
Kaempferitrin-Treated HepG2 Differentially Expressed Exosomal Markers and Affect Extracellular Vesicle Sizes in the Secretome

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture, Drug Treatment, and Sample Collection for Proteomics and Western Blotting
2.2. Quantitative Proteomics Using Dimethyl Labelling
2.3. Data Analyses
2.4. Atomic Force Microscopy (AFM)
3. Results
3.1. Proteomics
3.1.1. First Round Selection
3.1.2. Second Round Selection
3.1.3. Double selection
3.2. Identification of Exosomal Markers
3.3. Western Blots
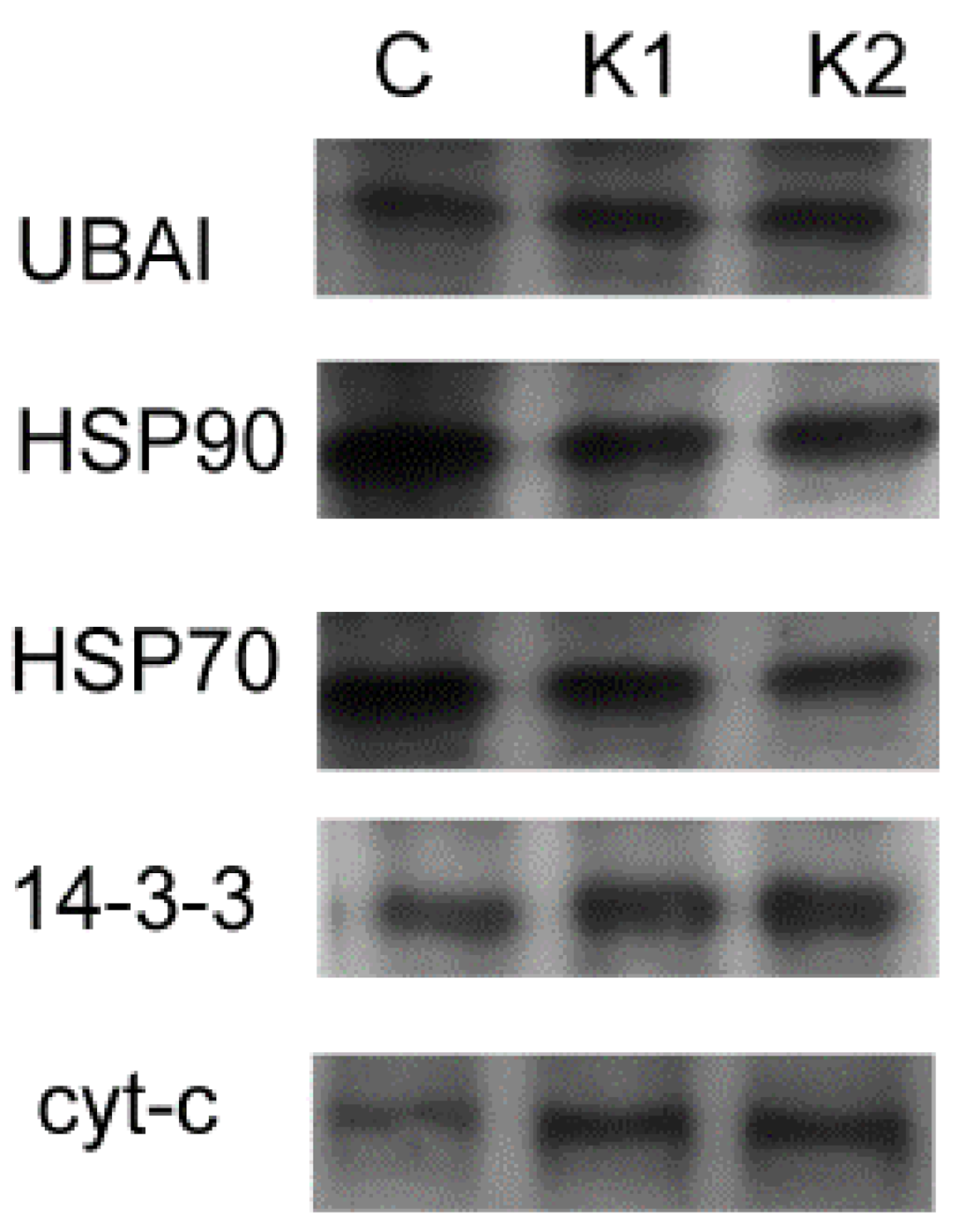
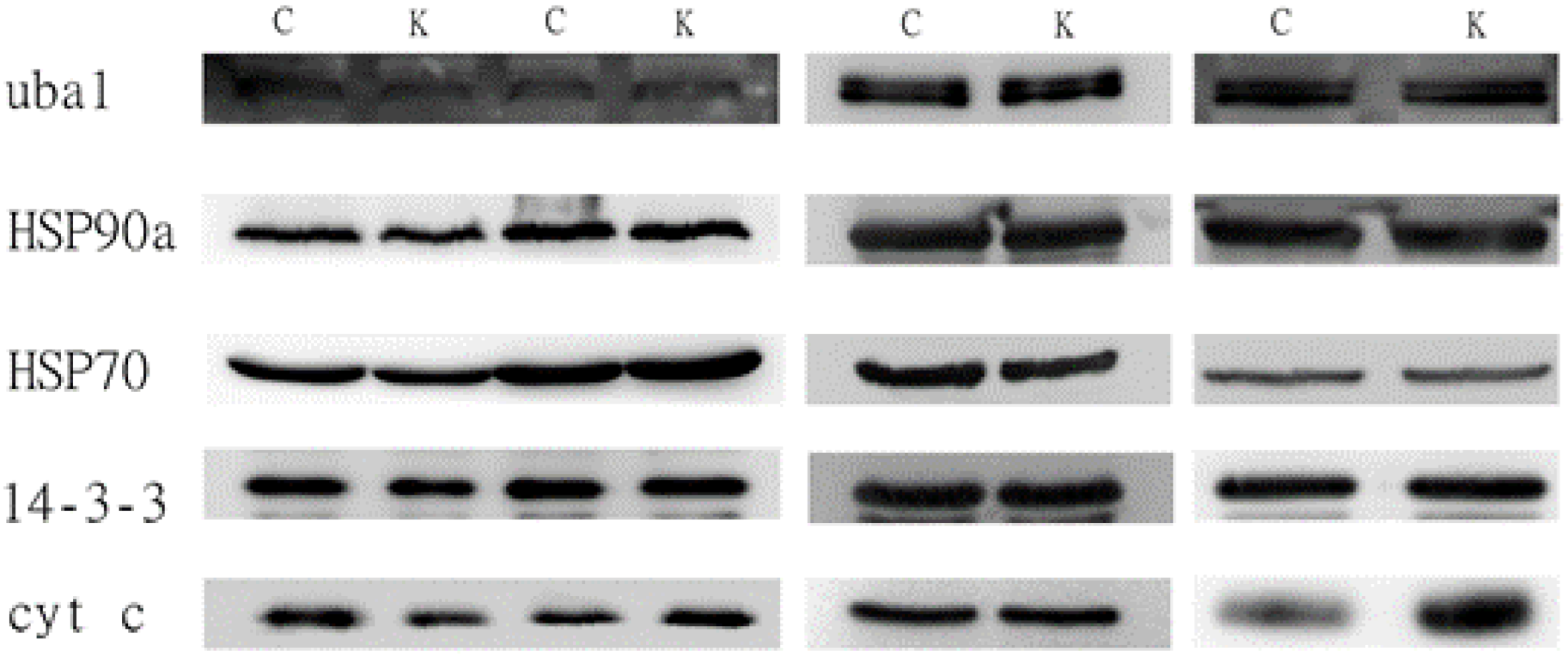
3.3.1. The 0.22-μm Filter
3.3.2. The 0.45-μm Filter
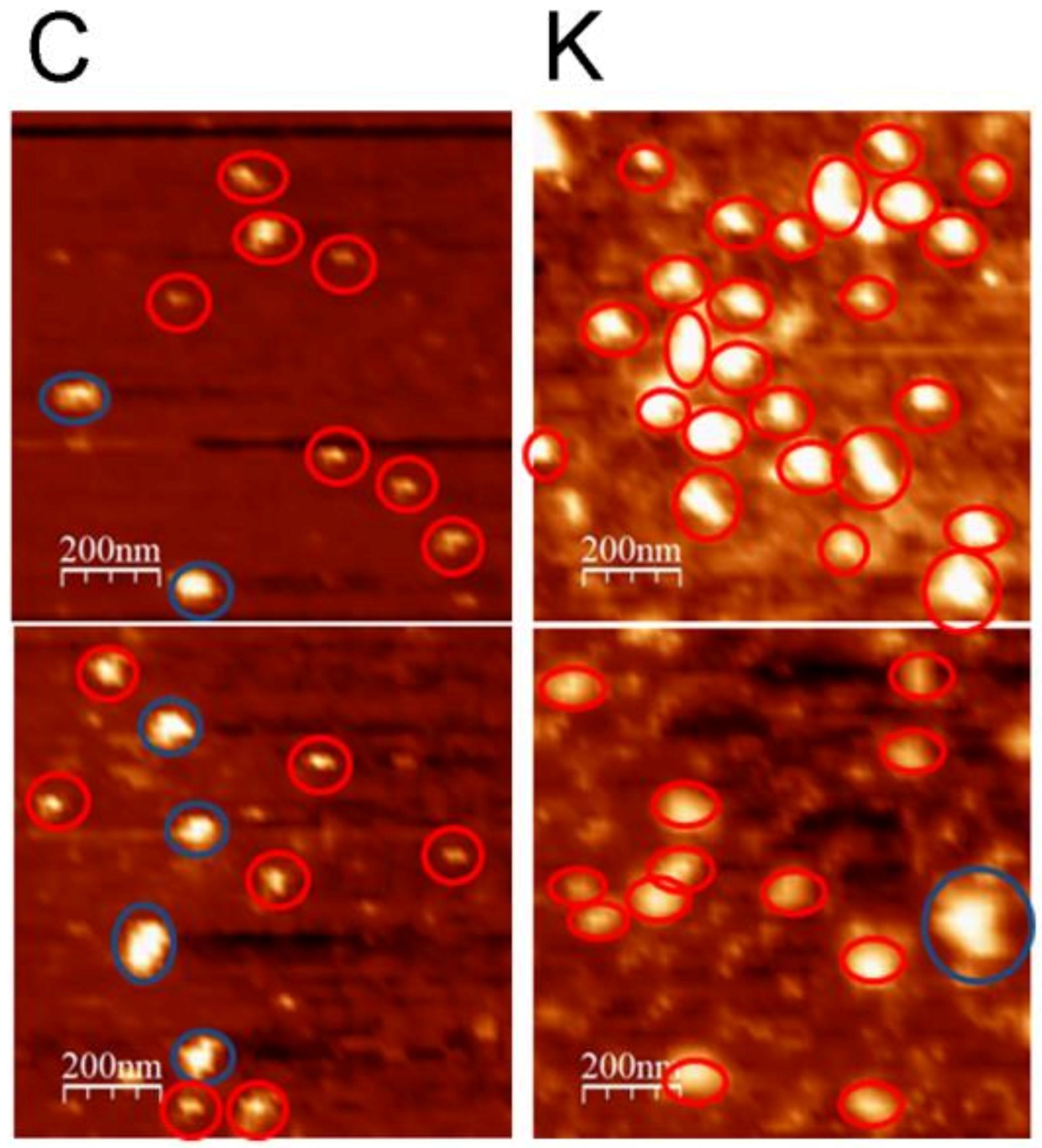
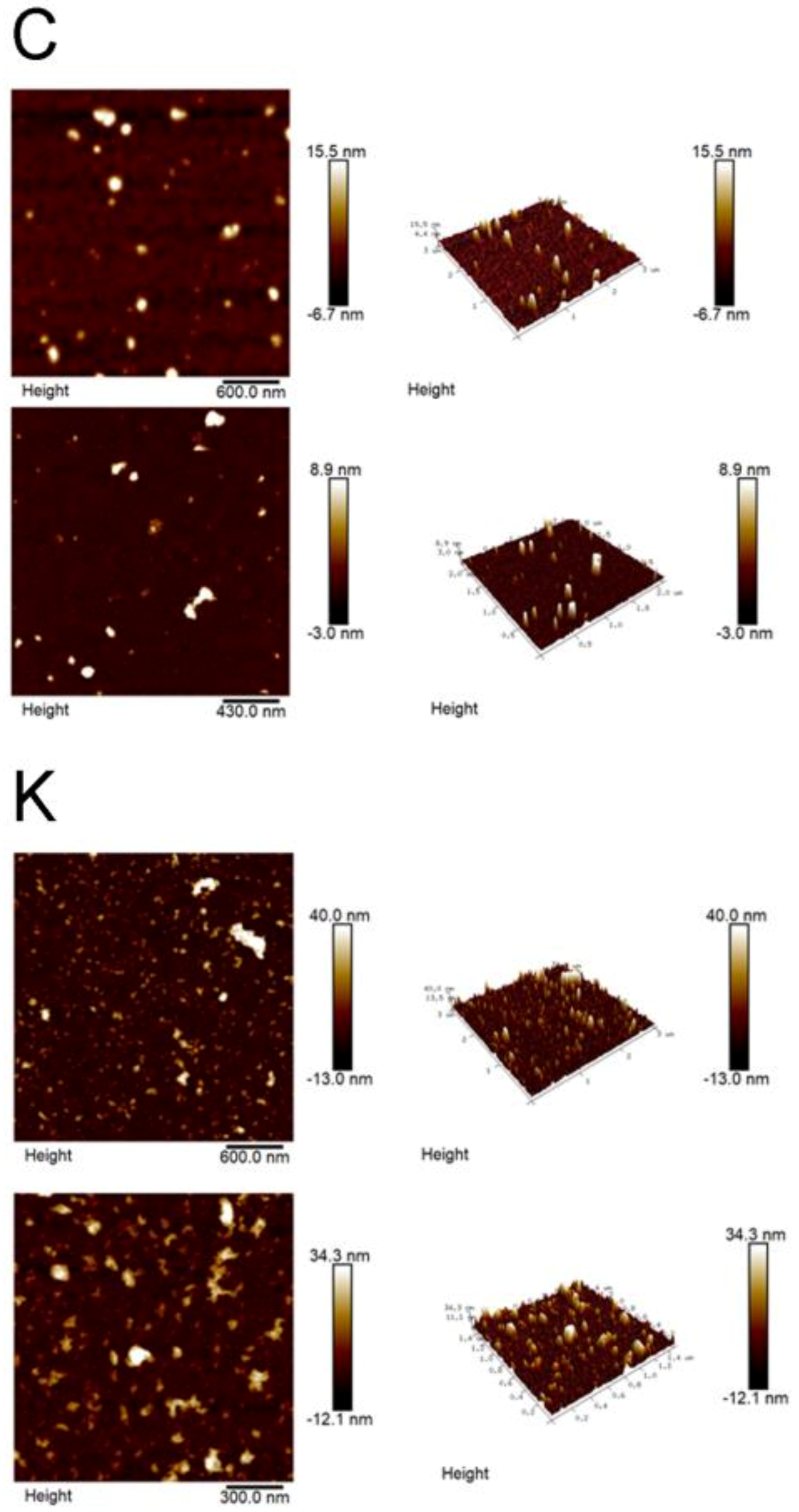
3.4. AFM Studies
3.4.1. Nanoview1000
3.4.2. Brucker Dimension Icon
4. Discussion
4.1. Regulation of Exosomal Markers
4.2. The Kaempferitrin Treatment Results in Alteration to the EVs
4.3. Western Blot Result for Conditioned Medium
4.4. Why Are Kaempferitrin Treated EVs Larger?
4.5. Regulation of Lipid Metabolism
4.6. Proteomic as a Tool for Study Exososomal Regulation
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fang, S.-H.; Rao, Y.K.; Tzeng, Y.-M. Inhibitory effects of flavonol glycosides from Cinnamomum osmophloeum on inflammatory mediators in LPS/IFN-γ-activated murine macrophages. Bioorg. Med. Chem. 2005, 13, 2381–2388. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Liang, T.; Xu, L.; Li, Y.; Zhang, S.; Duan, X. Protective effect of cinnamon polyphenols against STZ-diabetic mice fed high-sugar, high-fat diet and its underlying mechanism. Food Chem. Toxicol. 2013, 51, 419–425. [Google Scholar] [CrossRef] [PubMed]
- De Sousa, E.; Zanatta, L.; Seifriz, I.; Creczynski-Pasa, T.B.; Pizzolatti, M.G.; Szpoganicz, B.; Silva, F.R. Hypoglycemic effect and antioxidant potential of kaempferol-3,7-O-(alpha)-dirhamnoside from Bauhinia forficata leaves. J. Nat. Prod. 2004, 67, 829–832. [Google Scholar] [CrossRef] [PubMed]
- Lino Cde, S.; Diogenes, J.P.; Pereira, B.A.; Faria, R.A.; Neto, M.A.; Alves, R.S.; de Queiroz, M.G.; de Sousa, F.C.; Viana, G.S. Antidiabetic activity of Bauhinia forficata extracts in alloxan-diabetic rats. Biol. Pharm. Bull. 2004, 27, 125–127. [Google Scholar] [CrossRef] [Green Version]
- Lin, G.-M.; Chen, Y.-H.; Yen, P.-L.; Chang, S.-T. Antihyperglycemic and antioxidant activities of twig extract from Cinnamomum osmophloeum. J. Tradit. Complement. Med. 2016, 6, 281–288. [Google Scholar] [CrossRef] [Green Version]
- Ku, W.-C.; Chang, Y.-L.; Wu, S.-F.; Shih, H.-N.; Tzeng, Y.-M.; Kuo, H.-R.; Chang, K.-M.; Agrawal, D.C.; Liu, B.-L.; Chang, C.-A. A comparative proteomic study of secretomes in kaempferitrin-treated CTX TNA2 astrocytic cells. Phytomedicine 2017, 36, 137–144. [Google Scholar] [CrossRef]
- Villarreal, L.; Méndez, O.; Salvans, C.; Gregori, J.; Baselga, J.; Villanueva, J. Unconventional secretion is a major contributor of cancer cell line secretomes. Mol. Cell. Proteom. 2013, 12, 1046–1060. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, P.; Leray, V.; Diez, M.; Serisier, S.; Bloc’h, J.L.; Siliart, B.; Dumon, H. Liver lipid metabolism. J. Anim. Physiol. Anim. Nutr. 2008, 92, 272–283. [Google Scholar] [CrossRef]
- Lin, T.Y.; Liao, J.W.; Chang, S.T.; Wang, S.Y. Antidyslipidemic activity of hot-water extracts from leaves of Cinnamomum osmophloeum Kaneh. Phytother. Res. 2011, 25, 1317–1322. [Google Scholar] [CrossRef]
- Boersema, P.J.; Raijmakers, R.; Lemeer, S.; Mohammed, S.; Heck, A.J. Multiplex peptide stable isotope dimethyl labeling for quantitative proteomics. Nat. Protoc. 2009, 4, 484–494. [Google Scholar] [CrossRef]
- Rappsilber, J.; Mann, M.; Ishihama, Y. Protocol for micro-purification, enrichment, pre-fractionation and storage of peptides for proteomics using StageTips. Nat. Protoc. 2007, 2, 1896–1906. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized ppb-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef] [PubMed]
- Abudoureyimu, M.; Zhou, H.; Zhi, Y.; Wang, T.; Feng, B.; Wang, R.; Chu, X. Recent progress in the emerging role of exosome in hepatocellular carcinoma. Cell Prolif. 2018, 52, e12541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Numair, K.S.; Chandramohan, G.; Veeramani, C.; Alsaif, M.A. Ameliorative effect of kaempferol, a flavonoid, on oxidative stress in streptozotocin-induced diabetic rats. Redox Rep. 2015, 20, 198–209. [Google Scholar] [CrossRef]
- Tzeng, Y.-M.; Chen, K.; Rao, Y.K.; Lee, M.-J. Kaempferitrin activates the insulin signaling pathway and stimulates secretion of adiponectin in 3T3-L1 adipocytes. Eur. J. Pharmacol. 2009, 607, 27–34. [Google Scholar] [CrossRef]
- Wolska, A.; Dunbar, R.L.; Freeman, L.A.; Ueda, M.; Amar, M.J.; Sviridov, D.O.; Remaley, A.T. Apolipoprotein C-II: New findings related to genetics, biochemistry, and role in triglyceride metabolism. Atherosclerosis 2017, 267, 49–60. [Google Scholar] [CrossRef]
- Kei, A.A.; Filippatos, T.D.; Tsimihodimos, V.; Elisaf, M.S. A review of the role of apolipoprotein C-II in lipoprotein metabolism and cardiovascular disease. Metabolism 2012, 61, 906–921. [Google Scholar] [CrossRef]
- Yalcin, E.B.; More, V.; Neira, K.L.; Lu, Z.J.; Cherrington, N.J.; Slitt, A.L.; King, R.S. Downregulation of sulfotransferase expression and activity in diseased human livers. Drug Metab. Dispos. 2013, 41, 1642–1650. [Google Scholar] [CrossRef] [Green Version]
- Cazarolli, L.H.; Zanatta, L.; Jorge, A.P.; de Sousa, E.; Horst, H.; Woehl, V.M.; Pizzolatti, M.G.; Szpoganicz, B.; Silva, F.R. Follow-up studies on glycosylated flavonoids and their complexes with vanadium: Their anti-hyperglycemic potential role in diabetes. Chem. Biol. Interact. 2006, 163, 177–191. [Google Scholar] [CrossRef]
- Jorge, A.P.; Horst, H.; de Sousa, E.; Pizzolatti, M.G.; Silva, F.R. Insulinomimetic effects of kaempferitrin on glycaemia and on 14C-glucose uptake in rat soleus muscle. Chem. Biol. Interact. 2004, 149, 89–96. [Google Scholar] [CrossRef]
- Chua, M.T.; Tung, Y.T.; Chang, S.T. Antioxidant activities of ethanolic extracts from the twigs of Cinnamomum osmophloeum. Bioresour. Technol. 2008, 99, 1918–1925. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.-J.; Rao, Y.K.; Chen, K.; Lee, Y.-C.; Tzeng, Y.-M. Effect of flavonol glycosides from Cinnamomum osmophloeum leaves on adiponectin secretion and phosphorylation of insulin receptor-β in 3T3-L1 adipocytes. J. Ethnopharmacol. 2009, 126, 79–85. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Majority Protein IDs | Protein Names | Gene Names | Peptides |
|---|---|---|---|
| Q8NCF0; A5D8T8; Q6UXF7 | C-type lectin domain family 18 member C; C-type lectin domain family 18 member A; C-type lectin domain family 18 member B | CLEC18C; CLEC18A; CLEC18B | 1 |
| A6NC57 | Ankyrin repeat domain-containing protein 62 | ANKRD62 | 1 |
| O95433 | Activator of 90 kDa heat shock protein ATPase homolog 1 | AHSA1 | 3 |
| P02655 | Apolipoprotein C-II; Proapolipoprotein C-II | APOC2 | 2 |
| P06748 | Nucleophosmin | NPM1 | 8 |
| P07900 | Heat shock protein HSP 90-α | HSP90AA1 | 27 |
| P08238 | Heat shock protein HSP 90-β | HSP90AB1 | 29 |
| P08243 | Asparagine synthetase [glutamine-hydrolyzing] | ASNS | 1 |
| P0DP25; P0DP24; P0DP23 | Calmodulin-3, Calmodulin-2, Calmodulin-1 | CALM3, CALM2, CALM1 | 7 |
| P13473 | Lysosome-associated membrane glycoprotein 2 | LAMP2 | 1 |
| P14174 | Macrophage migration inhibitory factor | MIF | 1 |
| P14868 | Aspartate-tRNA ligase, cytoplasmic | DARS | 5 |
| P16422 | Epithelial cell adhesion molecule | EPCAM | 2 |
| P22314 | Ubiquitin-like modifier-activating enzyme 1 | UBA1 | 12 |
| P28799 | Granulins; Acrogranin; Paragranulin; Granulin-1; Granulin-2; Granulin-3; Granulin-4; Granulin-5; Granulin-6; Granulin-7 | GRN | 8 |
| P31946 | 14-3-3 protein β/α;14-3-3 protein β/α, N-terminally processed | YWHAB | 6 |
| P39060 | Collagen α-1(XVIII) chain; Endostatin | COL18A1 | 1 |
| P50897 | Palmitoyl-protein thioesterase 1 | PPT1 | 5 |
| P51857 | 3-oxo-5-β-steroid 4-dehydrogenase | AKR1D1 | 1 |
| P62258 | 14-3-3 protein epsilon | YWHAE | 12 |
| P63104 | 14-3-3 protein zeta/delta | YWHAZ | 7 |
| P68402 | Platelet-activating factor acetylhydrolase IB subunit β | PAFAH1B2 | 1 |
| P99999 | Cytochrome c | CYCS | 4 |
| Q06520 | Bile salt sulfotransferase | SULT2A1 | 3 |
| Q07955 | Serine/arginine-rich splicing factor 1 | SRSF1 | 3 |
| Q16576; Q09028 | Histone-binding protein RBBP7; Histone-binding protein RBBP4 | RBBP7; RBBP4 | 2 |
| Q58FF8 | Putative heat shock protein HSP 90-β 2 | HSP90AB2P | 6 |
| Q8TDY2 | RB1-inducible coiled-coil protein 1 | RB1CC1 | 1 |
| Q92688 | Acidic leucine-rich nuclear phosphoprotein 32 family member B | ANP32B | 3 |
| Q92896 | Golgi apparatus protein 1 | GLG1 | 1 |
| Q969E1 | Liver-expressed antimicrobial peptide 2 | LEAP2 | 1 |
| Q99832 | T-complex protein 1 subunit eta | CCT7 | 3 |
| Majority Protein IDs | Protein Names | Gene Names | Peptides |
|---|---|---|---|
| A6NC57 | Ankyrin repeat domain-containing protein 62 | ANKRD62 | 1 |
| P02655 | Apolipoprotein C-II; Proapolipoprotein C-II | APOC2 | 2 |
| P07900 | Heat shock protein HSP 90-α | HSP90AA1 | 27 |
| P08238 | Heat shock protein HSP 90-β | HSP90AB1 | 29 |
| P08833 | Insulin-like growth factor-binding protein 1 | IGFBP1 | 12 |
| P0DP25; P0DP24; P0DP23 | 7 | ||
| P14174 | Macrophage migration inhibitory factor | MIF | 1 |
| P22314 | Ubiquitin-like modifier-activating enzyme 1 | UBA1 | 12 |
| P26641 | Elongation factor 1-γ | EEF1G | 6 |
| P28799 | Granulins; Acrogranin; Paragranulin; Granulin-1; Granulin-2; Granulin-3; Granulin-4; Granulin-5; Granulin-6; Granulin-7 | GRN | 8 |
| P39060 | Collagen α-1(XVIII) chain; Endostatin | COL18A1 | 1 |
| P51857 | 3-oxo-5-β-steroid 4-dehydrogenase | AKR1D1 | 1 |
| P62258 | 14-3-3 protein epsilon | YWHAE | 12 |
| P63104 | 14-3-3 protein zeta/delta | YWHAZ | 7 |
| P68402 | Platelet-activating factor acetylhydrolase IB subunit β | PAFAH1B2 | 1 |
| P84103 | Serine/arginine-rich splicing factor 3 | SRSF3 | 2 |
| P99999 | Cytochrome c | CYCS | 4 |
| Q06033 | Inter-α-trypsin inhibitor heavy chain H3 | ITIH3 | 7 |
| Q06520 | Bile salt sulfotransferase | SULT2A1 | 3 |
| Q07955 | Serine/arginine-rich splicing factor 1 | SRSF1 | 3 |
| Q16576; Q09028 | Histone-binding protein RBBP7; Histone-binding protein RBBP4 | RBBP7; RBBP4 | 2 |
| Q58FF8 | Putative heat shock protein HSP 90-β 2 | HSP90AB2P | 6 |
| Q5R3I4 | Tetratricopeptide repeat protein 38 | TTC38 | 3 |
| Q6UW63 | KDEL motif-containing protein 1 | KDELC1 | 3 |
| Q92563 | Testican-2 | SPOCK2 | 9 |
| Q92688 | Acidic leucine-rich nuclear phosphoprotein 32 family member B | ANP32B | 3 |
| Q96FW1 | Ubiquitin thioesterase OTUB1 | OTUB1 | 3 |
| Q9Y4L1 | Hypoxia up-regulated protein 1 | HYOU1 | 4 |
| Majority Protein IDs | Protein Names | Gene Names | Peptides |
|---|---|---|---|
| A6NC57 | Ankyrin repeat domain-containing protein 62 | ANKRD62 | 1 |
| P02655 | Apolipoprotein C-II; Proapolipoprotein C-II | APOC2 | 2 |
| P07900 | Heat shock protein HSP 90-α | HSP90AA1 | 27 |
| P08238 | Heat shock protein HSP 90-β | HSP90AB1 | 29 |
| P14174 | Macrophage migration inhibitory factor | MIF | 1 |
| P22314 | Ubiquitin-like modifier-activating enzyme 1 | UBA1 | 12 |
| P28799 | Granulins; Acrogranin; Paragranulin; Granulin-1; Granulin-2; Granulin-3; Granulin-4; Granulin-5; Granulin-6; Granulin-7 | GRN | 8 |
| P39060 | Collagen α-1(XVIII) chain; Endostatin | COL18A1 | 1 |
| P51857 | 3-oxo-5-β-steroid 4-dehydrogenase | AKR1D1 | 1 |
| P62258 | 14-3-3 protein epsilon | YWHAE | 12 |
| P63104 | 14-3-3 protein zeta/delta | YWHAZ | 7 |
| P68402 | Platelet-activating factor acetylhydrolase IB subunit β | PAFAH1B2 | 1 |
| P99999 | Cytochrome c | CYCS | 4 |
| Q06520 | Bile salt sulfotransferase | SULT2A1 | 3 |
| Q07955 | Serine/arginine-rich splicing factor 1 | SRSF1 | 3 |
| Q16576; Q09028 | Histone-binding protein RBBP7; Histone-binding protein RBBP4 | RBBP7; RBBP4 | 2 |
| Q58FF8 | Putative heat shock protein HSP 90-β 2 | HSP90AB2P | 6 |
| Q92688 | Acidic leucine-rich nuclear phosphoprotein 32 family member B | ANP32B | 3 |
| Majority Protein IDs | Protein Names | Gene Names |
|---|---|---|
| P07900 | Heat shock protein HSP 90-α | HSP90AA1 |
| P08238 | Heat shock protein HSP 90-β | HSP90AB1 |
| P22314 | Ubiquitin-like modifier-activating enzyme 1 | UBA1 |
| P62258 | 14-3-3 protein epsilon | YWHAE |
| P63104 | 14-3-3 protein zeta/delta | YWHAZ |
| K/C Ratio | Filter (µm) | UBA1 | HSP90A | HSP70 | 14-3-3 Epsilon | Cytochrome c |
|---|---|---|---|---|---|---|
| sample 1 | 0.22 | 1.27 | 0.71 | 0.50 | 1.04 | 1.25 |
| sample 2 | 0.22 | 1.39 | 0.69 | 0.35 | 0.83 | 1.37 |
| sample 3 | 0.45 | 0.96 | 0.87 | 0.87 | 0.99 | 1.10 |
| sample 4 | 0.45 | 0.85 | 0.94 | 0.99 | 1.00 | 1.77 |
| sample 5 | 0.45 | 0.93 | 1.17 | 0.90 | 0.98 | 0.86 |
| sample 6 | 0.45 | 0.91 | 0.99 | 0.92 | 0.99 | 1.24 |
| Control | K 1 | t Test | |
|---|---|---|---|
| EVs number | 33 | 49 | 0.076379451 |
| Average size (nm) | 131.84 | 146.02 |
| Control | K 1 | t Test | |
|---|---|---|---|
| EVs number | 101 | 39 | 0.00005 |
| Average size (nm) | 64.72 | 77.44 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ku, W.-C.; Sridharan, B.; Chen, J.-Y.; Li, J.-Y.; Yang, S.-Y.; Lee, M.-J. Kaempferitrin-Treated HepG2 Differentially Expressed Exosomal Markers and Affect Extracellular Vesicle Sizes in the Secretome. Biomolecules 2021, 11, 187. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11020187
Ku W-C, Sridharan B, Chen J-Y, Li J-Y, Yang S-Y, Lee M-J. Kaempferitrin-Treated HepG2 Differentially Expressed Exosomal Markers and Affect Extracellular Vesicle Sizes in the Secretome. Biomolecules. 2021; 11(2):187. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11020187
Chicago/Turabian StyleKu, Wei-Chi, Badrinathan Sridharan, Jiann-Yeu Chen, Jen-Ying Li, Shu-Yu Yang, and Meng-Jen Lee. 2021. "Kaempferitrin-Treated HepG2 Differentially Expressed Exosomal Markers and Affect Extracellular Vesicle Sizes in the Secretome" Biomolecules 11, no. 2: 187. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11020187