
Xylan Decomposition in Plant Cell Walls as an Inducer of Surfactin Synthesis by Bacillus subtilis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Culture Media, and Culture Conditions
2.2. Determination of Neutral Detergent Fiber (NDF) and Acid Detergent Fiber (ADF) Content in the Fermented Biomass
2.3. Xylanase Activity Assay
2.4. Surfactin Production
2.5. Whole Genome Sequencing
2.6. Sequence Alignments
2.7. Quantitative PCR (qRT-PCR)
2.8. Macroscopic and Microscopic Analyses of Oats
2.9. Statistical Analysis
3. Results and Discussion
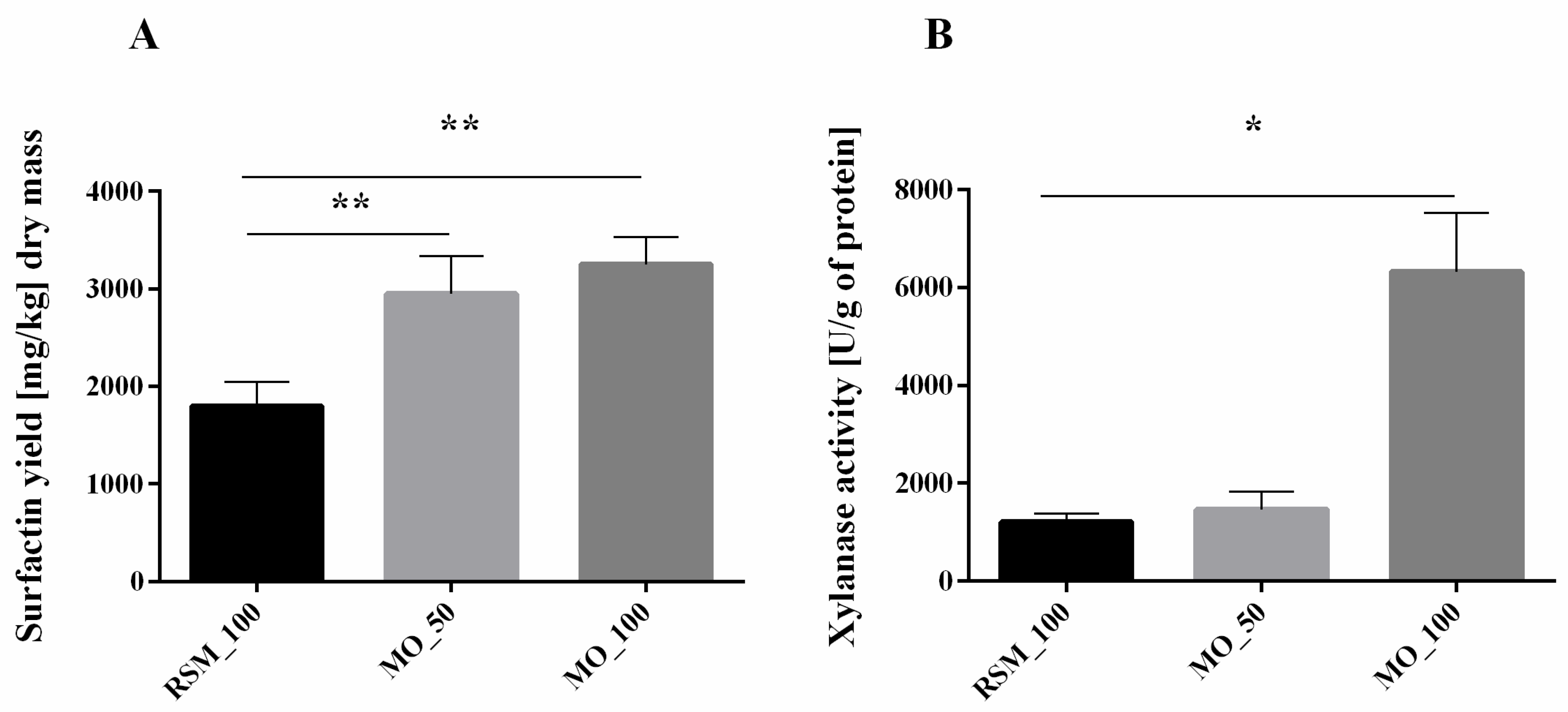
3.1. Production of Surfactin and Activity of Xylanase Depend on Hemicellulose Content in Cell Walls
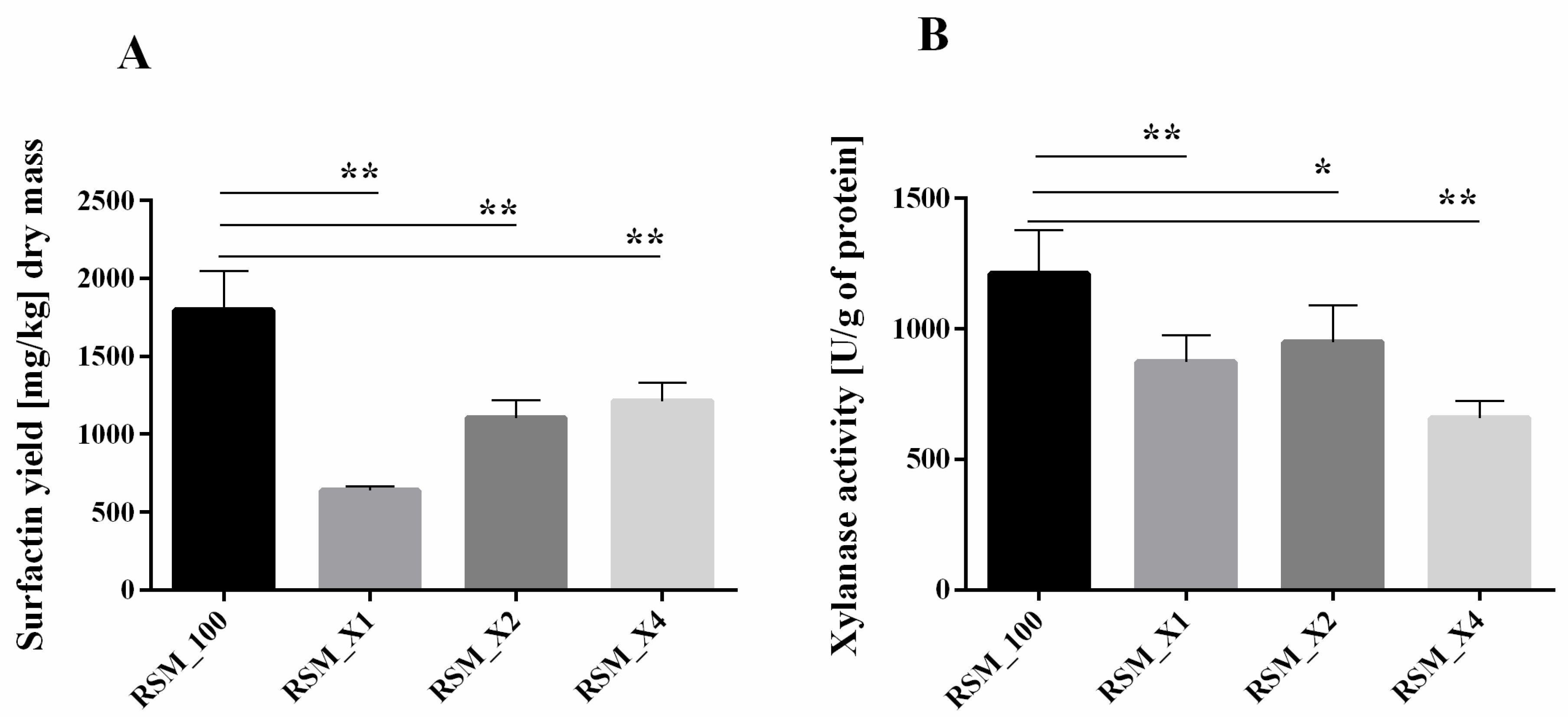
3.2. Xylan Addition Affects Surfactin Production or Xylanase Activity during SSF
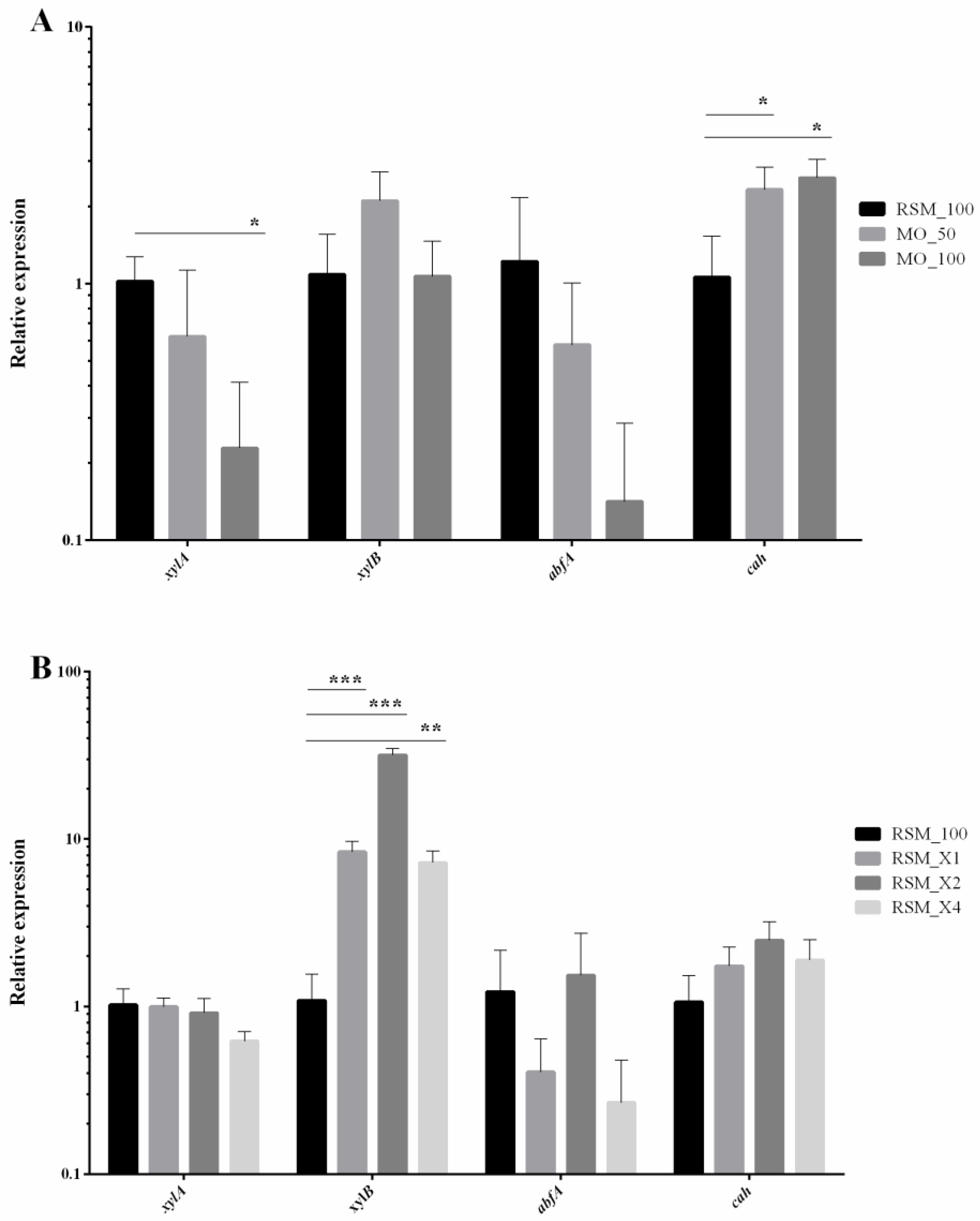
3.3. Milled Oats and Pure Xylan Increase Expression of Specific Xylanolytic Enzymes
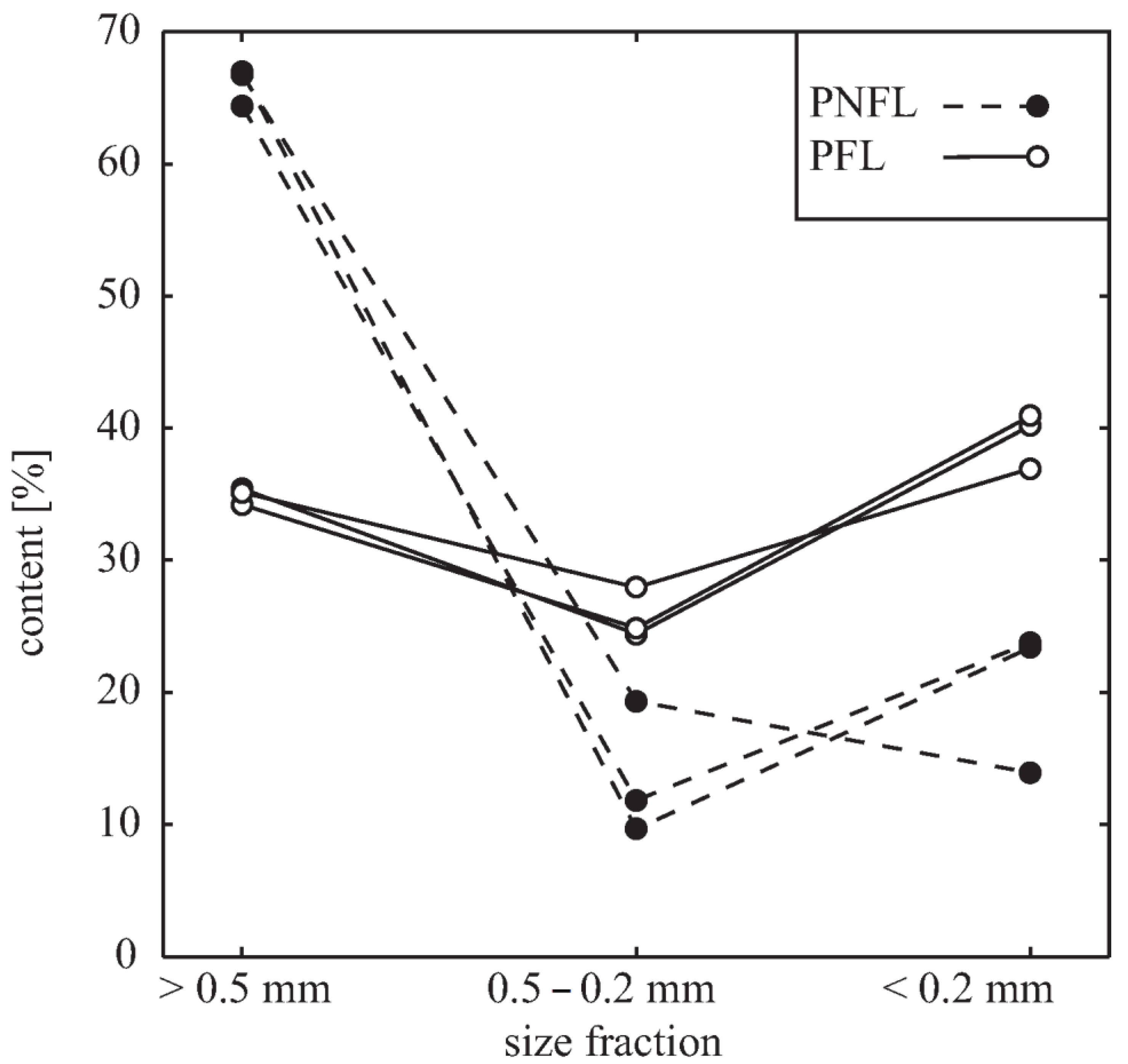
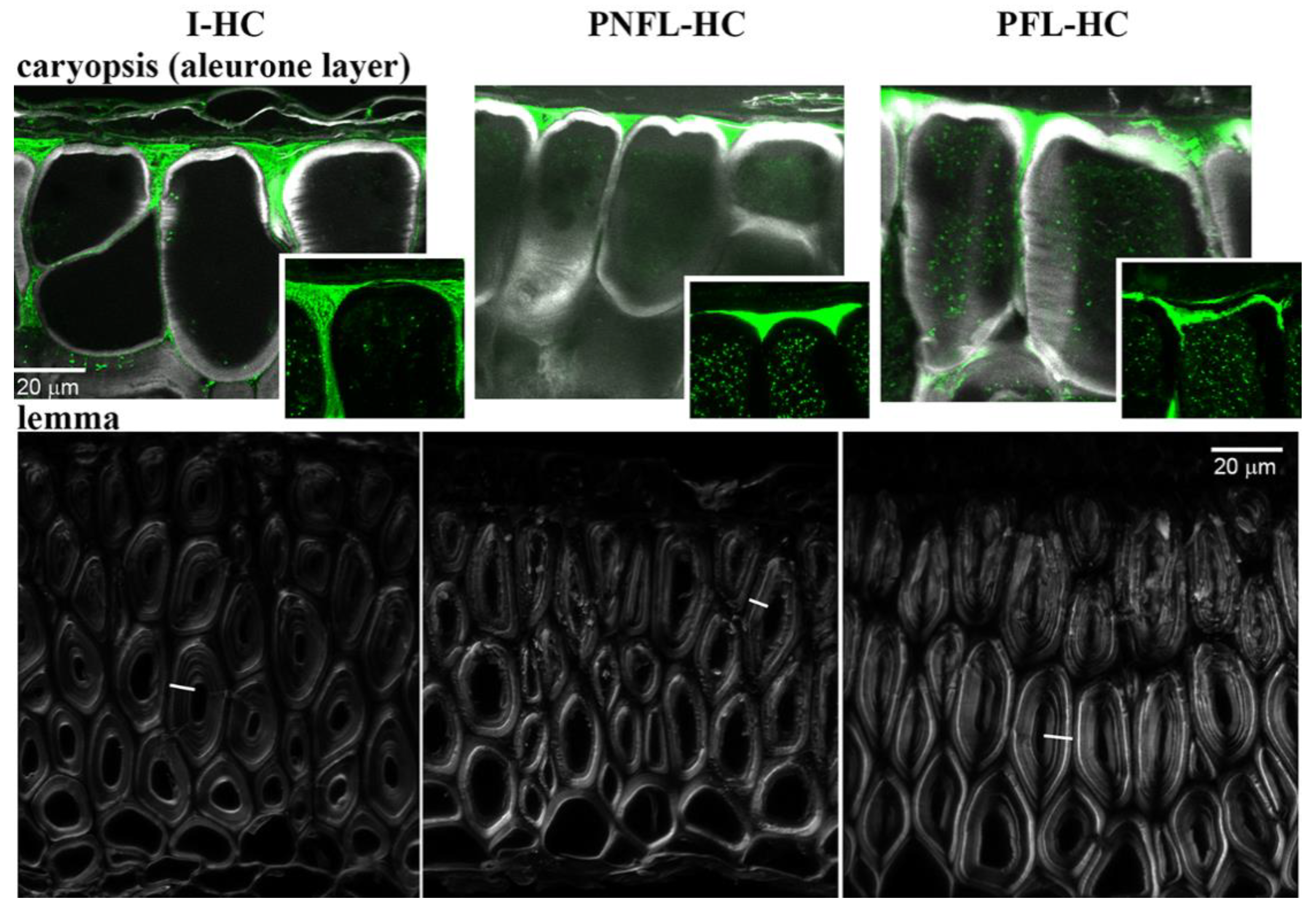
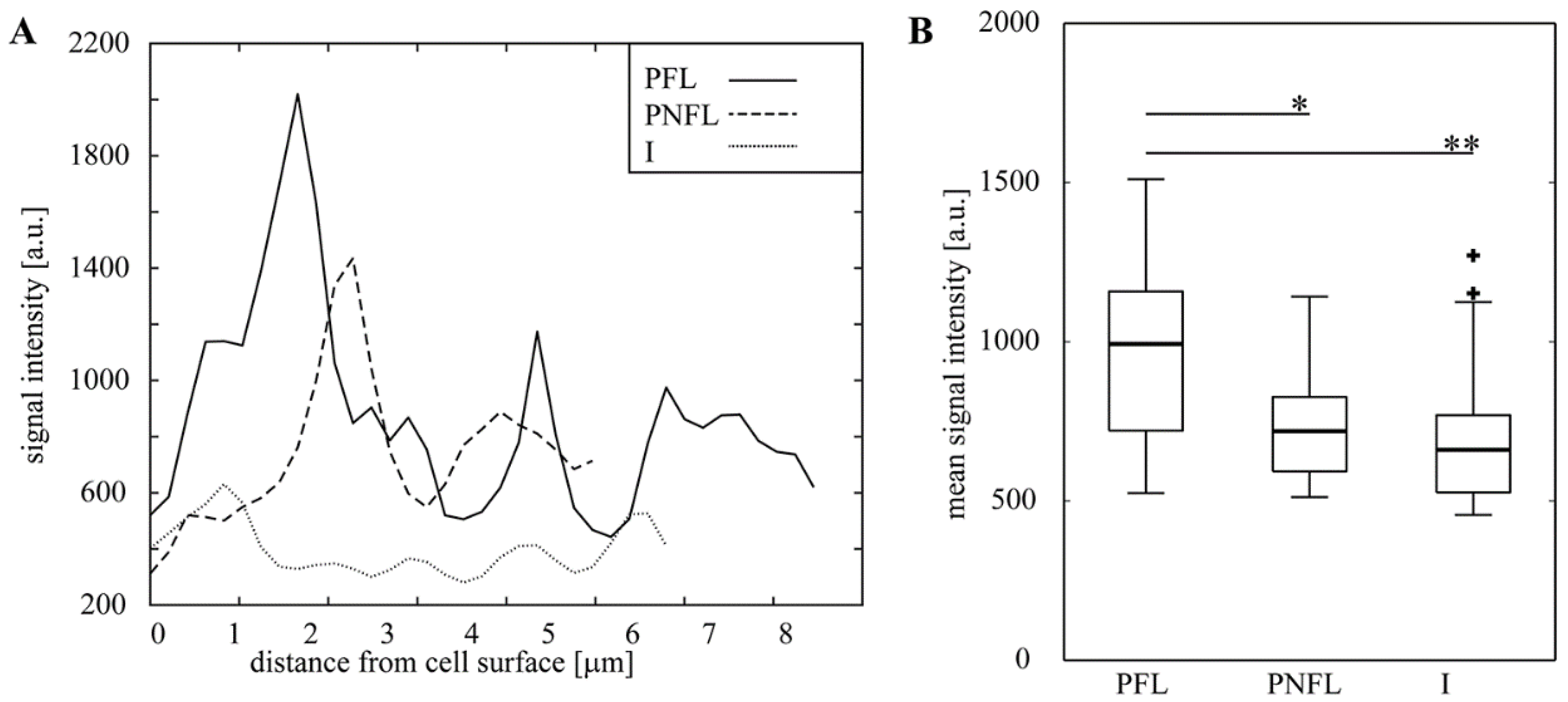
3.4. Fermentation of Oats Affects the Cell Wall Content and Integrity
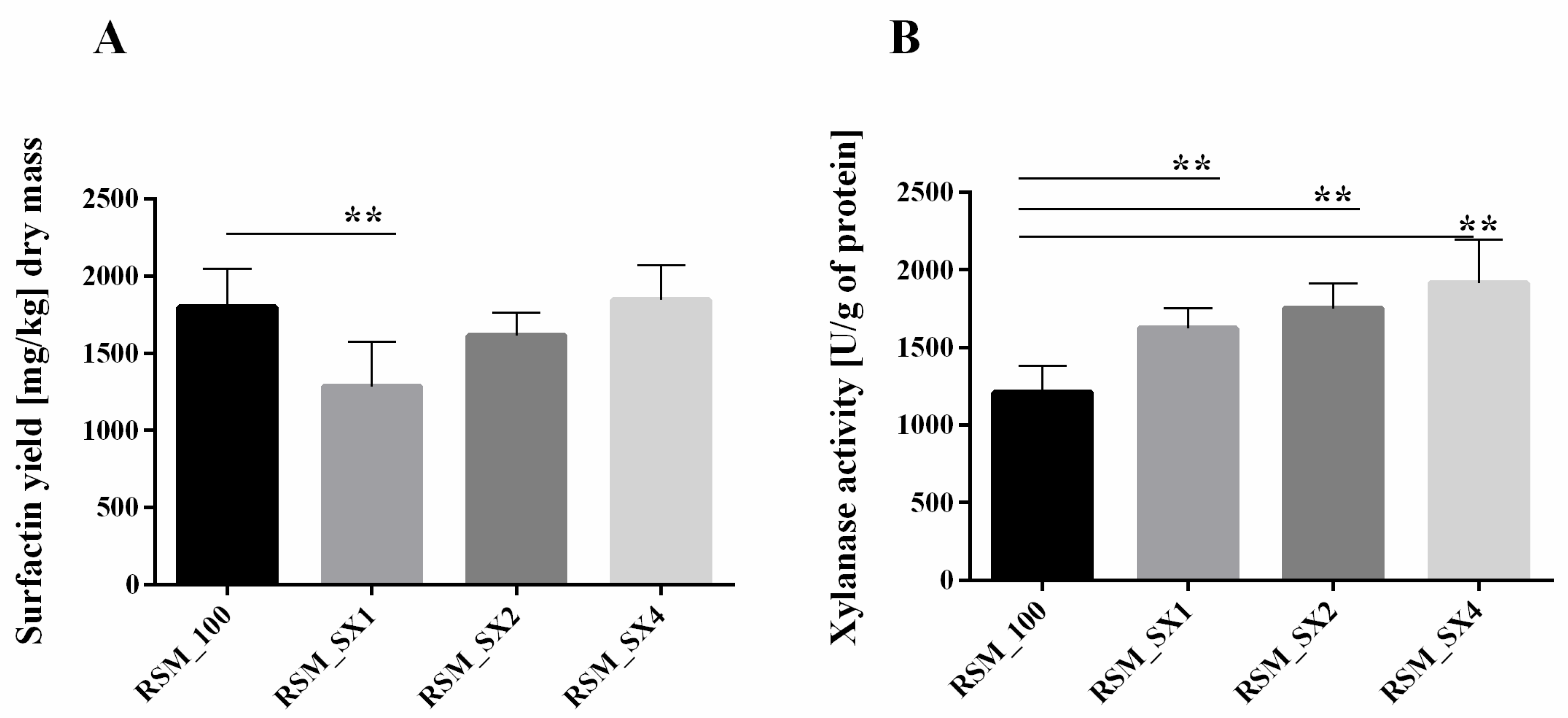
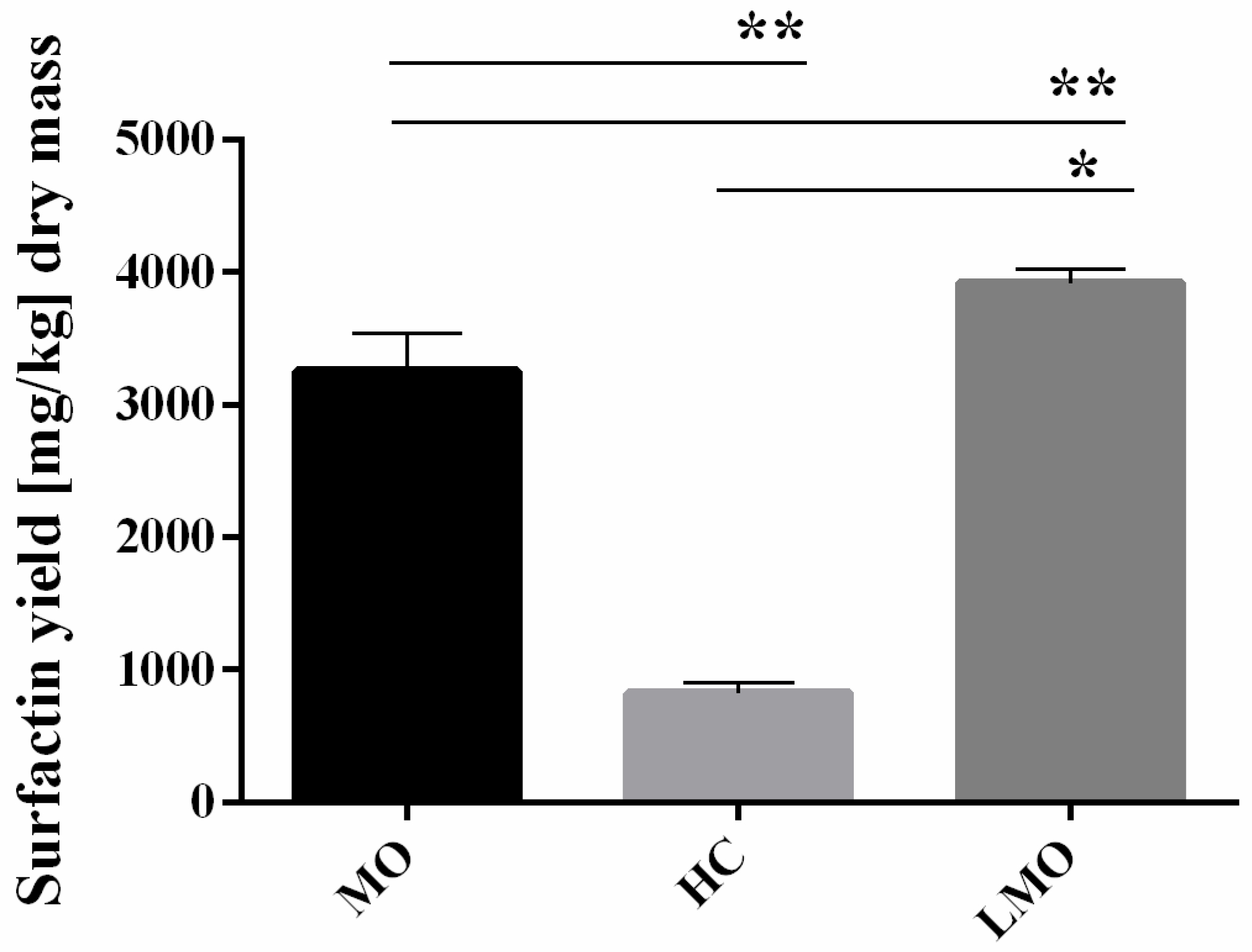
3.5. Components of Oat Hulls Induce Surfactin Production
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Han, X.; Guo, Y.; Liu, X.; Xia, Q.; Wang, Y. Catalytic conversion of lignocellulosic biomass into hydrocarbons: A mini review. Catal. Today 2019, 319, 2–13. [Google Scholar] [CrossRef]
- Lee, Y.; Kwon, E.E.; Lee, J. Polymers derived from hemicellulosic parts of lignocellulosic biomass. Rev. Environ. Sci. Biotechnol. 2019, 18, 317–334. [Google Scholar] [CrossRef]
- Matsagar, B.M.; Hossain, S.A.; Islam, T.; Alamri, H.R.; Alothman, Z.A.; Yamauchi, Y.; Dhepe, P.L.; Wu, K.C.W. Direct Production of Furfural in One-pot Fashion from Raw Biomass Using Brønsted Acidic Ionic Liquids. Sci. Rep. 2017, 7, 4–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Wu, Y.; Li, J.; He, T.; Xiao, Y.; Zhou, C.; Hu, C. Directing the Simultaneous Conversion of Hemicellulose and Cellulose in Raw Biomass to Lactic Acid. ACS Sustain. Chem. Eng. 2020, 8, 4244–4255. [Google Scholar] [CrossRef]
- Figueiredo, R.; Araújo, P.; Llerena, J.P.P.; Mazzafera, P. Suberin and hemicellulose in sugarcane cell wall architecture and crop digestibility: A biotechnological perspective. Food Energy Secur. 2019, 8, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Annu. Rev. Plant Biol. 2010, 61, 263–289. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.M.; Susan van Dyk, J.; Pletschke, B.I. Bacillus subtilis SJ01 produces hemicellulose degrading milti-enzyme complexes. BioResources 2012, 7, 1294–1309. [Google Scholar] [CrossRef]
- Naidu, D.S.; Hlangothi, S.P.; John, M.J. Bio-based products from xylan: A review. Carbohydr. Polym. 2018, 179, 28–41. [Google Scholar] [CrossRef] [PubMed]
- Biely, P.; Singh, S.; Puchart, V. Towards enzymatic breakdown of complex plant xylan structures: State of the art. Biotechnol. Adv. 2016, 34, 1260–1274. [Google Scholar] [CrossRef]
- Paraszkiewicz, K.; Bernat, P.; Kuśmierska, A.; Chojniak, J.; Płaza, G. Structural identification of lipopeptide biosurfactants produced by Bacillus subtilis strains grown on the media obtained from renewable natural resources. J. Environ. Manag. 2018, 209, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Liang, Y.; Wu, M.; Chen, Z.; Lin, J.; Yang, L. Natural products from Bacillus subtilis with antimicrobial properties. Chin. J. Chem. Eng. 2015, 23, 744–754. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; de Bruijn, I.; Nybroe, O.; Ongena, M. Natural functions of lipopeptides from Bacillus and Pseudomonas: More than surfactants and antibiotics. FEMS Microbiol. Rev. 2010, 34, 1037–1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jajor, P.; Piłakowska-Pietras, D.; Krasowska, A.; Łukaszewicz, M. Surfactin analogues produced by Bacillus subtilis strains grown on rapeseed cake. J. Mol. Struct. 2016, 1126, 141–146. [Google Scholar] [CrossRef]
- Debois, D.; Fernandez, O.; Franzil, L.; Jourdan, E.; de Brogniez, A.; Willems, L.; Clément, C.; Dorey, S.; De Pauw, E.; Ongena, M. Plant polysaccharides initiate underground crosstalk with bacilli by inducing synthesis of the immunogenic lipopeptide surfactin. Environ. Microbiol. Rep. 2015, 7, 570–582. [Google Scholar] [CrossRef] [PubMed]
- Szmigiel, I.; Suchodolski, J.; Łukaszewicz, M.; Krasowska, A. The influence of Bacillus subtilis 87Y isolated from Eisenia fetida on the growth of pathogenic and probiotic microorganisms. Biomass Convers. Biorefinery 2019. [Google Scholar] [CrossRef] [Green Version]
- Pandey, A. Solid-state fermentation. Biochem. Eng. J. 2003, 13, 81–84. [Google Scholar] [CrossRef]
- Cerda, A.; Artola, A.; Barrena, R.; Font, X.; Gea, T.; Sánchez, A. Innovative Production of Bioproducts From Organic Waste Through Solid-State Fermentation. Front. Sustain. Food Syst. 2019, 3, 1–6. [Google Scholar] [CrossRef] [Green Version]
- van Soest, P.J.; Wine, R.H. Use of Detergents in the Analysis of Fibrous Feeds. IV. Determination of Plant Cell-Wall Constituents. AOAC 1963, 13, 825–835. [Google Scholar]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Irfan, M.; Nadeem, M.; Syed, Q.; Baig, S. Effect of Medium Composition on Xylanase Production by Bacillus subtilis using Various Agricultural Wastes. J. Agric. Environ. Sci. 2012, 12, 561–565. [Google Scholar]
- McCartney, L.; Marcus, S.E.; Knox, J.P. Monoclonal antibodies to plant cell wall xylans and arabinoxylans. J. Histochem. Cytochem. 2005, 53, 543–546. [Google Scholar] [CrossRef] [PubMed]
- Voiniciuc, C.; Pauly, M.; Usadel, B. Monitoring polysaccharide dynamics in the plant cell wall. Plant Physiol. 2018, 176, 2590–2600. [Google Scholar] [CrossRef] [Green Version]
- Costa, J.A.V.; Treichel, H.; Santos, L.O.; Martins, V.G. Solid-State Fermentation for the Production of Biosurfactants and Their Applications. Curr. Dev. Biotechnol. Bioeng. 2018, 357–372. [Google Scholar] [CrossRef]
- Zhi, Y.; Wu, Q.; Xu, Y. Production of surfactin from waste distillers’ grains by co-culture fermentation of two Bacillus amyloliquefaciens strains. Bioresour. Technol. 2017, 235, 96–103. [Google Scholar] [CrossRef]
- Chen, C.; Lin, J.; Wang, W.; Huang, H.; Li, S. Cost-Effective Production of Surfactin from Xylose-Rich Corncob Hydrolysate Using Bacillus subtilis BS-37. Waste Biomass Valorization 2019, 10, 341–347. [Google Scholar] [CrossRef]
- Gudiña, E.J.; Fernandes, E.C.; Rodrigues, A.I.; Teixeira, J.A.; Rodrigues, L.R. Biosurfactant production by Bacillus subtilis using corn steep liquor as culture medium. Front. Microbiol. 2015, 6, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Cagri-Mehmetoglu, A.; Kusakli, S.; van de Venter, M. Production of polysaccharide and surfactin by Bacillus subtilis ATCC 6633 using rehydrated whey powder as the fermentation medium. J. Dairy Sci. 2012, 95, 3643–3649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurjar, J.; Sengupta, B. Production of surfactin from rice mill polishing residue by submerged fermentation using Bacillus subtilis MTCC 2423. Bioresour. Technol. 2015, 189, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Geissler, M.; Oellig, C.; Moss, K.; Schwack, W.; Henkel, M.; Hausmann, R. High-performance thin-layer chromatography (HPTLC) for the simultaneous quantification of the cyclic lipopeptides Surfactin, Iturin A and Fengycin in culture samples of Bacillus species. J. Chromatogr. B 2017, 1044–1045, 214–224. [Google Scholar] [CrossRef]
- Vogel, J. Unique aspects of the grass cell wall. Curr. Opin. Plant Biol. 2008, 11, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Balakrishnan, H.; Srinivasan, M.C.; Rele, M.V. Extracellular protease activities in relation to xylanase secretion in an alkalophilic Bacillus sp. Biotechnol. Lett. 1997, 19, 599–601. [Google Scholar] [CrossRef]
- Lindner, C.; Stulke, J.; Hecker, M. Regulation of xylanolytic enzymes in Bacillus subtilis. Microbiology 1994, 140, 753–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmiedel, D.; Hillen, W. A Bacillus subtilis 168 mutant with increased xylose uptake can utilize xylose as sole carbon source. FEMS Microbiol. Lett. 1996, 135, 175–178. [Google Scholar] [CrossRef]
- Walia, A.; Guleria, S.; Mehta, P.; Chauhan, A.; Parkash, J. Microbial xylanases and their industrial application in pulp and paper biobleaching: A review. 3 Biotech 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajpai, P. Microbial xylanolytic enzyme system: Properties and applications. Adv. Appl. Microbiol. 1997, 43, 141–194. [Google Scholar] [CrossRef]
- Knudsen, K.E.B. Carbohydrate and lignin contents of plant materials used in animal feeding. Anim. Feed Sci. Technol. 1997, 67, 319–338. [Google Scholar] [CrossRef]
- Burton, R.A.; Fincher, G.B. Evolution and development of cell walls in cereal grains. Front. Plant Sci. 2014, 5, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Van Craeyveld, V.; Dornez, E.; Holopainen, U.; Selinheimo, E.; Poutanen, K.; Delcour, J.A.; Courtin, C.M. Wheat Bran AX Properties and Choice of Xylanase Affect Enzymic Production of Wheat Bran-Derived Arabinoxylan-Oligosaccharides. Cereal Chem. 2010, 87, 283–291. [Google Scholar] [CrossRef]
- Dornez, E.; Holopainen, U.; Cuyvers, S.; Poutanen, K.; Delcour, J.A.; Courtin, C.M.; Nordlund, E. Study of grain cell wall structures by microscopic analysis with four different staining techniques. J. Cereal Sci. 2011. [Google Scholar] [CrossRef]
- Dornez, E.; Cuyvers, S.; Holopainen, U.; Nordlund, E.; Poutanen, K.; Delcour, J.A.; Courtin, C.M. Inactive Fluorescently Labeled Xylanase as a Novel Probe for Microscopic Analysis of Arabinoxylan Containing Cereal Cell Walls. J. Agric. Food Chem. 2011, 59, 6369–6375. [Google Scholar] [CrossRef] [PubMed]
- Jääskeläinen, A.S.; Holopainen-Mantila, U.; Tamminen, T.; Vuorinen, T. Endosperm and aleurone cell structure in barley and wheat as studied by optical and Raman microscopy. J. Cereal Sci. 2013, 57, 543–550. [Google Scholar] [CrossRef]
- Slivinski, C.T.; Mallmann, E.; de Araújo, J.M.; Mitchell, D.A.; Krieger, N. Production of surfactin by Bacillus pumilus UFPEDA 448 in solid-state fermentation using a medium based on okara with sugarcane bagasse as a bulking agent. Process Biochem. 2012, 47, 1848–1855. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Zhang, F.; Wei, Z.; Ran, W.; Shen, Q. The usage of rice straw as a major substrate for the production of surfactin by Bacillus amyloliquefaciens XZ-173 in solid-state fermentation. J. Environ. Manage. 2013, 127, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Redaelli, R.; Berardo, N. Prediction of fibre components in oat hulls by near infrared reflectance spectroscopy. J. Sci. Food Agric. 2007, 1243, 1237–1243. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence [5’–3’] | Amplification Efficiency [%] |
|---|---|---|
| gyrB_F | TAATGGCGGCAAGAGCAAGA | 102.08 |
| gyrB_R | ATGTCTGTCGCGTCCTTGTT | |
| xynA_F | GCGAACCTGTAGTCCAACCTT | 102.6 |
| xynA_R | TTTTCGGCAACCGCCTCT | |
| xynB_F | AAACTGACAGAAGCTCCGCA | 89.3 |
| xynB_R | GGATTTCCTGGGTCATGCCA | |
| abfA_F | AGAGCCTTTCGGATGGTTGC | 100.2 |
| abfA_R | GACGGCTTGACTTGGCATG | |
| cah_F | TGCAGGCGATGAAGACACTT | 86.16 |
| cah_R | GCGGTACACCTTCAGCTCTT |
| Solid Base | Hemicellulose [%] | Hemicellulose Loss [%] | |
|---|---|---|---|
| NF | F | ||
| RSM_100 | 12.17 | 12.11 | −0.49 |
| MO_50 | 19.58 | 15.43 | −21.19 |
| MO_100 | 39.86 | 26.83 | −32.68 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szmigiel, I.; Kwiatkowska, D.; Łukaszewicz, M.; Krasowska, A. Xylan Decomposition in Plant Cell Walls as an Inducer of Surfactin Synthesis by Bacillus subtilis. Biomolecules 2021, 11, 239. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11020239
Szmigiel I, Kwiatkowska D, Łukaszewicz M, Krasowska A. Xylan Decomposition in Plant Cell Walls as an Inducer of Surfactin Synthesis by Bacillus subtilis. Biomolecules. 2021; 11(2):239. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11020239
Chicago/Turabian StyleSzmigiel, Ida, Dorota Kwiatkowska, Marcin Łukaszewicz, and Anna Krasowska. 2021. "Xylan Decomposition in Plant Cell Walls as an Inducer of Surfactin Synthesis by Bacillus subtilis" Biomolecules 11, no. 2: 239. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11020239