
Effects of Local Acidification on Benthic Communities at Shallow Hydrothermal Vents of the Aeolian Islands (Southern Tyrrhenian, Mediterranean Sea)

 ,
,  , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
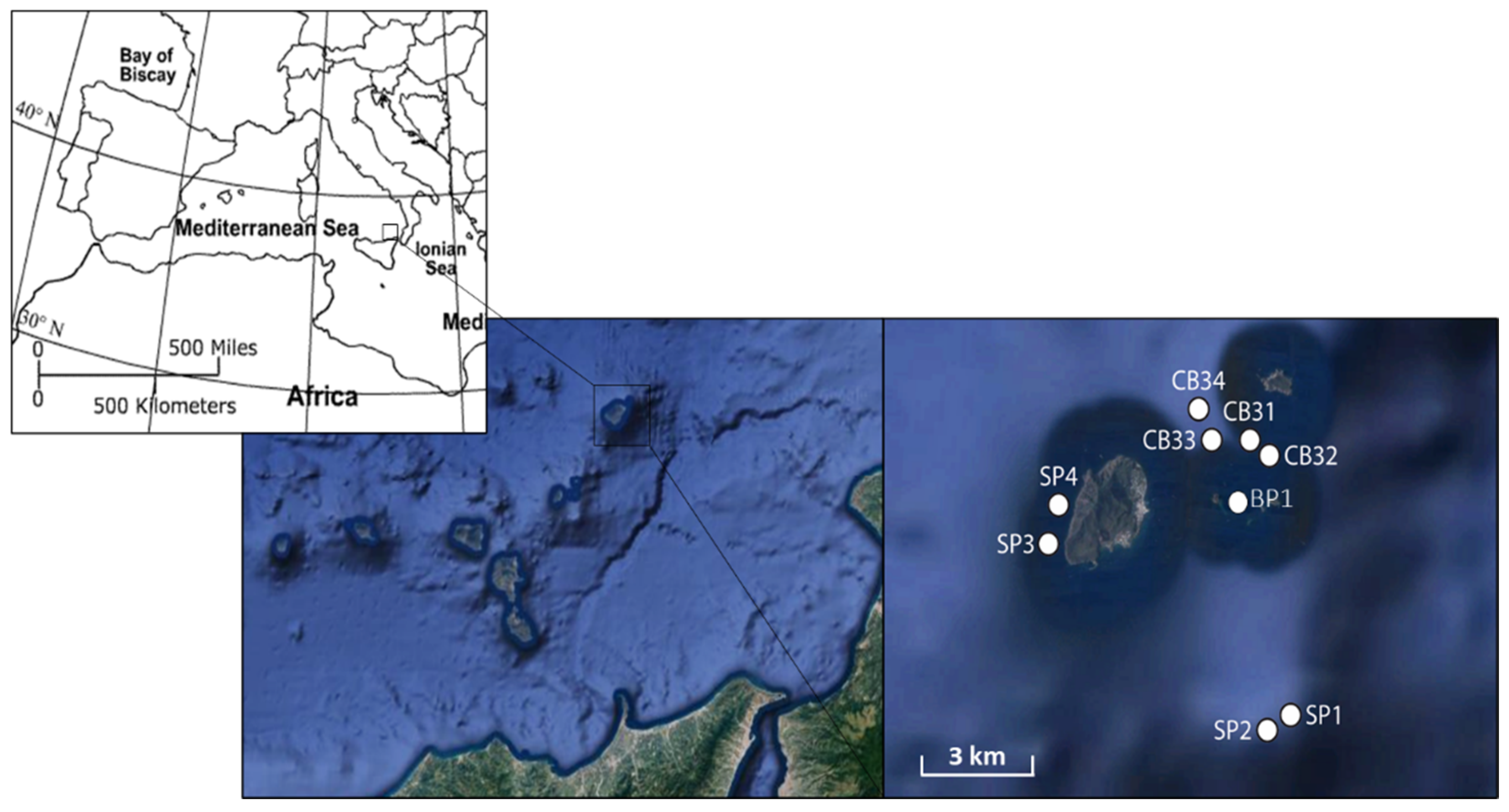
2.1. Study Area
2.2. Sampling Strategy and Samples Collection
2.3. Biochemical Composition of Sedimentary Organic Matter
2.4. Meio- and Macrofauna Samples Processing
2.5. Data Analysis
3. Results
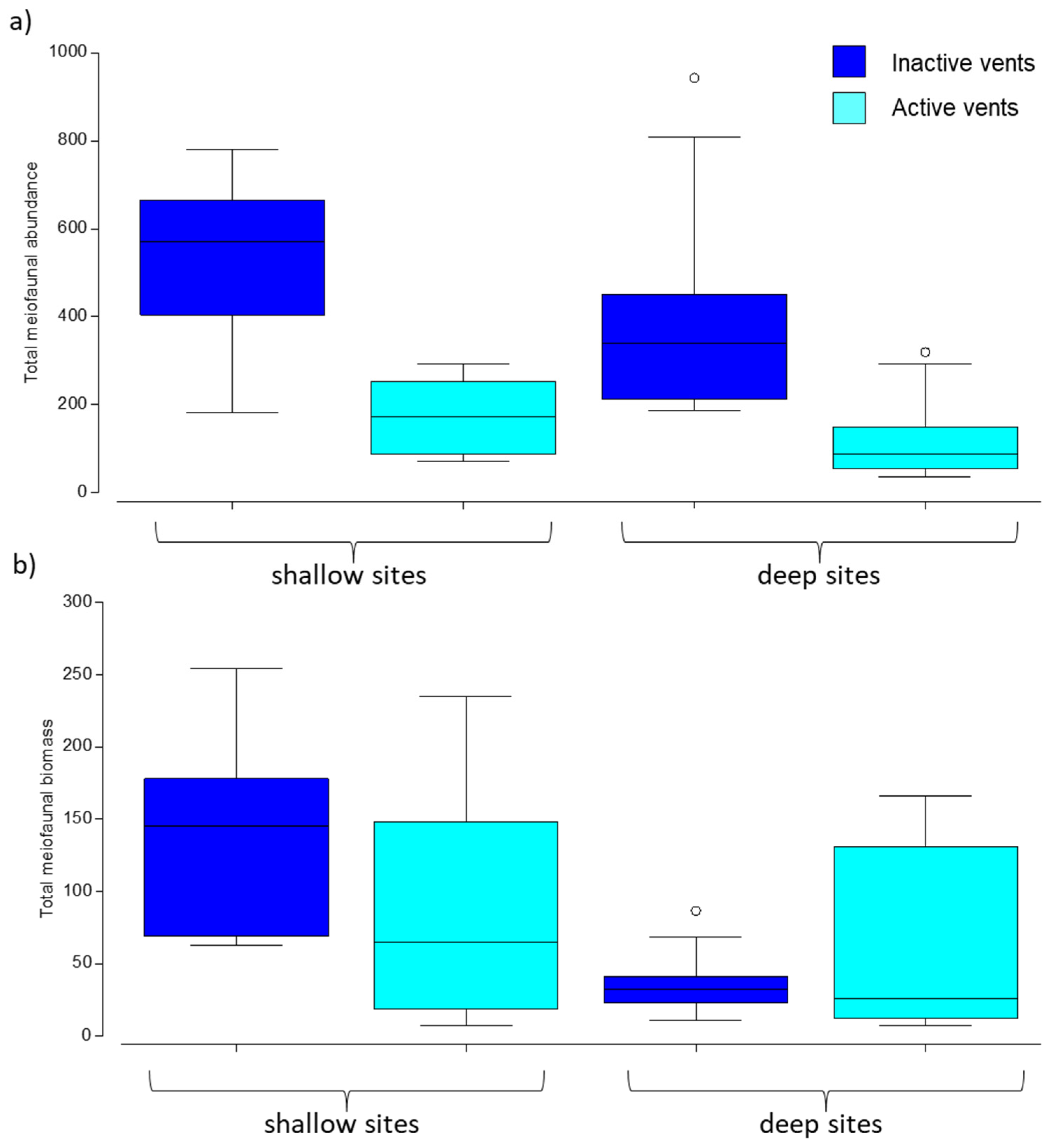
3.1. Abundance and Biomass of Meiofauna
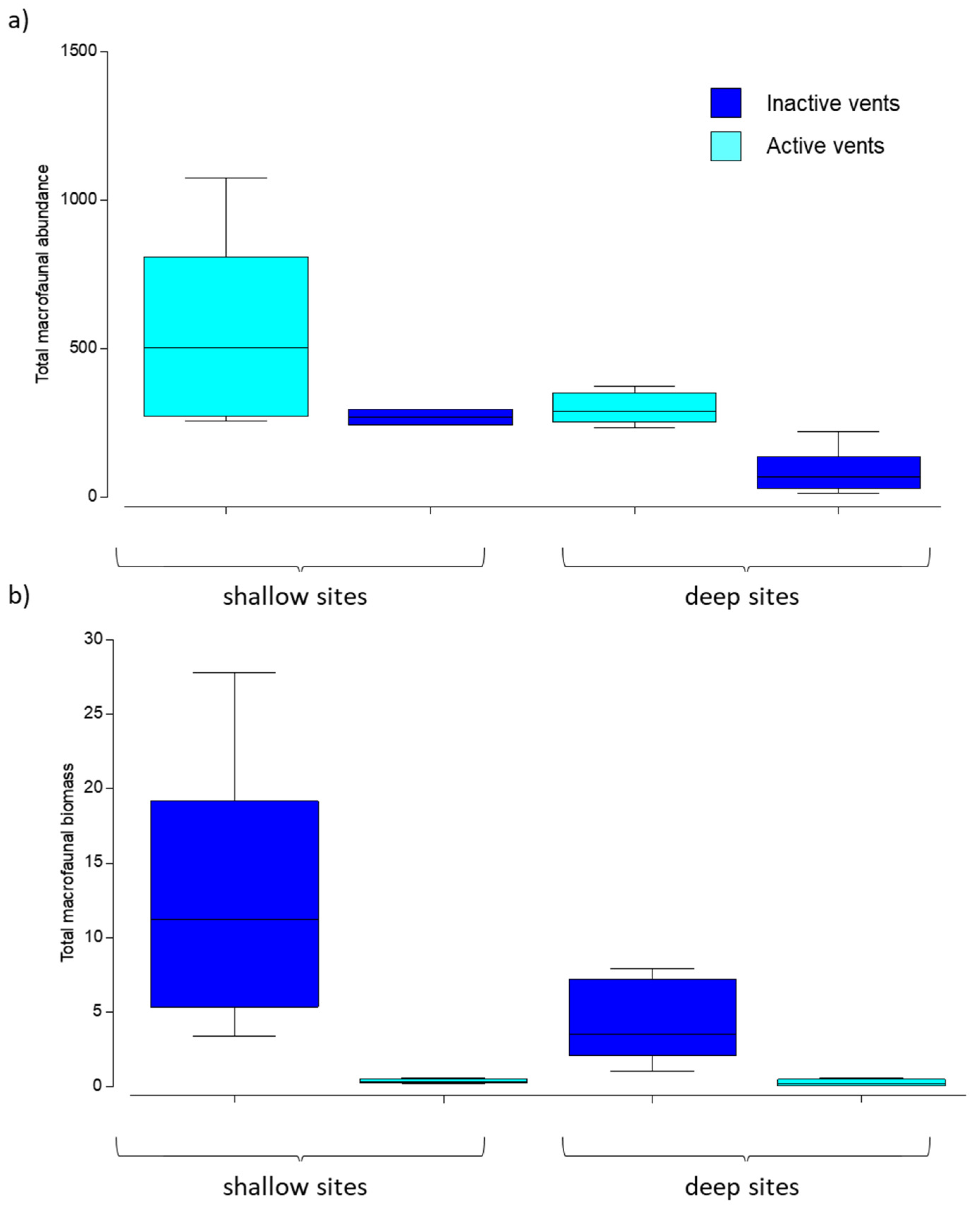
3.2. Abundance and Biomass of Macrofauna
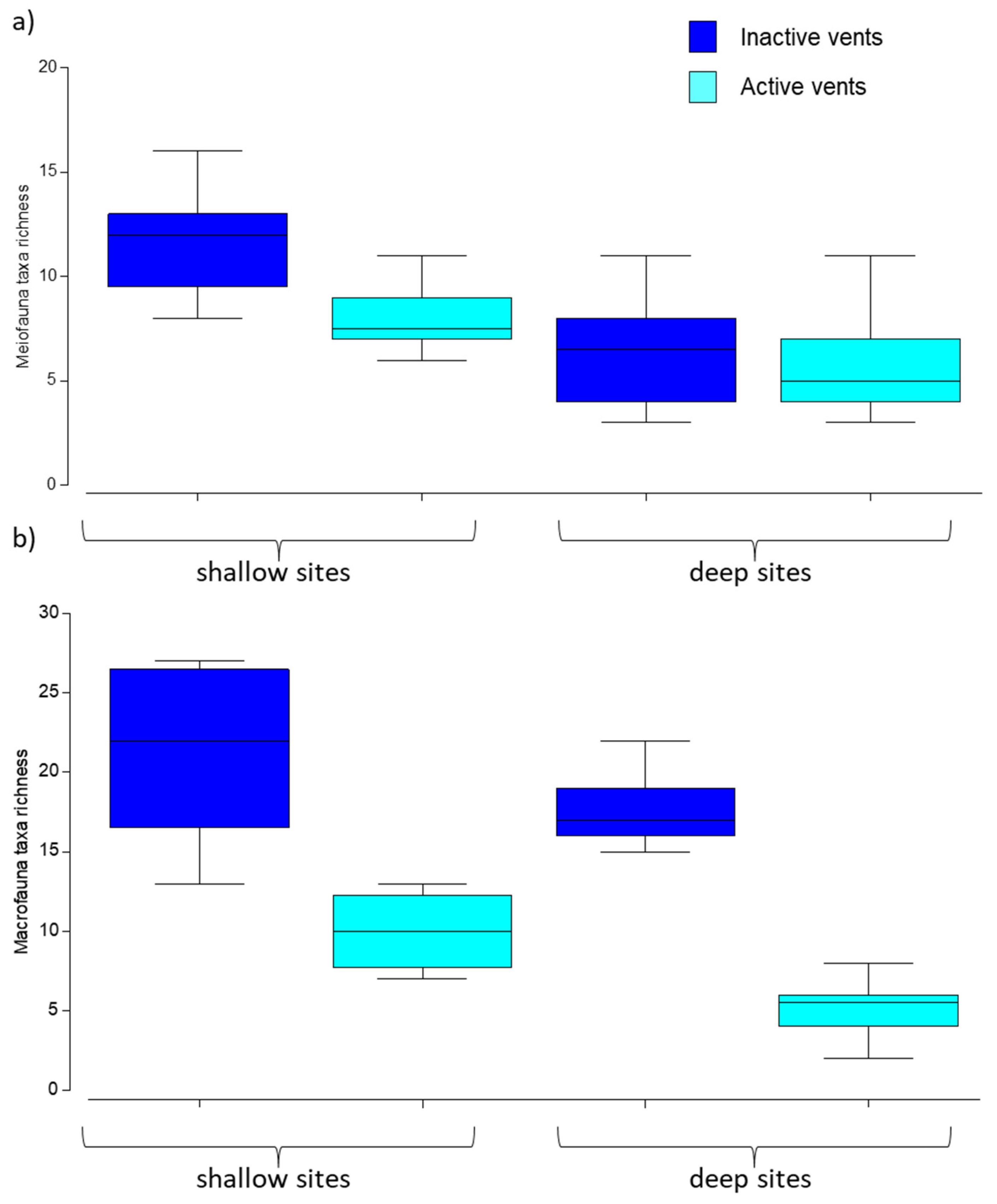
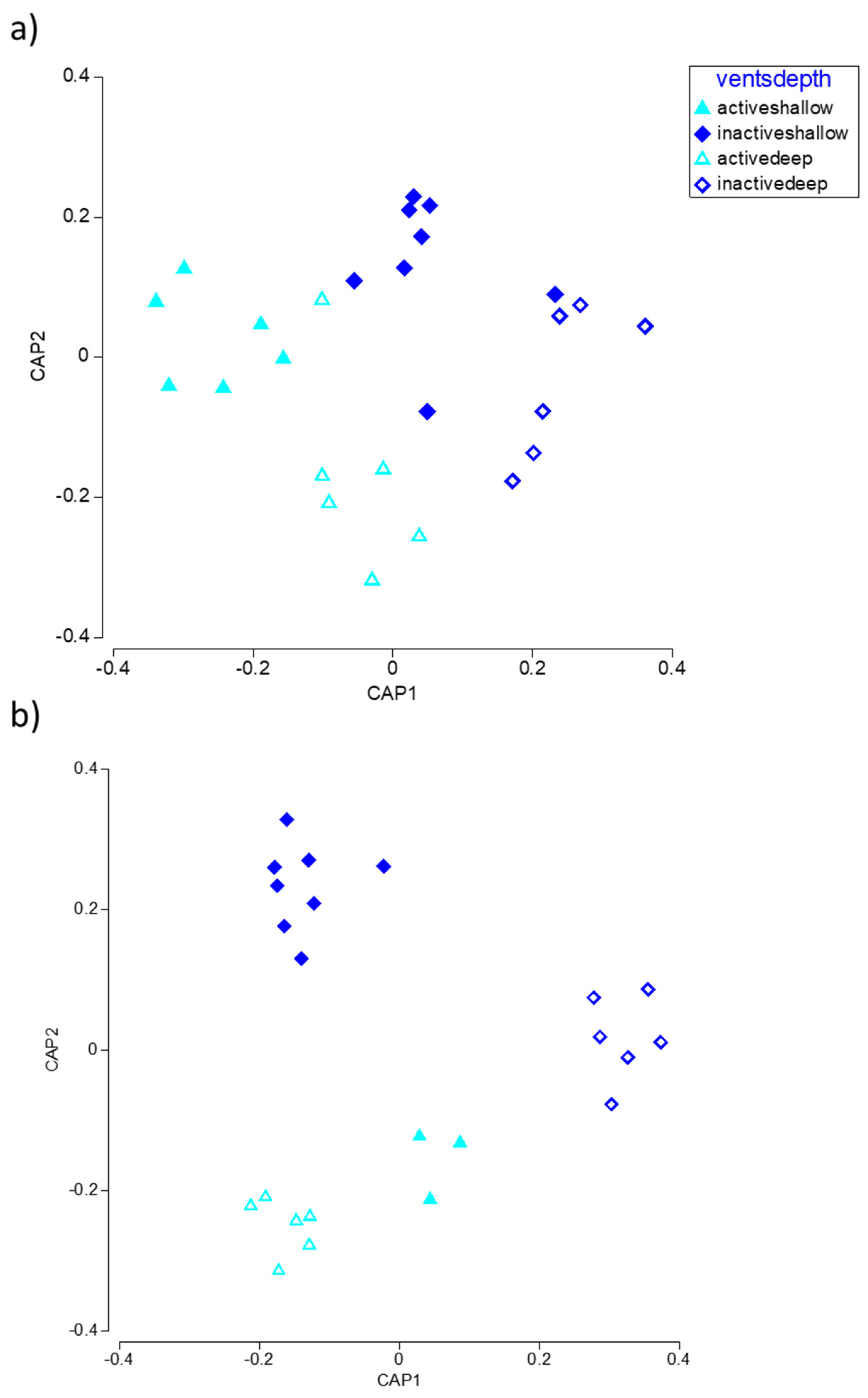
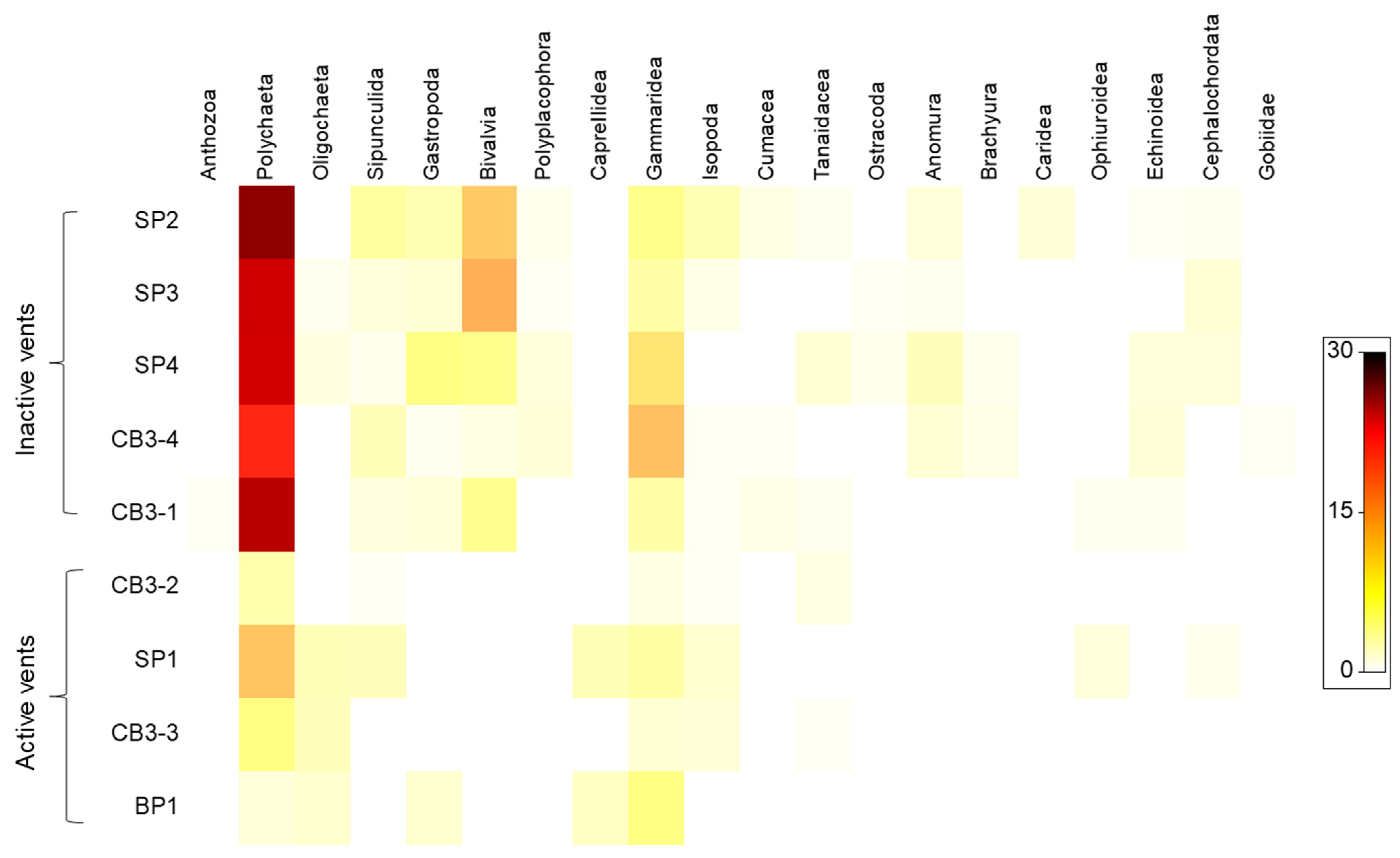
3.3. Meio- and Macrofaunal Diversity and Community Composition
3.4. Correlation of Meio- and Macrofaunal Communities and Organic Matter Quality and Quantity
4. Discussion
4.1. Changes in Meio- and Macrobenthic Abundance and Biomass Due to CO2 Emission
4.2. Effects of Acidification on Meio- and Macrofauna Community Composition and Diversity
4.3. Ocean Acidification Changes in Food Availability as Drivers of Community Composition Shifts
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Intergovernmental Panel on Climate Change. Climate change 2021: The physical science basis. In Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Doney, S.C.; Busch, D.S.; Cooley, S.R.; Kroeker, K.J. The impacts of ocean acidification on marine ecosystems and reliant human communities. Annu. Rev. Environ. Resour. 2020, 45, 83–112. [Google Scholar] [CrossRef]
- Tittensor, D.P.; Baco, A.R.; Brewin, P.E.; Clark, M.R.; Consalvey, M.; Hall-Spencer, J.; Rowden, A.A.; Schlacher, T.; Stocks, K.I.; Rogers, A.D. Predicting global habitat suitability for stony corals on seamounts. J. Biogeogr. 2009, 36, 1111–1128. [Google Scholar] [CrossRef] [Green Version]
- Kilias, S.P.; Nomikou, P.; Papanikolaou, D.; Polymenakou, P.N.; Godelitsas, A.; Argyraki, A.; Carey, S.; Gamaletsos, P.; Mertzimekis, T.J.; Stathopoulou, E.; et al. New insights into hydrothermal vent processes in the unique shallow-submarine arc-volcano, Kolumbo (Santorini), Greece. Sci. Rep. 2013, 3, 2421. [Google Scholar] [CrossRef] [PubMed]
- Cerrano, C.; Cardini, U.; Bianchelli, S.; Corinaldesi, C.; Pusceddu, A.; Danovaro, R. Red coral extinction risk enhanced by ocean acidification. Sci. Rep. 2013, 3, 1457. [Google Scholar] [CrossRef] [Green Version]
- Koch, M.; Bowes, G.; Ross, C.; Zhang, X.-H. Climate change and ocean acidification effects on seagrasses and marine macroalgae. Glob. Chang. Biol. 2013, 19, 103–132. [Google Scholar] [CrossRef]
- Ravaglioli, C.; Bulleri, F.; Rühl, S.; McCoy, S.J.; Findlay, H.S.; Widdicombe, S.; Queirós, A.M. Ocean acidification and hypoxia alter organic carbon fluxes in marine soft sediments. Glob. Chang. Biol. 2019, 25, 4165–4178. [Google Scholar] [CrossRef]
- Zeppilli, D.; Danovaro, R. Meiofaunal diversity and assemblage structure in a shallow-water hydrothermal vent in the Pacific Ocean. Aquat. Biol. 2009, 5, 75–84. [Google Scholar] [CrossRef] [Green Version]
- Danovaro, R.; Scopa, M.; Gambi, C.; Fraschetti, S. Trophic importance of subtidal metazoan meiofauna: Evidence from in situ exclusion experiments on soft and rocky substrates. Mar. Biol. 2007, 152, 339–350. [Google Scholar] [CrossRef]
- Carpentier, A.; Como, S.; Dupuy, C.; Lefrançois, C.; Feunteun, E. Feeding ecology of Liza spp. in a tidal flat: Evidence of the importance of primary production (biofilm) and associated meiofauna. J. Sea Res. 2014, 92, 86–91. [Google Scholar] [CrossRef] [Green Version]
- Hall-Spencer, J.M.; Rodolfo-Metalpa, R.; Martin, S.; Ransome, E.; Fine, M.; Turner, S.M.; Rowley, S.J.; Tedesco, D.; Buia, M.-C. Volcanic carbon dioxide vents show ecosystem effects of ocean acidification. Nature 2008, 454, 96–99. [Google Scholar] [CrossRef] [Green Version]
- Kroeker, K.J.; Micheli, F.; Gambi, M.C.; Martz, T.R. Divergent ecosystem responses within a benthic marine community to ocean acidification. Proc. Natl. Acad. Sci. USA 2011, 108, 14515–14520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guilini, K.; Weber, M.; de Beer, D.; Schneider, M.; Molari, M.; Lott, C.; Bodnar, W.; Mascart, T.; De Troch, M.; Vanreusel, A. Response of Posidonia oceanica seagrass and its epibiont communities to ocean acidification. PLoS ONE 2017, 12, e0181531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiss, H.; Birchenough, S.; Borja, A.; Buhl-Mortensen, L.; Craeymeersch, J.; Dannheim, J.; Darr, A.; Galparsoro, I.; Gogina, M.; Neumann, H.; et al. Benthos distribution modelling and its relevance for marine ecosystem management. ICES J. Mar. Sci. 2014, 72, 297–315. [Google Scholar] [CrossRef] [Green Version]
- Borja, A.; Elliott, M.; Andersen, J.H.; Berg, T.; Carstensen, J.; Halpern, B.S.; Heiskanen, A.-S.; Korpinen, S.; Lowndes, J.S.S.; Martin, G.; et al. Overview of integrative assessment of marine systems: The ecosystem approach in practice. Front. Mar. Sci. 2016, 3, 20. [Google Scholar] [CrossRef] [Green Version]
- Van Hoey, G.; Borja, A.; Birchenough, S.; Buhl-Mortensen, L.; Degraer, S.; Fleischer, D.; Kerckhof, F.; Magni, P.; Muxika, I.; Reiss, H.; et al. The use of benthic indicators in Europe: From the Water Framework Directive to the Marine Strategy Framework Directive. Mar. Pollut. Bull. 2010, 60, 2187–2196. [Google Scholar] [CrossRef] [Green Version]
- Ellis, J.I.; Norkko, A.; Thrush, S.F. Broad-scale disturbance of intertidal and shallow sublittoral soft-sediment habitats; effects on the benthic macrofauna. J. Aquat. Ecosyst. Stress Recovery 2000, 7, 57–74. [Google Scholar] [CrossRef]
- Gray, J.S. The Ecology of Marine Sediments: An Introduction to the Structure and Function of Benthic Communities; Cambridge University Press: Cambridge, UK, 1981. [Google Scholar]
- Danovaro, R.; Gambi, C.; Höss, S.; Mirto, S.; Traunspurger, W.; Zullini, A. Case studies using nematode assemblage analysis in aquatic habitats. In Nematodes as Environmental Indicators; Wilson, M.J., Kakouli-Duarte, T., Eds.; CABI: Wallingford, UK, 2009; pp. 146–171. [Google Scholar] [CrossRef]
- Gambi, C.; Dell’Anno, A.; Corinaldesi, C.; Lo Martire, M.; Musco, L.; Da Ros, Z.; Armiento, G.; Danovaro, R. Impact of historical contamination on meiofaunal assemblages: The case study of the Bagnoli-Coroglio Bay (southern Tyrrhenian Sea). Mar. Environ. Res. 2020, 156, 104907. [Google Scholar] [CrossRef]
- Giere, O. Meiobenthology: The Microscopic Motile Fauna of Aquatic Sediments, 2nd ed.; Springer GmbH: Heidelberg/Berlin, Germany, 2009; pp. 1–527. [Google Scholar] [CrossRef]
- Italiano, F. Hydrothermal fluids vented at shallow depths at the Aeolian Islands: Relationships with volcanic and geothermal systems. FOG Freib. Online Geosci. 2009, 22, 55–60. [Google Scholar]
- Italiano, F.; Nuccio, P.M. Geochemical investigations of submarine volcanic exhalations to the east of Panarea, Aeolian Islands, Italy. J. Volcanol. Geotherm. Res. 1991, 46, 125–141. [Google Scholar] [CrossRef]
- Caracausi, A.; Ditta, M.; Italiano, F.; Longo, M.; Nuccio, P.M.; Paonita, A.; Rizzo, A. Changes in fluid geochemistry and physico-chemical conditions of geothermal systems caused by magmatic input: The recent abrupt outgassing off the island of Panarea (Aeolian Islands, Italy). Geochim. Cosmochim. Acta 2005, 69, 3045–3059. [Google Scholar] [CrossRef]
- Heinicke, J.; Maugeri, R.; Merkel, B.; Pohl, T.; Schipek, M.; Braun, T. Evidence of tectonic control on active arc volcanism: The Panarea-Stromboli tectonic link inferred by submarine hydrothermal vents monitoring (Aeolian arc, Italy). Geophys. Res. Lett. 2009, 36, 4301. [Google Scholar] [CrossRef] [Green Version]
- Savelli, C.; Marani, M.; Gamberi, F. Geochemistry of metalliferous, hydrothermal deposits in the Aeolian arc (Tyrrhenian Sea). J. Volcanol. Geotherm. Res. 1999, 88, 305–323. [Google Scholar] [CrossRef]
- Esposito, V.; Andaloro, F.; Canese, S.; Bortoluzzi, G.; Bo, M.; Di Bella, M.; Italiano, F.; Sabatino, G.; Battaglia, P.; Consoli, P.; et al. Exceptional discovery of a shallow-water hydrothermal site in the SW area of Basiluzzo islet (Aeolian archipelago, South Tyrrhenian Sea): An environment to preserve. PLoS ONE 2018, 13, e0190710. [Google Scholar] [CrossRef] [PubMed]
- Romagnoli, C.; Bortoluzzi, G.; Chiocci, F.; D’Oriano, F.; Gamberi, F.; Ligi, M.; Marani, M. Bathy-morphological setting of the Aeolian Islands. In The Aeolian Island Volcanoes; Lucchi, F., Peccerillo, A., Keller, J., Tranne, C.A., Rossi, P.L., Eds.; Geological Society of London: London, UK, 2013; Volume 38, pp. 27–36. [Google Scholar]
- Capaccioni, B.; Tassi, F.; Vaselli, O.; Tedesco, D.; Poreda, R. Submarine gas burst at Panarea Island (southern Italy) on 3 November 2002: A magmatic versus hydrothermal episode. J. Geophys. Res. 2007, 112, B050201. [Google Scholar] [CrossRef] [Green Version]
- Danovaro, R. Methods for the Study of Deep-Sea Sediments, Their Functioning and Biodiversity; CRC Press: Boca Raton, FL, USA, 2010; p. 428. [Google Scholar]
- International Atomic Energy Agency. Collection and Preparation of Bottom Sediment Samples for Analysis of Radionuclides and Trace Elements; TECDOC Series; IAEA: Vienna, Austria, 2003. [Google Scholar]
- Tangherlini, M.; Corinaldesi, C.; Ape, F.; Greco, S.; Romeo, T.; Andaloro, F.; Danovaro, R. Ocean acidification induces changes in virus–host relationships in Mediterranean benthic ecosystems. Microorganisms 2021, 9, 769. [Google Scholar] [CrossRef] [PubMed]
- Danovaro, R.; Gambi, C.; Mirto, S.; Sandulli, R.; Ceccherelli, V.U. Meiofauna. In Mediterranean Marine Benthos: A Manual of Methods for Its Sampling and Study; Gambi, M.C., Dappiano, M., Eds.; Biologia Marina Mediterranea; SIBM: Genova, Italy, 2004; Volume 11, pp. 55–97. [Google Scholar]
- Feller, R.J.; Warwick, R.M. Energetics. In Introduction to the Study of Meiofauna; Higgins, R.P., Thiel, H., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1988; pp. 181–196. [Google Scholar]
- Wisser, W. Die Beziehungen zwischen Mundhohlengestalt, Ernahrungsweise und Vorkommen beifreilebenden marinen Nematoden. Ark. Zool. 1953, 4, 439–484. [Google Scholar]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA+ for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, UK, 2008. [Google Scholar]
- Anderson, M.J.; Willis, T.J. Canonical analysis of principal coordinates: A useful method of constrained ordination for ecology. Ecology 2003, 84, 511–525. [Google Scholar] [CrossRef]
- Bongers, T.; Ferris, H. Nematode community structure as a bioindicator in environmental monitoring. Trends Ecol. Evol. 1999, 14, 224–228. [Google Scholar] [CrossRef]
- McArdle, B.H.; Anderson, M.J. Fitting multivariate models to community data: A comment on distance-based redundancy analysis. Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. Getting Started with PRIMER v7; PRIMER-E: Plymouth, UK, 2015; Available online: https://www.primer-e.com/ (accessed on 20 December 2021).
- Hall-Spencer, J.M.; Harvey, B.P. Ocean acidification impacts on coastal ecosystem services due to habitat degradation. Emerg. Top. Life Sci. 2019, 3, 197–206. [Google Scholar] [CrossRef] [Green Version]
- Kroeker, K.; Kordas, R.; Crim, R.; Hendriks, I.; Ramajo, L.; Singh, G.; Duarte, C.; Gattuso, J.-P. Impacts of ocean acidification on marine organisms: Quantifying sensitivities and interaction with warming. Glob. Chang. Biol. 2013, 19, 1884–1896. [Google Scholar] [CrossRef] [Green Version]
- Garilli, V.; Rodolfo-Metalpa, R.; Scuderi, D.; Brusca, L.; Parrinello, D.; Rastrick, S.P.S.; Foggo, A.; Twichett, R.J.; Hall-Spencer, J.M.; Milazzo, M. Physiological advantages of dwarfing in surviving extinctions in high-CO2 oceans. Nat. Clim. Chang. 2015, 5, 678–682. [Google Scholar] [CrossRef]
- Harvey, B.P.; Agostini, S.; Wada, S.; Inaba, K.; Hall-Spencer, J.M. Dissolution: The Achilles’ heel of the triton shell in an acidifying ocean. Front. Mar. Sci. 2018, 5, 371. [Google Scholar] [CrossRef]
- Barry, J.P.; Buck, K.R.; Lovera, C.F.; Kuhnz, L.; Whaling, P.J.; Peltzer, E.T.; Walz, P.; Brewer, P.G. Effects of Direct Ocean CO2 Injection on Deep-Sea Meiofauna. J. Oceanogr. 2004, 60, 759–766. [Google Scholar] [CrossRef]
- Hale, R.; Calosi, P.; Mieszkowska, N.; Widdicombe, S.; McNeill, L. Predicted levels of future ocean acidification and temperature rise could alter community structure and biodiversity in marine benthic communities. Oikos 2011, 120, 661–674. [Google Scholar] [CrossRef]
- Kroeker, K.J.; Micheli, F.; Gambi, M.C. Ocean acidification causes ecosystem shifts via altered competitive interactions. Nat. Clim. Chang. 2013, 3, 156–159. [Google Scholar] [CrossRef]
- Sarmento, V.; Souza, T.; Esteves, A.; Santos, P. Effects of seawater acidification on a coral reef meiofauna community. Coral Reefs 2015, 34, 955–966. [Google Scholar] [CrossRef]
- Schade, H.; Mevenkamp, L.; Guilini, K.; Meyer, S.; Gorb, S.N.; Abele, D.; Vanreusel, A.; Melzner, F. Simulated leakage of high pCO2 water negatively impacts bivalve dominated infaunal communities from the Western Baltic Sea. Sci. Rep. 2016, 6, 31447. [Google Scholar] [CrossRef] [Green Version]
- Rassmann, J.; Lansard, B.; Gazeau, F.; Guidi-Guilvard, L.; Pozzato, L.; Alliouane, S.; Grenz, C.; Rabouille, C. Impact of ocean acidification on the biogeochemistry and meiofaunal assemblage of carbonate-rich sediments: Results from core incubations (Bay of Villefranche, NW Mediterranean Sea). Mar. Chem. 2018, 203, 102–119. [Google Scholar] [CrossRef]
- Dashfield, S.; Somerfield, P.; Widdicombe, S.; Austen, M. Impacts of ocean acidification and burrowing urchins on within-sediment pH profiles and subtidal nematode communities. J. Exp. Mar. Biol. Ecol. 2008, 365, 46–52. [Google Scholar] [CrossRef]
- Widdicombe, S.; Dashfield, S.L.; McNeill, C.L.; Needham, H.R.; Beesley, A.; McEvoy, A.; Øxnevad, S.; Clarke, K.R.; Berge, J.A. Effects of CO2 induced seawater acidification on infaunal diversity and sediment nutrient fluxes. Mar. Ecol. Prog. Ser. 2009, 379, 59–75. [Google Scholar] [CrossRef] [Green Version]
- Fricke, H.; Giere, O.; Stetter, K.; Alfredsson, G.A.; Kristjansson, J.K.; Stoffers, P.; Svavarsson, J. Hydrothermal vent communities at the shallow subpolar Mid-Atlantic ridge. Mar. Biol. 1989, 102, 425–429. [Google Scholar] [CrossRef]
- Kamenev, G.M.; Fadeev, V.I.; Selin, N.I.; Tarasov, V.G.; Malakhov, V.V. Composition and distribution of macro- and meiobenthos around sublittoral hydrothermal vents in the Bay of Plenty, New Zealand. N. Z. J. Mar. Freshw. Res. 1993, 27, 407–418. [Google Scholar] [CrossRef]
- Dando, P.R.; Hughes, J.A.; Thiermann, F. Preliminary observations on biological communities at shallow hydrothermal vents in the Aegean Sea. Geol. Soc. Lond. Spec. Publ. 1995, 87, 303–317. [Google Scholar] [CrossRef]
- Dando, P.R.; Hughes, J.A.; Leahy, Y.; Niven, S.J.; Taylor, L.J.; Smith, C. Gas venting rates from submarine hydrothermal areas around the island of Milos, Hellenic Volcanic Arc. Cont. Shelf Res. 1995, 15, 913–929. [Google Scholar] [CrossRef]
- Thiermann, F.; Akoumianaki, I.; Hughes, J.A.; Giere, O. Benthic fauna of a shallow-water gaseohydrothermal vent area in the Aegean Sea (Milos, Greece). Mar. Biol. 1997, 128, 149–159. [Google Scholar] [CrossRef]
- Bianchelli, S.; Gambi, C.; Zeppilli, D.; Danovaro, R. Metazoan meiofauna in deep-sea canyons and adjacent open slopes: A large-scale comparison with focus on the rare taxa. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2010, 57, 420–433. [Google Scholar] [CrossRef]
- Pusceddu, A.; Bianchelli, S.; Gambi, C.; Danovaro, R. Assessment of benthic trophic status of marine coastal ecosystems: Significance of meiofaunal rare taxa. Estuar. Coast. Shelf Sci. 2011, 93, 420–430. [Google Scholar] [CrossRef]
- Vanreusel, A.; De Groote, A.; Gollner, S.; Bright, M. Ecology and Biogeography of Free-Living Nematodes Associated with Chemosynthetic Environments in the Deep Sea: A Review. PLoS ONE 2010, 5, e12449. [Google Scholar] [CrossRef]
- Levin, L.A.; Baco, A.R.; Bowden, D.A.; Colaco, A.; Cordes, E.E.; Cunha, M.R.; Demopoulos, A.W.; Gobin, J.; Grupe, B.M.; Le, J.; et al. Hydrothermal vents and methane seeps: Rethinking the sphere of influence. Front. Mar. Sci. 2016, 3, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Whiteley, N.M. Physiological and ecological responses of crustaceans to ocean acidification. Mar. Ecol. Prog. Ser. 2011, 430, 257–271. [Google Scholar] [CrossRef] [Green Version]
- Cigliano, M.; Gambi, M.C.; Rodolfo-Metalpa, R.; Patti, F.; Hall-Spencer, J. Effects of ocean acidification on invertebrate settlement at volcanic CO2 vents. Mar. Biol. 2010, 157, 2489–2502. [Google Scholar] [CrossRef] [Green Version]
- Byrne, M.; Ho, M.; Wong, E.; Soars, N.A.; Selvakumaraswamy, P.; Shepard-Brennand, H.; Dworjanyn, S.A.; Davis, A.R. Unshelled abalone and corrupted urchins: Development of marine calcifiers in a changing ocean. Proc. R. Soc. B Biol. Sci. 2011, 278, 2376–2383. [Google Scholar] [CrossRef] [PubMed]
- Byrne, M. Global change ecotoxicology: Identification of early life history bottlenecks in marine invertebrates, variable species responses and variable experimental approaches. Mar. Enviromental Res. 2012, 76, 3–15. [Google Scholar] [CrossRef]
- Chan, K.Y.K.; Grünbaum, D.; O’Donnell, M.J. Effects of ocean-acidification-induced morphological changes on larval swimming and feeding. J. Exp. Biol. 2011, 214, 3857–3867. [Google Scholar] [CrossRef] [Green Version]
- Gaylord, B.; Hill, T.M.; Sanford, E.; Lenz, E.A.; Jacobs, L.A.; Sato, K.N.; Russell, A.D.; Hettinger, A. Functional impacts of ocean acidification in an ecologically critical foundation species. J. Exp. Biol. 2011, 214, 2586–2594. [Google Scholar] [CrossRef] [Green Version]
- Michaelidis, B.; Ouzounis, C.; Paleras, A.; Pörtner, H.O. Effects of long-term moderate hypercapnia on acid-base balance and growth rate in marine mussels Mytilus galloprovincialis. Mar. Ecol. Prog. Ser. 2005, 293, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Melzner, F.; Gutowska, M.A.; Langenbuch, M.; Dupont, S.; Lucassen, M.; Thorndyke, M.C.; Bleich, M.; Pörtner, H.O. Physiological basis for high CO2 tolerance in marine ectothermic animals: Pre-adaptation through lifestyle and ontogeny? Biogeosciences 2009, 6, 2313–2331. [Google Scholar] [CrossRef] [Green Version]
- Miles, H.; Widdicombe, S.; Spicer, J.I.; Hall-Spencer, J. Effects of anthropogenic seawater acidification on acid–base balance in the sea urchin Psammechinus miliaris. Mar. Pollut. Bull. 2007, 54, 89–96. [Google Scholar] [CrossRef]
- Melatunan, S.; Calosi, P.; Rundle, S.; Moody, A.; Widdicombe, S. Exposure to elevated temperature and pCO2 reduces respiration rate and energy status in the periwinkle Littorina littorea. Physiol. Biochem. Zool. 2011, 84, 583–594. [Google Scholar] [CrossRef]
- Tomanek, L.; Zuzow, M.J.; Ivanina, A.V.; Beniash, E.; Sokolova, I.M. Proteomic response to elevated PCO2 level in eastern oysters, Crassostrea virginica: Evidence for oxidative stress. J. Exp. Biol. 2011, 214, 1836–1844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hylleberg, J. On the ecology of the Sipunculian Phascolion strombi (Montagu). In Proceedings of the International Symposium on the Biology of Sipuncula and Echiura I, Kotor, Montenegro, 18–25 June 1970. [Google Scholar]
- Bergamo, G.; Carrerette, O.; Zanol, J.; Nogueira, J. Species of Eunicidae (Annelida, Errantia, Eunicida) from rhodolith beds off the states of Paraíba and Pernambuco, northeastern Brazilian coast. Zootaxa 2018, 4521, 376. [Google Scholar] [CrossRef] [PubMed]
- Caine, E.A. First case of caprellid amphipod-hydrozoan mutualism. J. Crustacean Biol. 1998, 18, 317–320. [Google Scholar] [CrossRef] [Green Version]
- Levin, L.A.; James, D.W.; Martin, C.M.; Rathburn, A.E.; Harris, L.H.; Michener, R. Do methane seeps support distinct infaunal assemblages? Observations on community structure and nutrition from the northern California slope and shelf. Mar. Ecol. Prog. Ser. 2000, 208, 21–39. [Google Scholar] [CrossRef] [Green Version]
- Chapman, P.M.; Farrell, M.A.; Brinkhurst, R.O. Relative tolerances of selected aquatic oligochaetes to individual pollutants and environmental factors. Aquat. Toxicol. 1982, 2, 47–67. [Google Scholar] [CrossRef]
- Donnarumma, L.; Appolloni, L.; Chianese, E.; Bruno, R.; Baldrighi, E.; Guglielmo, R.; Russo, G.; Zeppilli, D.; Sandulli, R. Environmental and benthic community patterns of the shallow hydrothermal area of Secca delle Fumose (Baia, Naples, Italy). Front. Mar. Sci. 2019, 6, 685. [Google Scholar] [CrossRef] [Green Version]
- Kaartvedt, S.; Van Dover, C.L.; Mullineaux, L.S.; Wiebe, P.H.; Bollens, S.M. Amphipods on a deep-sea hydrothermal treadmill. Deep. Sea Res. Part I Oceanogr. Res. Pap. 1994, 41, 179–195. [Google Scholar] [CrossRef]
- Gambi, M.C.; Musco, L.; Giangrande, A.; Badalamenti, F.; Micheli, F.; Kroeker, K. Distribution and functional traits of polychaetes in a CO2 vent system: Winners and losers among closely related species. Mar. Ecol. Prog. Ser. 2016, 550, 121–134. [Google Scholar] [CrossRef] [Green Version]
- Vizzini, S.; Martínez-Crego, B.; Andolina, C.; Massa-Gallucci, A.; Connell, S.D.; Gambi, M.C. Ocean acidification as a driver of community simplification via the collapse of higher-order and rise of lower-order consumers. Sci. Rep. 2017, 7, 4018. [Google Scholar] [CrossRef] [Green Version]
- Appolloni, L.; Zeppilli, D.; Donnarumma, L.; Baldrighi, E.; Chianese, E.; Russo, G.F.; Sandulli, R. The acidification of sea water affects the beta-diversity of benthic communities in a shallow hydrothermal vent in a protected marine area of the Mediterranean. Diversity 2020, 12, 464. [Google Scholar] [CrossRef]
- Linares, C.; Vidal, M.; Canals, M.; Kersting, D.K.; Amblas, D.; Aspillaga, E.; Cebrián, E.; Delgado-Huertas, A.; Díaz, D.; Garrabou, J.; et al. Persistent natural acidification drives major distribution shifts in marine benthic ecosystems. Proc. R. Soc. B Biol. Sci. 2015, 282, 20150587. [Google Scholar] [CrossRef] [PubMed]
- Cohen, A.; Michael, H. Why corals care about ocean acidification: Uncovering the mechanism. Oceanography 2009, 22, 118–127. [Google Scholar] [CrossRef]
- Holcomb, M.; Cohen, A.; McCorkle, D. A gender bias in the calcification response to ocean acidification. Biogeosci. Discuss. 2011, 8, 8485–8513. [Google Scholar] [CrossRef]
- Melzner, F.; Stange, P.; Trübenbach, K.; Thomsen, J.; Casties, I.; Panknin, U.; Gorb, S.N.; Gutowska, M.A. Food supply and seawater pCO2 impact calcification and internal shell dissolution in the blue mussel Mytilus edulis. PLoS ONE 2011, 6, e24223. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, J.; Casties, I.; Pansch, C.; Körtzinger, A.; Melzner, F. Food availability outweighs ocean acidification effects in juvenile Mytilus edulis: Laboratory and field experiments. Glob. Chang. Biol. 2013, 19, 1017–1027. [Google Scholar] [CrossRef] [Green Version]
- Falkenberg, L.; Russell, B.; Connell, S. Future herbivory: The indirect effects of enriched CO2 may rival its direct effects. Mar. Ecol. Prog. Ser. 2013, 492, 85–95. [Google Scholar] [CrossRef] [Green Version]
- Ghedini, G.; Connell, S. Organismal homeostasis buffers the effects of abiotic change on community dynamics. Ecology 2016, 97, 2671–2679. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) | Abundance | Biomass | ||||
| Source | df | MS | F | MS | F | |
| Vents | 1 | 9.05 | 22.87 *** | 0.07 | 0.18 ns | |
| Depth | 1 | 0.45 | 1.15 ns | 4.42 | 11.15 ** | |
| Vents × Depth | 1 | 0.01 | 0.02 ns | 0.17 | 0.44 ns | |
| Residuals | 19 | 0.4 | 0.4 | |||
| Total | 22 | |||||
| (b) | Within level ‘shallow’ of factor ‘Depth’ for abundance | |||||
| Groups | t | |||||
| Active vs. Inactive | 3.76 * | |||||
| Within level ‘deep’ of factor ‘Depth’ for abundance | ||||||
| Groups | t | |||||
| Active vs. Inactive | 3.22 * | |||||
| (a) | Abundance | Biomass | ||||
| Source | df | MS | F | MS | F | |
| Vents | 1 | 262,080 | 6.25 * | 276.88 | 7.70 * | |
| Depth | 1 | 206,290 | 4.92 * | 80.89 | 2.25 ns | |
| Vents × Depth | 1 | 7410.1 | 0.18 ns | 74.07 | 2.06 ns | |
| Residuals | 18 | 41,912 | 35.94 | |||
| Total | 21 | |||||
| (b) | Within level ‘shallow’ of factor ‘depth’ | |||||
| Groups | t | t | ||||
| Active vs. Inactive | 1.24 ns | 1.79 * | ||||
| Within level ‘deep’ of factor ‘depth’ | ||||||
| Groups | t | t | ||||
| Active vs. Inactive | 5.38 ** | 3.40 ** | ||||
| (a) | Meiofaunal Richness of Higher Taxa | Macrofaunal Species Richness | |||||
| Source | df | MS | F | df | MS | F | |
| Vents | 1 | 19.79 | 2.60 ns | 1 | 712.5 | 45.18 *** | |
| Depth | 1 | 73.44 | 9.63 ** | 1 | 89.48 | 5.67 * | |
| Vents × Depth | 1 | 8.7 | 1.14 ns | 1 | 1.97 | 0.12 ns | |
| Residuals | 19 | 7.63 | 18 | 15.77 | |||
| Total | 22 | 21 | |||||
| (b) | Within level ‘shallow’ of factor ‘depth’ | ||||||
| Groups | t | ||||||
| Active vs. Inactive | _ | 3.17 * | |||||
| Within level ‘deep’ of factor ‘depth’ | |||||||
| Groups | t | ||||||
| Active vs. Inactive | _ | 9.48 ** | |||||
| (a) | Meiofauna | Macrofauna | |||||
| Source | df | MS | Pseudo-F | df | MS | Pseudo-F | |
| Vents | 1 | 3819.8 | 5.95 *** | 1 | 10,354 | 5.28 *** | |
| Depth | 1 | 5665.4 | 8.83 *** | 1 | 6838 | 3.49 *** | |
| Vents × Depth | 1 | 602.05 | 0.94 ns | 1 | 5186.3 | 2.64 ** | |
| Residuals | 19 | 641.78 | 18 | 1962.1 | |||
| Total | 22 | 21 | |||||
| (b) | Within level ‘shallow’ of factor ‘depth’ | ||||||
| Groups | t | t | |||||
| Inactive vs. Active | 2.49 ** | 2.16 ** | |||||
| Within level ‘deep’ of factor ‘depth’ | |||||||
| Groups | t | t | |||||
| Inactive vs. Active | 1.41 ns | 2.06 ** | |||||
| Meiofauna | ||||||
| Variable | Adj R2 | Pseudo-F | p | Prop. | Cumul. | res.df |
| PRT | 0.13 | 4.76 | 0.004 | 0.17 | 0.17 | 24 |
| TPH | 0.24 | 4.54 | 0.005 | 0.14 | 0.30 | 23 |
| LIP | 0.25 | 1.20 | 0.321 | 0.04 | 0.34 | 22 |
| %PRT | 0.28 | 1.99 | 0.088 | 0.06 | 0.40 | 21 |
| Macrofauna | ||||||
| Variable | Adj R2 | Pseudo-F | p | Prop. | Cumul. | res.df |
| TPH | 0.07 | 2.57 | 0.004 | 0.11 | 0.11 | 21 |
| PRT | 0.11 | 2.02 | 0.023 | 0.08 | 0.19 | 20 |
| %PRT | 0.15 | 1.87 | 0.031 | 0.07 | 0.26 | 19 |
| %CHO | 0.18 | 1.84 | 0.029 | 0.07 | 0.33 | 18 |
| CHO | 0.21 | 1.59 | 0.085 | 0.06 | 0.39 | 17 |
| Nematoda | 0.22 | 1.25 | 0.276 | 0.04 | 0.43 | 16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fanelli, E.; Di Giacomo, S.; Gambi, C.; Bianchelli, S.; Da Ros, Z.; Tangherlini, M.; Andaloro, F.; Romeo, T.; Corinaldesi, C.; Danovaro, R. Effects of Local Acidification on Benthic Communities at Shallow Hydrothermal Vents of the Aeolian Islands (Southern Tyrrhenian, Mediterranean Sea). Biology 2022, 11, 321. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11020321
Fanelli E, Di Giacomo S, Gambi C, Bianchelli S, Da Ros Z, Tangherlini M, Andaloro F, Romeo T, Corinaldesi C, Danovaro R. Effects of Local Acidification on Benthic Communities at Shallow Hydrothermal Vents of the Aeolian Islands (Southern Tyrrhenian, Mediterranean Sea). Biology. 2022; 11(2):321. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11020321
Chicago/Turabian StyleFanelli, Emanuela, Simone Di Giacomo, Cristina Gambi, Silvia Bianchelli, Zaira Da Ros, Michael Tangherlini, Franco Andaloro, Teresa Romeo, Cinzia Corinaldesi, and Roberto Danovaro. 2022. "Effects of Local Acidification on Benthic Communities at Shallow Hydrothermal Vents of the Aeolian Islands (Southern Tyrrhenian, Mediterranean Sea)" Biology 11, no. 2: 321. https://0-doi-org.brum.beds.ac.uk/10.3390/biology11020321