.; Johnston, B.; Lohse, C.; Chattopadhyay, S.; Tchesnokova, V.; Sokurenko, E.V.; Johnson, J.R. The clonal distribution and diversity of extraintestinal Escherichia coli isolates vary according to patient characteristics. Antimicrob. Agents Chemother. 2013, 57, 5912–5917. [Google Scholar] [CrossRef] [Green Version]
- Yamaji, R.; Rubin, J.; Thys, E.; Friedman, C.R.; Riley, L.W. Persistent pandemic lineages of uropathogenic Escherichia coli in a college community from 1999 to 2017. J. Clin. Microbiol. 2018, 56, e01834-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams-Sapper, S.; Diep, B.A.; Perdreau-Remington, F.; Riley, L.W. Clonal composition and community clustering of drug-susceptible and -resistant Escherichia coli isolates from bloodstream infections. Antimicrob. Agents Chemother. 2013, 57, 490–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, B.K.; Ilikj, M.; McCloskey, C.B.; Chavez-Bueno, S. Antibiotic resistance and molecular characterization of bacteremia Escherichia coli isolates from newborns in the United States. PLoS ONE 2019, 14, e0219352. [Google Scholar] [CrossRef]
- Birgy, A.; Madhi, F.; Jung, C.; Levy, C.; Cointe, A.; Bidet, P.; Hobson, C.A.; Bechet, S.; Sobral, E.; Vuthien, H.; et al. Diversity and trends in population structure of ESBL-producing Enterobacteriaceae in febrile urinary tract infections in children in France from 2014 to 2017. J. Antimicrob. Chemother. 2020, 75, 96–105. [Google Scholar] [CrossRef]
- Chen, X.; Zou, Q.; Zhang, W.; Wang, R.; Yu, F.; Chen, Y. Clinical features and microbiological characteristics of hospital- and community-onset Escherichia coli bloodstream infection. J. Med. Microbiol. 2019, 68, 178–187. [Google Scholar] [CrossRef]
- Tchesnokova, V.; Billig, M.; Chattopadhyay, S.; Linardopoulou, E.; Aprikian, P.; Roberts, P.L.; Skrivankova, V.; Johnston, B.; Gileva, A.; Igusheva, I.; et al. Predictive diagnostics for Escherichia coli infections based on the clonal association of antimicrobial resistance and clinical outcome. J. Clin. Microbiol. 2013, 51, 2991–2999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumura, Y.; Noguchi, T.; Tanaka, M.; Kanahashi, T.; Yamamoto, M.; Nagao, M.; Takakura, S.; Ichiyama, S. Population structure of Japanese extraintestinal pathogenic Escherichia coli and its relationship with antimicrobial resistance. J. Antimicrob. Chemother. 2017, 72, 1040–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, D.M.; Geyik, S.; Clermont, O.; O’Brien, C.L.; Huang, S.; Abayasekara, C.; Rajesh, A.; Kennedy, K.; Collignon, P.; Pavli, P.; et al. Fine-Scale structure analysis shows epidemic patterns of clonal complex 95, a cosmopolitan Escherichia coli lineage responsible for extraintestinal infection. mSphere 2017, 2, e00168-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olesen, B.; Scheutz, F.; Andersen, R.L.; Menard, M.; Boisen, N.; Johnston, B.; Hansen, D.S.; Krogfelt, K.A.; Nataro, J.P.; Johnson, J.R. Enteroaggregative Escherichia coli O78:H10, the cause of an outbreak of urinary tract infection. J. Clin. Microbiol. 2012, 50, 3703–3711. [Google Scholar] [CrossRef] [Green Version]
- Abe, C.M.; Salvador, F.A.; Falsetti, I.N.; Vieira, M.A.M.; Blanco, J.; Blanco, J.E.; Blanco, M.; Machado, A.M.O.; Elias, W.P.; Hernandes, R.T.; et al. Uropathogenic Escherichia coli (UPEC) strains may carry virulence properties of diarrhoeagenic E. coli. FEMS Immunol. Med. Microbiol. 2008, 52, 397–406. [Google Scholar] [CrossRef] [Green Version]
- Lara, F.B.M.; Nery, D.R.; de Oliveira, P.M.; Araujo, M.L.; Carvalho, F.R.Q.; Messias-Silva, L.C.F.; Ferreira, L.B.; Faria-Junior, C.; Pereira, A.L. Virulence markers and phylogenetic analysis of Escherichia coli strains with hybrid EAEC/UPEC genotypes recovered from sporadic cases of extraintestinal infections. Front. Microbiol. 2017, 8, 146. [Google Scholar] [CrossRef] [Green Version]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia coli phylo-typing method revisited: Improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef]
- Guinée, P.A.M.; Jansen, W.H.; Wadström, T.; Sellwood, R. Escherichia coli associated with neonatal diarrhoea in piglets and calves. In Laboratory Diagnosis in Neonatal Calf and Pig Diarrhoea; Current Topics in Veterinary Medicine and Animal Science; De Leeuw, P.W., Guinée, P.A.M., Eds.; Martinus Nijhoff Publishers: Leiden, The Netherlands, 1981; Volume 13, pp. 126–162. [Google Scholar] [CrossRef]
- Wirth, T.; Falush, D.; Lan, R.; Colles, F.; Mensa, P.; Wieler, L.H.; Karch, H.; Reeves, P.R.; Maiden, M.C.J.; Ochman, H.; et al. Sex and virulence in Escherichia coli: An evolutionary perspective. Mol. Microbiol. 2006, 60, 1136–1151. [Google Scholar] [CrossRef] [Green Version]
- Roer, L.; Johannesen, T.B.; Hansen, F.; Stegger, M.; Tchesnokova, V.; Sokurenko, E.; Garibay, N.; Allesøe, R.; Thomsen, M.C.F.; Lund, O.; et al. CHTyper, a web tool for subtyping of extraintestinal pathogenic Escherichia coli based on the fumC and fimH alleles. J. Clin. Microbiol. 2018, 56, e00063-18. [Google Scholar] [CrossRef] [Green Version]
- Matsumura, Y.; Pitout, J.D.D.; Peirano, G.; DeVinney, R.; Noguchi, T.; Yamamoto, M.; Gomi, R.; Matsuda, T.; Nakano, S.; Nagao, M.; et al. Rapid identification of different Escherichia coli sequence type 131 clades. Antimicrob. Agents Chemother. 2017, 61, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Dahbi, G.; Mora, A.; Mamani, R.; López, C.; Alonso, M.P.; Marzoa, J.; Blanco, M.; Herrera, A.; Viso, S.; García-Garrote, F.; et al. Molecular epidemiology and virulence of Escherichia coli O16:H5-ST131: Comparison with H30 and H30-Rx subclones of O25b:H4-ST131. Int. J. Med. Microbiol. 2014, 304, 1247–1257. [Google Scholar] [CrossRef] [PubMed]
- Blanco, M.; Blanco, J.E.; Dahbi, G.; Alonso, M.P.; Mora, A.; Coira, M.A.; Madrid, C.; Juarez, A.; Bernardez, M.I.; Gonzalez, E.A.; et al. Identification of two new intimin types in atypical enteropathogenic Escherichia coli. Int. Microbiol. 2006, 9, 103–110. [Google Scholar] [PubMed]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 27th ed.; CLSI Supplement M100S; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco, M.; Alonso, M.P.; Nicolas-Chanoine, M.-H.; Dahbi, G.; Mora, A.; Blanco, J.E.; López, C.; Cortés, P.; Llagostera, M.; Leflon-Guibout, V.; et al. Molecular epidemiology of Escherichia coli producing extended-spectrum β-Lactamases in Lugo (Spain): Dissemination of clone O25b:H4-ST131 producing CTX-M-15. J. Antimicrob. Chemother. 2009, 63, 1135–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leflon-Guibout, V.; Jurand, C.; Bonacorsi, S.; Espinasse, F.; Guelfi, M.C.; Duportail, F.; Heym, B.; Bingen, E.; Nicolas-Chanoine, M.-H. Emergence and spread of three clonally related virulent isolates of CTX-M-15-producing Escherichia coli with variable resistance to aminoglycosides and tetracycline in a French geriatric hospital. Antimicrob. Agents Chemother. 2004, 48, 3736–3742. [Google Scholar] [CrossRef] [PubMed] [Green Version]







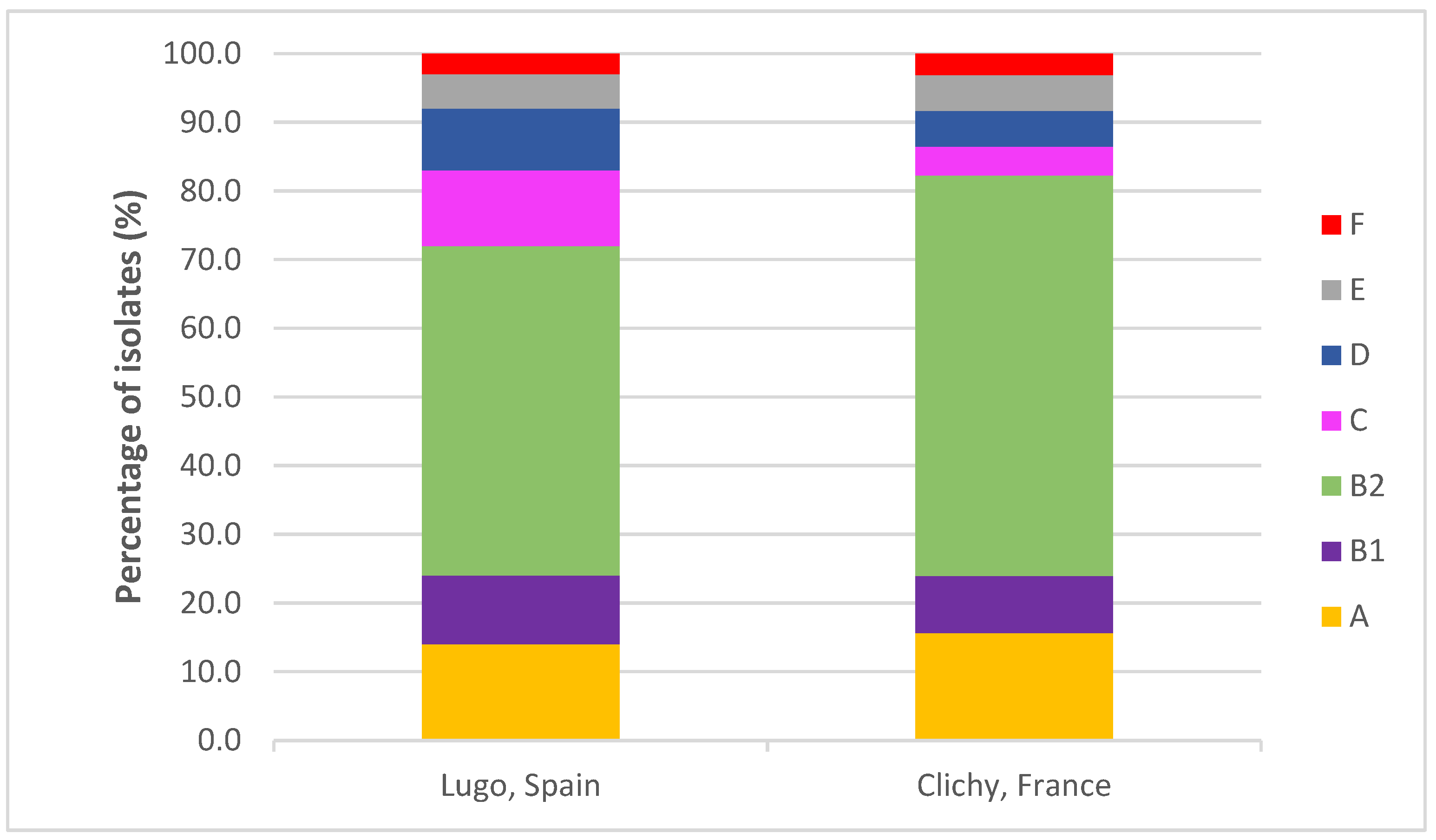{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clone (Number of Isolates from Spain and France) a,b | Serotype (Number of Isolates) | ExPEC Status | UPEC Status | MDR Status |
|---|---|---|---|---|
| A-CH11-54-ST10 (1/2) | O48:HNM (1), O133:H40 (1), O153:HNT (1) | 1 | 0 | 0 |
| B1-CH4-27-ST58 (2/1) | O8:HNM (1), O9:H4 (1), O9:H25 (1) | 0 | 0 | 1 |
| B2-CH13-27-ST12 (1/1) | O4:H5 (1), O4:HNM (1) | 2 | 2 | 0 |
| B2-CH13-106-ST12 (2/2) | O4:H1 (3), O4:HNM (1) | 4 | 4 | 0 |
| B2-CH24-10-ST73 (2/4) | O2:H1 (1), O2:HNM (1), O6:H1 (3), O6:HNM (1) | 6 | 6 | 1 |
| B2-CH24-12-ST73 (2/1) | O6:H1 (1), O25:H1 (1), O25:H4 (1) | 3 | 3 | 0 |
| B2-CH24-30-ST73 (0/3) | O6:H1 (3) | 3 | 3 | 1 |
| B2-CH24-103-ST73 (4/2) | O6:H1 (4), O22:H1 (1), O22:HNM (1) | 6 | 6 | 1 |
| B2-CH38-15-ST95 (2/2) | O2:H5 (1), O2:H7 (1), O18:H7 (2) | 4 | 4 | 0 |
| B2-CH38-27-ST95 (1/1) | O2:H4 (1), O25:H4 (1) | 2 | 2 | 1 |
| B2-CH38-41-ST95 (1/1) | O1:H7 (2) | 2 | 2 | 0 |
| B2-CH14-2-ST127 (1/2) | O6:H31 (1), O6:HNM (2) | 3 | 3 | 1 |
| B2-CH14-136-ST127 (1/1) | O6:H31 (1), O6:HNM (1) | 2 | 2 | 0 |
| B2-CH40-22-ST131 (1/2) | O25:H4 (3) | 3 | 3 | 2 |
| B2-CH40-30-ST131 (5/8) | O25:H4 (13) | 8 | 13 | 12 |
| B2-CH40-41-ST131 (5/1) | O4:H5 (2), O4:HNM (1) O12:HNM (1), ONT:H5 (2) | 3 | 3 | 4 |
| B2-CH52-5-ST141 (3/8) | O2:H6 (8), O14:HNM (1), ONT:H4 (1), ONT:H6 (1) | 11 | 11 | 0 |
| B2-CH52-14-ST141 (0/2) | O2:H6 (2) | 2 | 2 | 0 |
| B2-CH103-9-ST372 (1/1) | O18:H31 (2) | 0 | 2 | 0 |
| B2-CH14-64-ST1193 (1/1) | O75:HNM (1), ONT:HNM (1) | 1 | 2 | 2 |
| C-CH4-39-ST88 (9/1) | O8:H4 (5), O9:H4 (3), O9:HNM (1), ONT:H4 (1) | 5 | 0 | 9 |
| D-CH35-27-ST69 (8/5) | O15:H18 (2), O15:HNM (1), O18:HNM (1), O44:H18 (2), O77:HNM (1), O106:H4 (2), O175:HNM (1), ONT:H18 (3) | 9 | 0 | 6 |
| E-CH100-96-ST362 (2/1) | O7:H6 (2), O21:HNT (1) | 1 | 0 | 3 |
| E-CH37-27-ST405 (1/1) | O18:H6 (1), O102:H4 (1) | 1 | 0 | 1 |
| F-CH32-41-ST59 (3/0) | O1:H7 (3) | 3 | 3 | 0 |
| F-CH4-58-ST648 (0/2) | O25:H4 (1), ONT:H42 (1) | 2 | 1 | 1 |
| VF Gene | Number (%) of Isolates | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Total (n = 196) | B2 ST131 (n = 23) | A ST10 (n = 9) | B2 ST12 (n = 8) | B1 ST58 (n = 5) | D ST69 (n = 13) | B2 ST73 (n = 21) | C ST88 (n = 12) | B2 ST95 (n = 10) | B2 ST127 (n = 7) | B2 ST141 (n = 14) | |
| Adhesins | |||||||||||
| fimH | 193 (98.5) | 23 | 9 | 8 | 5 | 13 | 21 | 12 | 10 | 7 | 14 |
| fimAvMT78 | 27 (13.8) | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 0 |
| papAH | 84 (42.9) | 6 | 1 | 8 | 0 | 6 | 18 | 10 | 7 | 6 | 7 |
| papC | 87 (44.4) | 6 | 1 | 8 | 0 | 7 | 18 | 10 | 7 | 6 | 7 |
| papEF | 92 (46.9) | 6 | 1 | 8 | 0 | 11 | 18 | 10 | 7 | 6 | 7 |
| sfa/focDE | 58 (29.6) | 1 | 0 | 8 | 0 | 0 | 20 | 0 | 2 | 5 | 13 |
| afa/draBC | 11(5.6) | 7 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| yfcV | 106 (54.1) | 20 | 0 | 8 | 0 | 0 | 21 | 0 | 10 | 7 | 14 |
| Toxins | |||||||||||
| sat | 47 (24.0) | 13 | 3 | 0 | 0 | 9 | 12 | 0 | 0 | 0 | 0 |
| cnf1 | 49 (25.0) | 4 | 0 | 8 | 0 | 0 | 17 | 0 | 0 | 7 | 7 |
| hlyA | 55 (28.1) | 3 | 1 | 8 | 0 | 0 | 20 | 0 | 0 | 7 | 7 |
| hlyF | 44 (22.4) | 2 | 2 | 0 | 4 | 0 | 0 | 8 | 7 | 0 | 3 |
| cdtB | 12 (6.1) | 3 | 0 | 0 | 0 | 0 | 6 | 0 | 2 | 0 | 0 |
| tsh | 7 (3.6) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 1 |
| vat | 80 (40.8) | 0 | 0 | 7 | 0 | 0 | 20 | 0 | 10 | 7 | 14 |
| Iron uptake | |||||||||||
| iucD | 99 (50.5) | 16 | 6 | 2 | 4 | 11 | 12 | 8 | 7 | 0 | 3 |
| iutA | 99 (50.5) | 16 | 6 | 2 | 4 | 11 | 12 | 8 | 7 | 0 | 3 |
| iroN | 93 (47.4) | 3 | 2 | 8 | 4 | 0 | 20 | 8 | 6 | 4 | 14 |
| fyuA | 162 (82.7) | 23 | 7 | 8 | 4 | 12 | 21 | 10 | 10 | 7 | 14 |
| chuA | 135 (68.9) | 23 | 0 | 8 | 0 | 12 | 21 | 0 | 10 | 7 | 14 |
| Capsule | |||||||||||
| kpsM II | 119 (60.7) | 17 | 2 | 4 | 0 | 10 | 20 | 0 | 10 | 7 | 14 |
| neuC-K1 | 35 (17.9) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 10 | 1 | 12 |
| kpsM II-K2 | 17 (8.7) | 7 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 |
| kpsM II-K5 | 67 (34.2) | 10 | 2 | 4 | 0 | 7 | 20 | 0 | 0 | 6 | 2 |
| kpsM III | 6 (3.1) | 0 | 0 | 3 | 0 | 1 | 0 | 0 | 0 | 0 | 0 |
| Miscellaneous | |||||||||||
| cvaC | 35 (17.9) | 2 | 2 | 0 | 4 | 0 | 0 | 7 | 7 | 0 | 3 |
| iss | 46 (23.5) | 2 | 2 | 1 | 4 | 0 | 2 | 8 | 5 | 0 | 5 |
| traT | 111 (56.6) | 17 | 4 | 6 | 5 | 7 | 4 | 9 | 8 | 3 | 6 |
| ibeA | 23 (11.7) | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 3 |
| malX | 109 (55.6) | 21 | 0 | 8 | 0 | 0 | 21 | 0 | 10 | 7 | 14 |
| usp | 107 (54.6) | 23 | 0 | 8 | 0 | 0 | 21 | 0 | 10 | 7 | 14 |
| ompT | 151 (77.0) | 23 | 2 | 8 | 4 | 12 | 21 | 10 | 10 | 7 | 14 |
| ExPEC status | 121 (61.7) | 15 | 2 | 8 | 0 | 9 | 21 | 6 | 10 | 7 | 14 |
| UPEC status | 106 (54.1) | 20 | 0 | 8 | 0 | 0 | 21 | 0 | 10 | 7 | 14 |
| Range of VFs | 1 to 23 | 8 to 20 | 1 to 14 | 16-19 | 2-10 | 5-12 | 13-20 | 6-13 | 13-22 | 13-17 | 12-23 |
| Mean of VFs | 11.6 | 12.3 | 6.3 | 17.1 | 8.4 | 9.4 | 17.4 | 9.8 | 18.2 | 15.4 | 16.2 |
| Drug a | Number (%) of Resistant Isolates | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Total (n=196) | B2 ST131 (n=23) | A ST10 (n=9) | B2 ST12 (n=8) | B1 ST58 (n=5) | D ST69 (n=13) | B2 ST73 (n=21) | C ST88 (n=12) | B2 ST95 (n=10) | B2 ST127 (n=7) | B2 ST141 (n=14) | |
| Ampicillin AM10 | 111 (56.6)b | 19 (82.6) | 4 (44.4) | 7 (87.5) | 5 (100) | 8 (61.5) | 6 (28.6) | 12 (100) | 3 (30) | 4 (57.1) | 3 (21.4) |
| Amoxicillin-Clavulanate AMC30 | 27 (13.8) | 7 (30.4) | 0 | 2 (25%) | 0 | 1 (7.7) | 1 (4.8) | 9 (75) | 0 | 0 | 0 |
| Cefazolin CZ30 | 27 (13.8) | 13 (56.5) | 0 | 0 | 1 (20) | 0 | 0 | 3 (25) | 0 | 0 | 0 |
| Cefuroxime CXM30 | 21 (10.7) | 7 (30.4) | 1 (11.1) | 0 | 1 (20) | 0 | 0 | 3 (25) | 0 | 0 | 0 |
| Cefotaxime CTX30 | 15 (7.7) | 6 (26.1) | 0 | 0 | 1 (20) | 0 | 0 | 0 | 0 | 0 | 0 |
| Ceftazidime CAZ30 | 3 (1.5) | 1 (4.3) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Cefepime FED30 | 9 (4.6) | 2 (8.7) | 0 | 0 | 1 (20) | 0 | 0 | 1 (8.3) | 0 | 0 | 0 |
| Cefoxitin FOX30 | 7 (3.6) | 2 (8.7) | 0 | 0 | 0 | 0 | 0 | 1 (8.3) | 0 | 0 | 0 |
| Aztreonam ATM30 | 9 (4.6) | 2 (8.7) | 0 | 0 | 0 | 0 | 0 | 1 (8.3) | 0 | 0 | 0 |
| Gentamicin GM10 | 17 (3.6) | 4 (17.4) | 0 | 0 | 0 | 1 (7.7) | 0 | 5 (41.7) | 0 | 0 | 0 |
| Tobramycin NN10 | 13 (6.6) | 2 (8.7) | 0 | 0 | 0 | 1 (7.7) | 0 | 5 (41.7) | 0 | 0 | 0 |
| Doxycycline D30 | 60 (30.6) | 6 (26.1) | 3 (33.3) | 1 (12.5) | 3 (60) | 5 (38.5) | 4 (19) | 10 (83.3) | 1 (10) | 3 (42.9) | 0 |
| Chloramphenicol C30 | 23 (11.7) | 1 (4.3) | 1 (11.1) | 1 (12.5) | 0 | 1 (7.7) | 2 (9.5) | 9 (75) | 0 | 1 (14.3) | 0 |
| Nitrofurantoin FD300 | 2 (1.0) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Nalidixic Acid NAL30 | 65 (33.2) | 17 (73.9) | 2 (22.2) | 3 (37.5) | 3 (60) | 7 (53.8) | 0 | 9 (75) | 2 (20) | 0 | 0 |
| Ciprofloxacin CIP5 | 42 (21.4) | 13 (56.5) | 1 (11.1) | 0 | 1 (20) | 0 | 0 | 9 (75) | 1 (10) | 0 | 0 |
| Trimethoprim-Sulfamethoxazole SXT25 | 57 (29.1) | 9 (39.1) | 2 (22.2) | 2 (25%) | 3 (60) | 7 (53.8) | 3 (14.3) | 9 (75) | 1 (10) | 1 (14.3) | 1 (7.1) |
| Multidrug resistance | 73 (37.2) | 18 (78.3) | 3 (33.3) | 1 (12.5) | 2 (40) | 6 (42.6) | 3 (14.3) | 10 (83.3) | 1 (10) | 2 (28.6) | 0 |
| ESBL | 13 (6.6) | 5 (21.7) | 0 | 0 | 1 (20.0) | 0 | 0 | 0 | 0 | 0 | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flament-Simon, S.-C.; Nicolas-Chanoine, M.-H.; García, V.; Duprilot, M.; Mayer, N.; Alonso, M.P.; García-Meniño, I.; Blanco, J.E.; Blanco, M.; Blanco, J. Clonal Structure, Virulence Factor-encoding Genes and Antibiotic Resistance of Escherichia coli, Causing Urinary Tract Infections and Other Extraintestinal Infections in Humans in Spain and France during 2016. Antibiotics 2020, 9, 161. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9040161
Flament-Simon S-C, Nicolas-Chanoine M-H, García V, Duprilot M, Mayer N, Alonso MP, García-Meniño I, Blanco JE, Blanco M, Blanco J. Clonal Structure, Virulence Factor-encoding Genes and Antibiotic Resistance of Escherichia coli, Causing Urinary Tract Infections and Other Extraintestinal Infections in Humans in Spain and France during 2016. Antibiotics. 2020; 9(4):161. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9040161
Chicago/Turabian StyleFlament-Simon, Saskia-Camille, Marie-Hélène Nicolas-Chanoine, Vanesa García, Marion Duprilot, Noémie Mayer, María Pilar Alonso, Isidro García-Meniño, Jesús E. Blanco, Miguel Blanco, and Jorge Blanco. 2020. "Clonal Structure, Virulence Factor-encoding Genes and Antibiotic Resistance of Escherichia coli, Causing Urinary Tract Infections and Other Extraintestinal Infections in Humans in Spain and France during 2016" Antibiotics 9, no. 4: 161. https://0-doi-org.brum.beds.ac.uk/10.3390/antibiotics9040161