
In Silico Analysis of the Enzymes Involved in Haloarchaeal Denitrification

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Selection of Species for the Analysis of Sequences Alignments
2.2. Sequence Analysis and Alignments
2.3. Model Building
2.4. Analysis of Electrostatic Surface Potential and Illustration
3. Results and Discussion
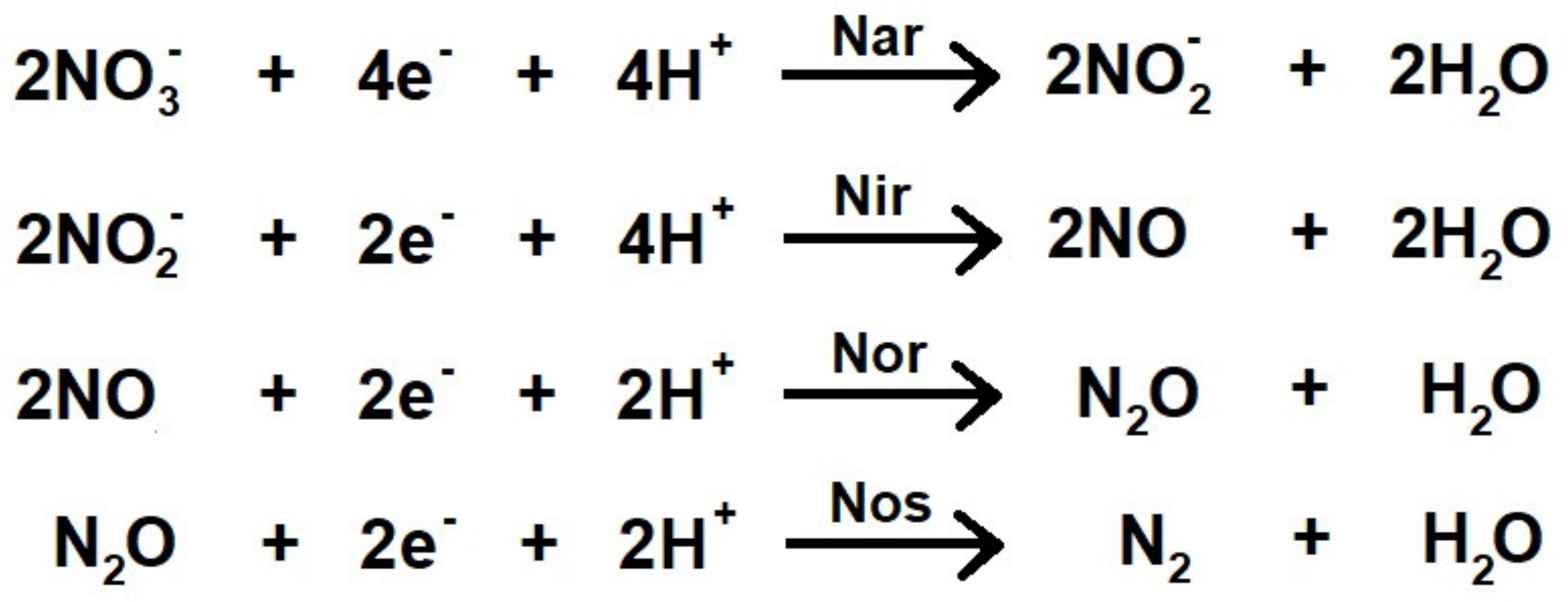
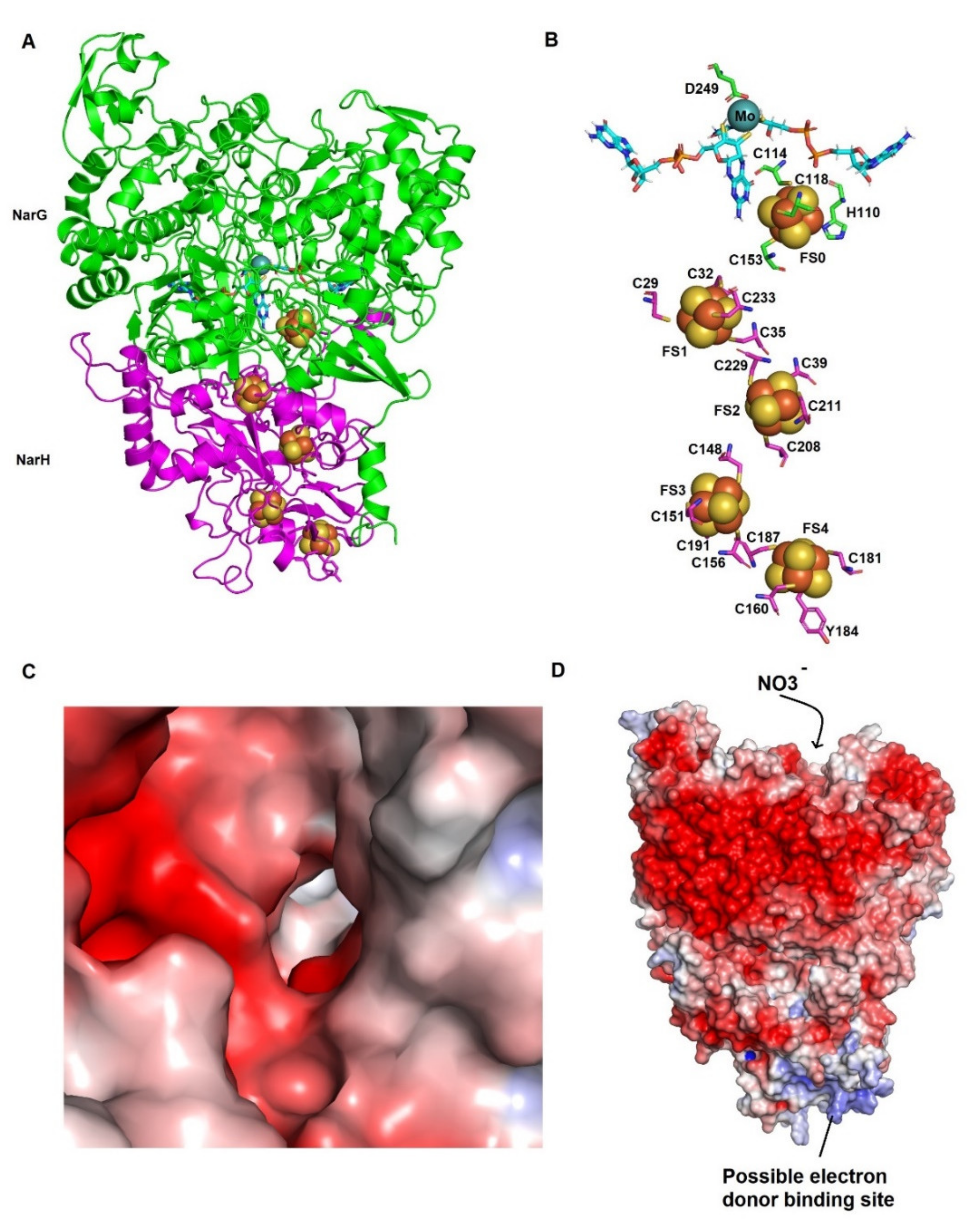
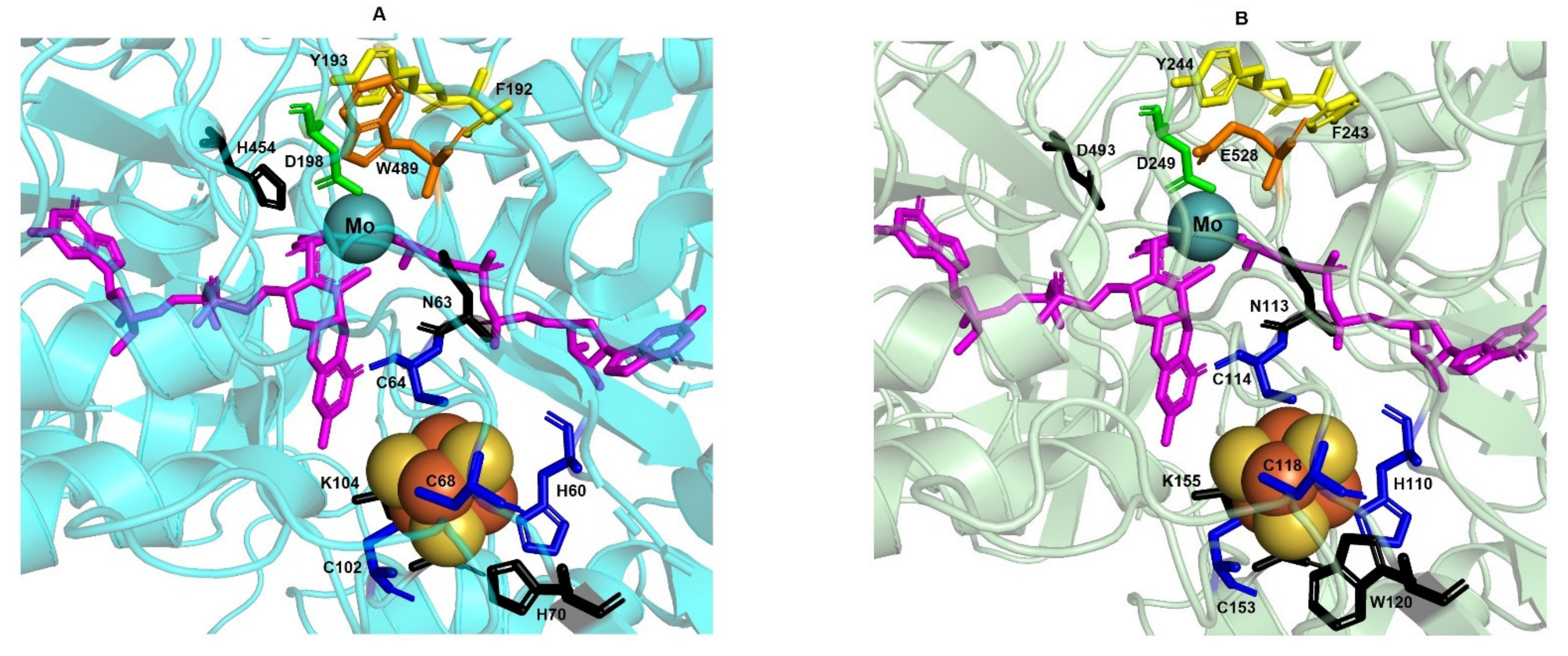
3.1. Respiratory Nitrate Reductase
3.1.1. N-terminal Analysis Exposes Conflict of Annotation in Several Haloarchaeal Species
3.1.2. The Structural Model Shows Spatial Conservation of All Residues Involved in Cofactors Binding
3.2. Respiratory Nitrite Reductase
3.2.1. N-Terminal Inspection Reveals a TAT Signal Peptide
3.2.2. The Overall Structure Is Consistent with an Homotrimer with Conserved Residues Involved in the Catalytic Mechanism
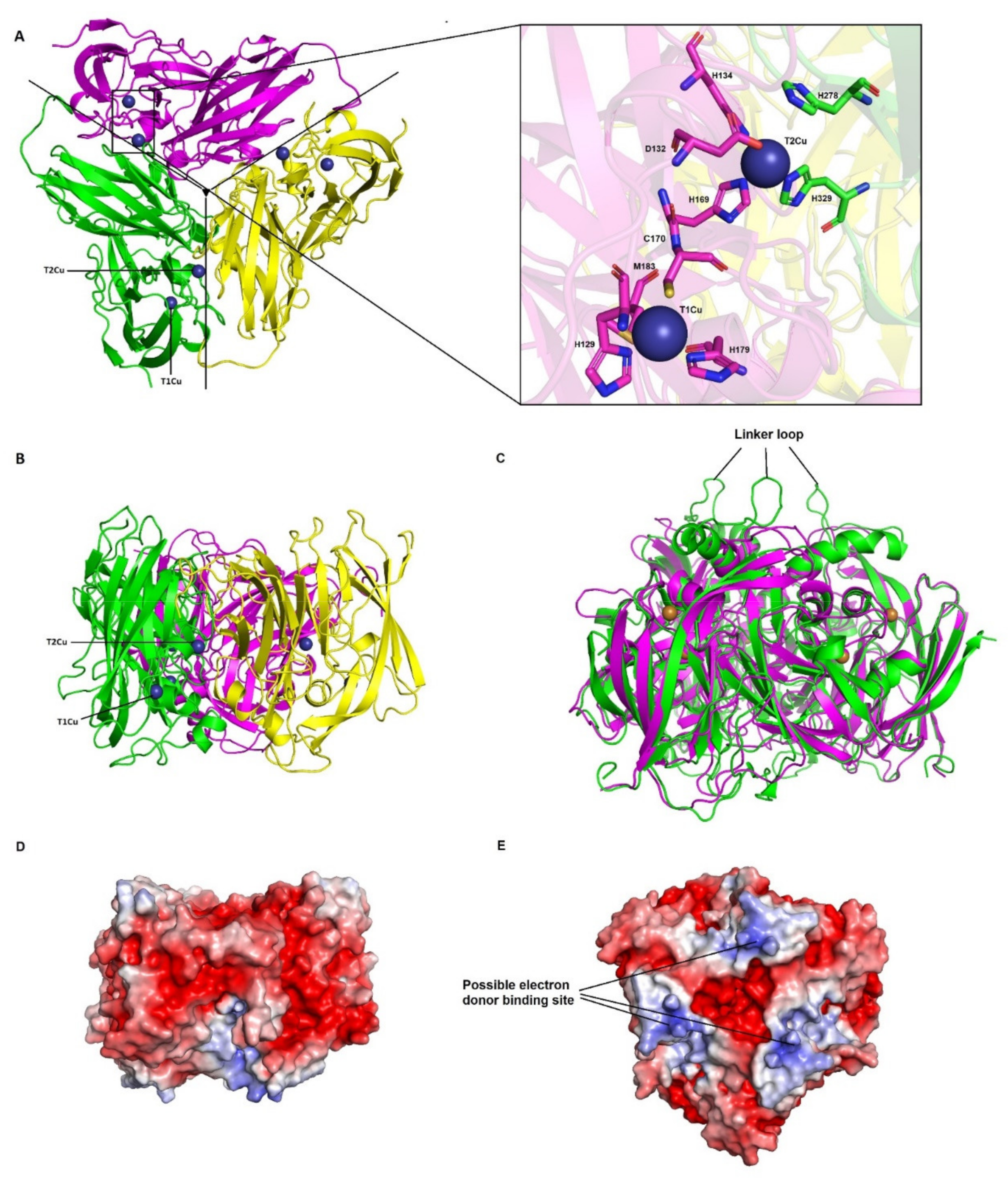
3.3. Nitrous Oxide Reductase
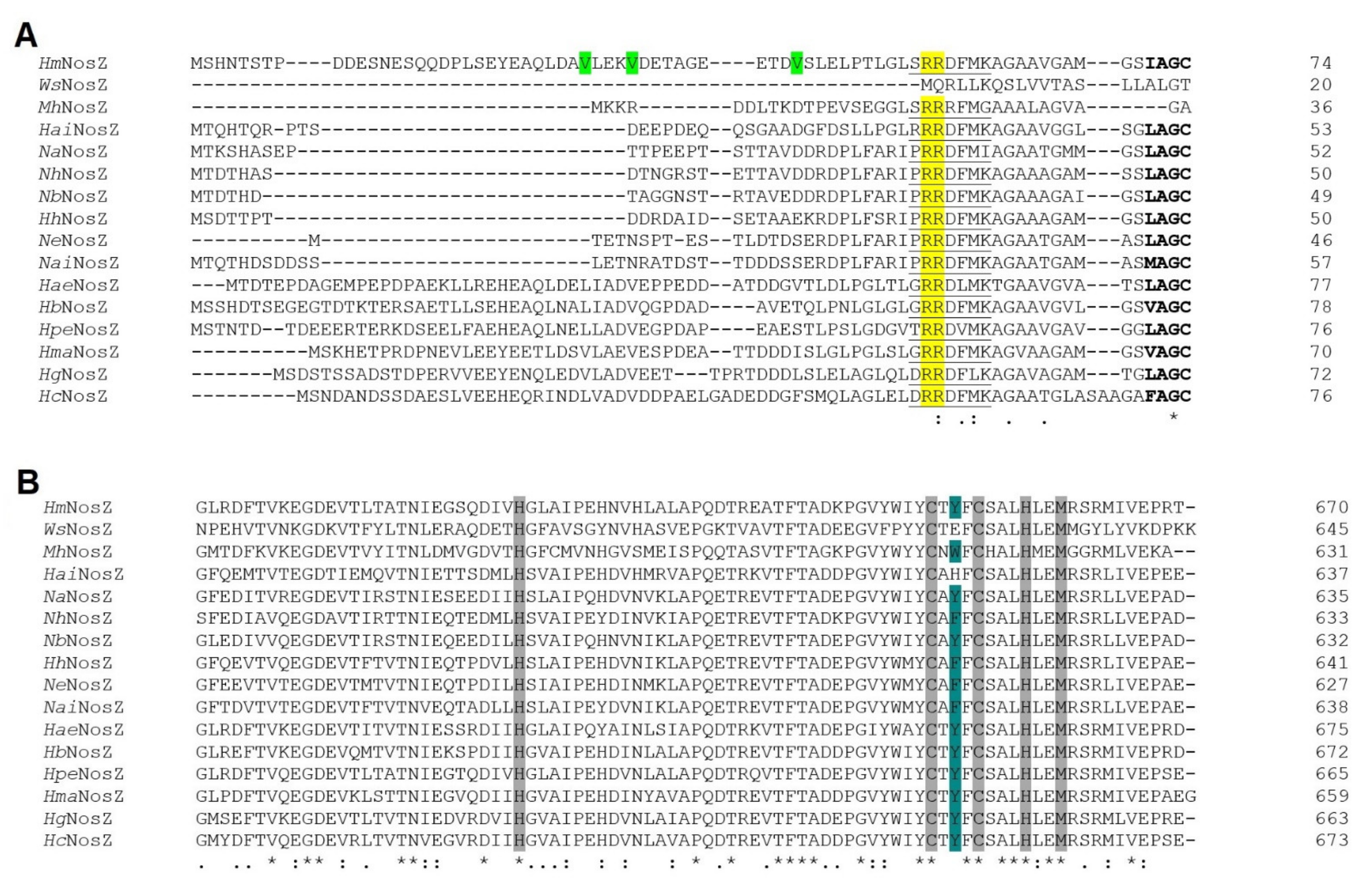
3.3.1. N-Terminal Analysis Shows Same Annotation Conflict as in Respiratory Nitrate Reductase
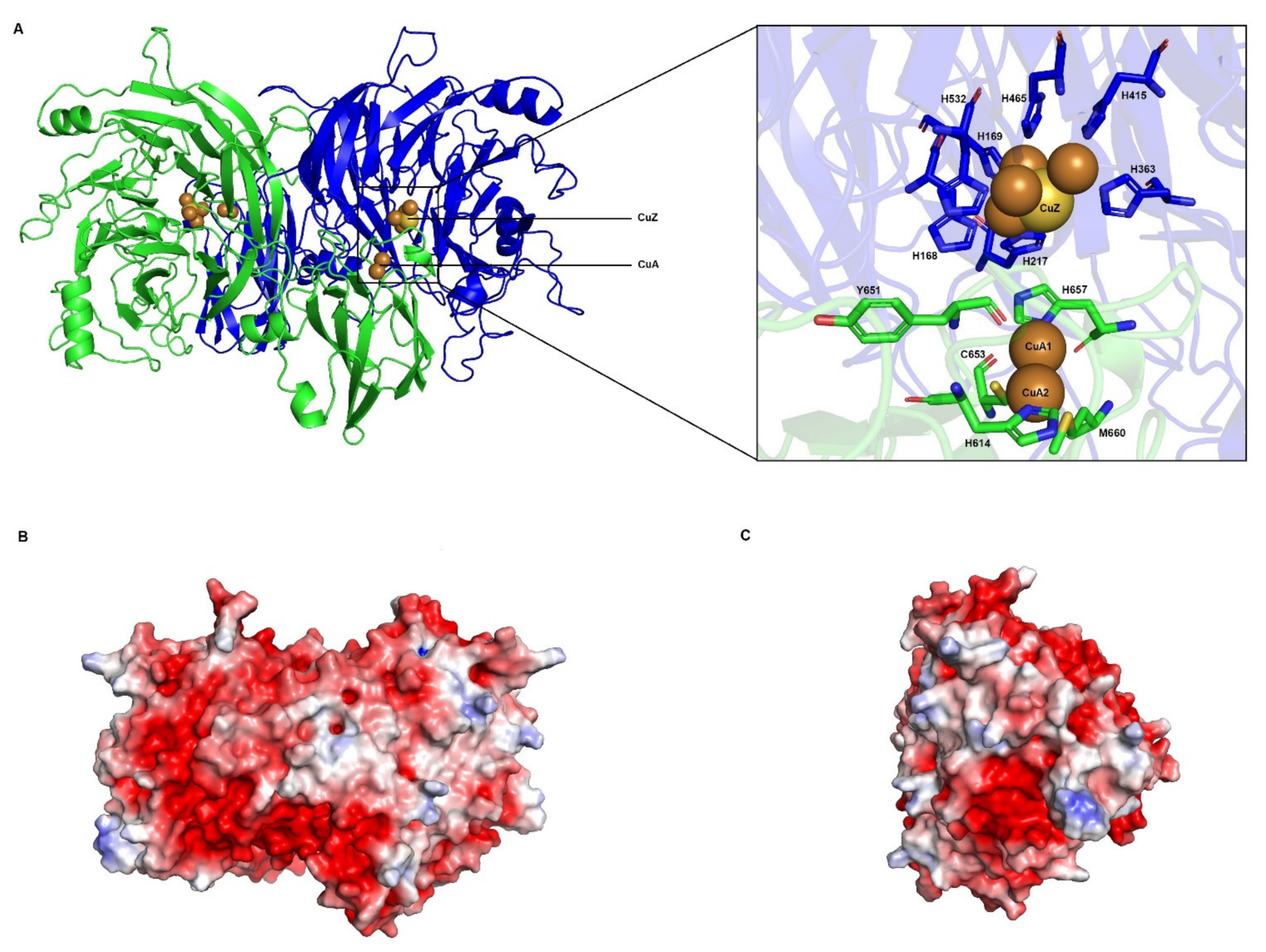
3.3.2. CuA and CuZ Centres Are Conserved in Haloarchaea
3.3.3. Electron Transfer Pathways Show Partial Conservation and Similarity to Clade II NosZ
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Thomson, A.J.; Giannopoulos, G.; Pretty, J.; Baggs, E.M.; Richardson, D.J. Biological Sources and Sinks of Nitrous Oxide and Strategies to Mitigate Emissions. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 1157–1168. [Google Scholar] [CrossRef] [Green Version]
- Philippot, L. Denitrifying Genes in Bacterial and Archaeal Genomes. Biochim. Biophys. Acta 2002, 1577, 355–376. [Google Scholar] [CrossRef]
- Zumft, W.G.; Kroneck, P.M.H. Respiratory Transformation of Nitrous Oxide (N2O) to Dinitrogen by Bacteria and Archaea. Adv. Microb. Physiol. 2007, 52, 107–227. [Google Scholar] [CrossRef] [PubMed]
- Richardson, D.J.; Watmough, N.J. Inorganic Nitrogen Metabolism in Bacteria. Curr. Opin. Chem. Biol. 1999, 3, 207–219. [Google Scholar] [CrossRef]
- Giannopoulos, G.; Sullivan, M.J.; Hartop, K.R.; Rowley, G.; Gates, A.J.; Watmough, N.J.; Richardson, D.J. Tuning the Modular Paracoccus Denitrificans Respirome to Adapt from Aerobic Respiration to Anaerobic Denitrification. Environ. Microbiol. 2017, 19, 4953–4964. [Google Scholar] [CrossRef] [Green Version]
- Lycus, P.; Lovise Bøthun, K.; Bergaust, L.; Peele Shapleigh, J.; Reier Bakken, L.; Frostegård, Å. Phenotypic and Genotypic Richness of Denitrifiers Revealed by a Novel Isolation Strategy. ISME J. 2017, 11, 2219–2232. [Google Scholar] [CrossRef] [Green Version]
- Torregrosa-Crespo, J.; Pire, C.; Martínez-Espinosa, R.M.; Bergaust, L. Denitrifying Haloarchaea within the Genus Haloferax Display Divergent Respiratory Phenotypes, with Implications for Their Release of Nitrogenous Gases. Environ. Microbiol. 2019, 21, 427–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torregrosa-Crespo, J.; Martínez-Espinosa, R.M.; Esclapez, J.; Bautista, V.; Pire, C.; Camacho, M.; Richardson, D.J.; Bonete, M.J. Anaerobic Metabolism in Haloferax Genus: Denitrification as Case of Study. Adv. Microb. Physiol. 2016, 68, 41–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, Z.; Bakken, L.R.; Molstad, L.; Frostegård, Å.; Bergaust, L.L. Transcriptional and Metabolic Regulation of Denitrification in Paracoccus denitrificans Allows Low but Significant Activity of Nitrous Oxide Reductase under Oxic Conditions. Environ. Microbiol. 2016, 18, 2951–2963. [Google Scholar] [CrossRef] [PubMed]
- Jormakka, M.; Richardson, D.; Byrne, B.; Iwata, S. Architecture of NarGH Reveals a Structural Classification of Mo-BisMGD Enzymes. Structure 2004, 12, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Bertero, M.G.; Rothery, R.A.; Palak, M.; Hou, C.; Lim, D.; Blasco, F.; Weiner, J.H.; Strynadka, N.C.J. Insights into the Respiratory Electron Transfer Pathway from the Structure of Nitrate Reductase A. Nat. Struct. Biol. 2003, 10, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Bergaust, L.; Shapleigh, J.; Frostegård, A.; Bakken, L. Transcription and Activities of NOx Reductases in Agrobacterium tumefaciens: The Influence of Nitrate, Nitrite and Oxygen Availability. Environ. Microbiol. 2008, 10, 3070–3081. [Google Scholar] [CrossRef] [PubMed]
- Oren, A. Halophilic Microbial Communities and Their Environments. Curr. Opin. Biotechnol. 2015, 33, 119–124. [Google Scholar] [CrossRef]
- Oren, A. Probing Saltern Brines with an Oxygen Electrode: What Can We Learn about the Community Metabolism in Hypersaline Systems? Life 2016, 6, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oren, A. Life at High Salt Concentrations, Intracellular KCl Concentrations, and Acidic Proteomes. Front. Microbiol. 2013, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeifer, K.; Ergal, İ.; Koller, M.; Basen, M.; Schuster, B.; Rittmann, S.K.-M.R. Archaea Biotechnology. Biotechnol. Adv. 2021, 47, 107668. [Google Scholar] [CrossRef]
- Feng, J.; Liu, B.; Zhang, Z.; Ren, Y.; Li, Y.; Gan, F.; Huang, Y.; Chen, X.; Shen, P.; Wang, L.; et al. The Complete Genome Sequence of Natrinema Sp. J7-2, a Haloarchaeon Capable of Growth on Synthetic Media without Amino Acid Supplements. PLoS ONE 2012, 7, e41621. [Google Scholar] [CrossRef] [Green Version]
- Yoshimatsu, K.; Sakurai, T.; Fujiwara, T. Purification and Characterization of Dissimilatory Nitrate Reductase from a Denitrifying Halophilic Archaeon, Haloarcula marismortui. FEBS Lett. 2000, 470, 216–220. [Google Scholar] [CrossRef] [Green Version]
- Ichiki, H.; Tanaka, Y.; Mochizuki, K.; Yoshimatsu, K.; Sakurai, T.; Fujiwara, T. Purification, Characterization, and Genetic Analysis of Cu-Containing Dissimilatory Nitrite Reductase from a Denitrifying Halophilic Archaeon, Haloarcula marismortui. J. Bacteriol. 2001, 183, 4149–4156. [Google Scholar] [CrossRef] [Green Version]
- Lledó, B.; Martínez-Espinosa, R.M.; Marhuenda-Egea, F.C.; Bonete, M.J. Respiratory Nitrate Reductase from Haloarchaeon Haloferax mediterranei: Biochemical and Genetic Analysis. Biochim. Biophys. Acta BBA Gen. Subj. 2004, 1674, 50–59. [Google Scholar] [CrossRef]
- Martinez-Espinosa, R.M.; Dridge, E.J.; Bonete, M.J.; Butt, J.N.; Butler, C.S.; Sargent, F.; Richardson, D.J. Look on the Positive Side! The Orientation, Identification and Bioenergetics of Archaeal Membrane-Bound Nitrate Reductases. FEMS Microbiol. Lett. 2007, 276, 129–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torregrosa-Crespo, J.; Pire, C.; Richardson, D.J.; Martínez-Espinosa, R.M. Exploring the Molecular Machinery of Denitrification in Haloferax mediterranei Through Proteomics. Front. Microbiol. 2020, 11, 3136. [Google Scholar] [CrossRef] [PubMed]
- Bonete, M.J.; Martínez-Espinosa, R.M.; Pire, C.; Zafrilla, B.; Richardson, D.J. Nitrogen Metabolism in Haloarchaea. Saline Syst. 2008, 4, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Espinosa, R.M. Heterologous and Homologous Expression of Proteins from Haloarchaea: Denitrification as Case of Study. Int. J. Mol. Sci. 2019, 21, 82. [Google Scholar] [CrossRef] [Green Version]
- Torregrosa-Crespo, J.; González-Torres, P.; Bautista, V.; Esclapez, J.M.; Pire, C.; Camacho, M.; Bonete, M.J.; Richardson, D.J.; Watmough, N.J.; Martínez-Espinosa, R.M. Analysis of Multiple Haloarchaeal Genomes Suggests That the Quinone-Dependent Respiratory Nitric Oxide Reductase Is an Important Source of Nitrous Oxide in Hypersaline Environments. Environ. Microbiol. Rep. 2017, 9, 788–796. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, Scalable Generation of High-Quality Protein Multiple Sequence Alignments Using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Söding, J. Protein Homology Detection by HMM–HMM Comparison. Bioinformatics 2005, 21, 951–960. [Google Scholar] [CrossRef] [Green Version]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 Improves Signal Peptide Predictions Using Deep Neural Networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- DE Castro, E.; Sigrist, C.J.A.; Gattiker, A.; Bulliard, V.; Langendijk-Genevaux, P.S.; Gasteiger, E.; Bairoch, A.; Hulo, N. ScanProsite: Detection of PROSITE Signature Matches and ProRule-Associated Functional and Structural Residues in Proteins. Nucleic Acids Res. 2006, 34, W362–W365. [Google Scholar] [CrossRef]
- Arnold, K.; Bordoli, L.; Kopp, J.; Schwede, T. The SWISS-MODEL Workspace: A Web-Based Environment for Protein Structure Homology Modelling. Bioinformatics 2006, 22, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Bordoli, L.; Kiefer, F.; Arnold, K.; Benkert, P.; Battey, J.; Schwede, T. Protein Structure Homology Modeling Using SWISS-MODEL Workspace. Nat. Protoc. 2009, 4, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Benkert, P.; Biasini, M.; Schwede, T. Toward the Estimation of the Absolute Quality of Individual Protein Structure Models. Bioinforma. Oxf. Engl. 2011, 27, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Studer, G.; Rempfer, C.; Waterhouse, A.M.; Gumienny, R.; Haas, J.; Schwede, T. QMEANDisCo-Distance Constraints Applied on Model Quality Estimation. Bioinforma. Oxf. Engl. 2020, 36, 1765–1771. [Google Scholar] [CrossRef] [PubMed]
- González, P.J.; Correia, C.; Moura, I.; Brondino, C.D.; Moura, J.J.G. Bacterial Nitrate Reductases: Molecular and Biological Aspects of Nitrate Reduction. J. Inorg. Biochem. 2006, 100, 1015–1023. [Google Scholar] [CrossRef] [PubMed]
- Youngblut, M.D.; Tsai, C.-L.; Clark, I.C.; Carlson, H.K.; Maglaqui, A.P.; Gau-Pan, P.S.; Redford, S.A.; Wong, A.; Tainer, J.A.; Coates, J.D. Perchlorate Reductase Is Distinguished by Active Site Aromatic Gate Residues. J. Biol.Chem. 2016, 291, 9190–9202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickner, W.; Schekman, R. Protein Translocation Across Biological Membranes. Science 2005, 310, 1452–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, P.A.; Tullman-Ercek, D.; Georgiou, G. The Bacterial Twin-Arginine Translocation Pathway. Annu. Rev. Microbiol. 2006, 60, 373–395. [Google Scholar] [CrossRef] [Green Version]
- Hutcheon, G.W.; Bolhuis, A. The Archaeal Twin-Arginine Translocation Pathway. Biochem. Soc. Trans. 2003, 31, 686–689. [Google Scholar] [CrossRef] [Green Version]
- Dilks, K.; Giménez, M.I.; Pohlschröder, M. Genetic and Biochemical Analysis of the Twin-Arginine Translocation Pathway in Halophilic Archaea. J. Bacteriol. 2005, 187, 8104–8113. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, D.; Boral, D.; Vankudoth, K.R.; Ramasamy, S. Analysis of Haloarchaeal Twin-Arginine Translocase Pathway Reveals the Diversity of the Machineries. Heliyon 2019, 5, e01587. [Google Scholar] [CrossRef] [Green Version]
- Giani, M.; Miralles-Robledillo, J.M.; Peiró, G.; Pire, C.; Martínez-Espinosa, R.M. Deciphering Pathways for Carotenogenesis in Haloarchaea. Molecules 2020, 25, 1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villegas, A.; Kropinski, A.M. An Analysis of Initiation Codon Utilization in the Domain Bacteria—Concerns about the Quality of Bacterial Genome Annotation. Microbiology 2008, 154, 2559–2661. [Google Scholar] [CrossRef] [Green Version]
- Miralles-Robledillo, J.M.; Torregrosa-Crespo, J.; Martínez-Espinosa, R.M.; Pire, C. DMSO Reductase Family: Phylogenetics and Applications of Extremophiles. Int. J. Mol. Sci. 2019, 20, 3349. [Google Scholar] [CrossRef] [Green Version]
- Iwata, S.; Saynovits, M.; Link, T.A.; Michel, H. Structure of a Water Soluble Fragment of the Rieske Iron-Sulfur Protein of the Bovine Heart Mitochondrial Cytochrome Bc1 Complex Determined by MAD Phasing at 1.5 å Resolution. Structure 1996, 4, 567–579. [Google Scholar] [CrossRef] [Green Version]
- Magalon, A.; Asso, M.; Guigliarelli, B.; Rothery, R.A.; Bertrand, P.; Giordano, G.; Blasco, F. Molybdenum Cofactor Properties and [Fe-S] Cluster Coordination in Escherichia coli Nitrate Reductase A: Investigation by Site-Directed Mutagenesis of the Conserved His-50 Residue in the NarG Subunit. Biochemistry 1998, 37, 7363–7370. [Google Scholar] [CrossRef]
- Jepson, B.J.N.; Mohan, S.; Clarke, T.A.; Gates, A.J.; Cole, J.A.; Butler, C.S.; Butt, J.N.; Hemmings, A.M.; Richardson, D.J. Spectropotentiometric and Structural Analysis of the Periplasmic Nitrate Reductase from Escherichia coli. J. Biol. Chem. 2007, 282, 6425–6437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marangon, J.; de Sousa, P.M.P.; Moura, I.; Brondino, C.D.; Moura, J.J.G.; González, P.J. Substrate-Dependent Modulation of the Enzymatic Catalytic Activity: Reduction of Nitrate, Chlorate and Perchlorate by Respiratory Nitrate Reductase from Marinobacter hydrocarbonoclasticus 617. Biochim. Biophys. Acta BBA Bioenerg. 2012, 1817, 1072–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sparacino-Watkins, C.; Stolz, J.F.; Basu, P. Nitrate and Periplasmic Nitrate Reductases. Chem. Soc. Rev. 2013, 43, 676–706. [Google Scholar] [CrossRef]
- Anderson, L.J.; Richardson, D.J.; Butt, J.N. Catalytic Protein Film Voltammetry from a Respiratory Nitrate Reductase Provides Evidence for Complex Electrochemical Modulation of Enzyme Activity. Biochemistry 2001, 40, 11294–11307. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, T.; Rova, M.; Smedja Bäcklund, A. Microbial Metabolism of Oxochlorates: A Bioenergetic Perspective. Biochim. Biophys. Acta 2013, 1827, 189–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Espinosa, R.M.; Richardson, D.J.; Bonete, M.J. Characterisation of Chlorate Reduction in the Haloarchaeon Haloferax mediterranei. Biochim. Biophys. Acta 2015, 1850, 587–594. [Google Scholar] [CrossRef] [Green Version]
- Morpeth, F.F.; Boxer, D.H. Kinetic Analysis of Respiratory Nitrate Reductase from Escherichia coli K12. Biochemistry 1985, 24, 40–46. [Google Scholar] [CrossRef]
- Heylen, K.; Gevers, D.; Vanparys, B.; Wittebolle, L.; Geets, J.; Boon, N.; De Vos, P. The Incidence of NirS and NirK and Their Genetic Heterogeneity in Cultivated Denitrifiers. Environ. Microbiol. 2006, 8, 2012–2021. [Google Scholar] [CrossRef]
- Shapleigh, J.P.; Payne, W.J. Differentiation of c, d1 Cytochrome and Copper Nitrite Reductase Production in Denitrifiers. FEMS Microbiol. Lett. 1985, 26, 275–279. [Google Scholar] [CrossRef]
- Xie, Y.; Inoue, T.; Seike, N.; Matsumura, H.; Kanbayashi, K.; Itoh, K.; Kataoka, K.; Yamaguchi, K.; Suzuki, S.; Kai, Y.; et al. Crystallization and Preliminary X-Ray Crystallographic Studies of Dissimilatory Nitrite Reductase Isolated from Hyphomicrobium denitrificans A3151. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 2383–2386. [Google Scholar] [CrossRef] [PubMed]
- Helen, D.; Kim, H.; Tytgat, B.; Anne, W. Highly Diverse NirK Genes Comprise Two Major Clades That Harbour Ammonium-Producing Denitrifiers. BMC Genom. 2016, 17, 155. [Google Scholar] [CrossRef] [Green Version]
- Merkle, A.C.; Lehnert, N. Binding and Activation of Nitrite and Nitric Oxide by Copper Nitrite Reductase and Corresponding Model Complexes. Dalton Trans. 2012, 41, 3355–3368. [Google Scholar] [CrossRef] [PubMed]
- Nojiri, M.; Xie, Y.; Inoue, T.; Yamamoto, T.; Matsumura, H.; Kataoka, K.; Deligeer; Yamaguchi, K.; Kai, Y.; Suzuki, S. Structure and Function of a Hexameric Copper-Containing Nitrite Reductase. Proc. Natl. Acad. Sci. USA 2007, 104, 4315–4320. [Google Scholar] [CrossRef] [Green Version]
- Opperman, D.J.; Murgida, D.H.; Dalosto, S.D.; Brondino, C.D.; Ferroni, F.M. A Three-Domain Copper-Nitrite Reductase with a Unique Sensing Loop. IUCrJ 2019, 6, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, A.; Ishikawa, R.; Koteishi, H.; Tange, K.; Fukuda, Y.; Kobayashi, K.; Inoue, T.; Nojiri, M. Structural and Mechanistic Insights into the Electron Flow through Protein for Cytochrome C-Tethering Copper Nitrite Reductase. J. Biochem. 2013, 154, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Antonyuk, S.V.; Cong, H.; Eady, R.R.; Hasnain, S.S. Structures of Protein-Protein Complexes Involved in Electron Transfer. Nature 2013, 496, 123–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, D.; Watanabe, T.F.; Eady, R.R.; Garratt, R.C.; Antonyuk, S.V.; Hasnain, S.S. Reverse Protein Engineering of a Novel 4-Domain Copper Nitrite Reductase Reveals Functional Regulation by Protein-Protein Interaction. FEBS J. 2020. [Google Scholar] [CrossRef]
- Giménez, M.I.; Dilks, K.; Pohlschröder, M. Haloferax Volcanii Twin-Arginine Translocation Substates Include Secreted Soluble, C-Terminally Anchored and Lipoproteins. Mol. Microbiol. 2007, 66, 1597–1606. [Google Scholar] [CrossRef] [PubMed]
- Storf, S.; Pfeiffer, F.; Dilks, K.; Chen, Z.Q.; Imam, S.; Pohlschröder, M. Mutational and Bioinformatic Analysis of Haloarchaeal Lipobox-Containing Proteins. Archaea 2010, 2010, e410975. [Google Scholar] [CrossRef] [Green Version]
- Esclapez, J.; Zafrilla, B.; Martínez-Espinosa, R.M.; Bonete, M.J. Cu-NirK from Haloferax mediterranei as an Example of Metalloprotein Maturation and Exportation via Tat System. Biochim. Biophys. Acta 2013, 1834, 1003–1009. [Google Scholar] [CrossRef]
- Hoehn, G.T.; Clark, V.L. The Major Anaerobically Induced Outer Membrane Protein of Neisseria gonorrhoeae, Pan 1, Is a Lipoprotein. Infect. Immun. 1992, 60, 4704–4708. [Google Scholar] [CrossRef] [Green Version]
- Boulanger, M.J.; Murphy, M.E.P. Crystal Structure of the Soluble Domain of the Major Anaerobically Induced Outer Membrane Protein (AniA) from Pathogenic Neisseria: A New Class of Copper-Containing Nitrite Reductases. J. Mol. Biol. 2002, 315, 1111–1127. [Google Scholar] [CrossRef] [PubMed]
- Sikora, A.E.; Mills, R.H.; Weber, J.V.; Hamza, A.; Passow, B.W.; Romaine, A.; Williamson, Z.A.; Reed, R.W.; Zielke, R.A.; Korotkov, K.V. Peptide Inhibitors Targeting the Neisseria gonorrhoeae Pivotal Anaerobic Respiration Factor AniA. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Jones, C.M.; Stres, B.; Rosenquist, M.; Hallin, S. Phylogenetic Analysis of Nitrite, Nitric Oxide, and Nitrous Oxide Respiratory Enzymes Reveal a Complex Evolutionary History for Denitrification. Mol. Biol. Evol. 2008, 25, 1955–1966. [Google Scholar] [CrossRef]
- Boulanger, M.J.; Kukimoto, M.; Nishiyama, M.; Horinouchi, S.; Murphy, M.E. Catalytic Roles for Two Water Bridged Residues (Asp-98 and His-255) in the Active Site of Copper-Containing Nitrite Reductase. J. Biol. Chem. 2000, 275, 23957–23964. [Google Scholar] [CrossRef] [Green Version]
- Antonyuk, S.V.; Strange, R.W.; Sawers, G.; Eady, R.R.; Hasnain, S.S. Atomic Resolution Structures of Resting-State, Substrate- and Product-Complexed Cu-Nitrite Reductase Provide Insight into Catalytic Mechanism. Proc. Natl. Acad. Sci. USA 2005, 102, 12041–12046. [Google Scholar] [CrossRef] [Green Version]
- Lawton, T.J.; Bowen, K.E.; Sayavedra-Soto, L.A.; Arp, D.J.; Rosenzweig, A.C. Characterization of a Nitrite Reductase Involved in Nitrifier Denitrification. J. Biol. Chem. 2013, 288, 25575–25583. [Google Scholar] [CrossRef] [Green Version]
- Kukimoto, M.; Nishiyama, M.; Murphy, M.E.P.; Turley, S.; Adman, E.T.; Horinouchi, S.; Beppu, T. X-Ray Structure and Site-Directed Mutagenesis of a Nitrite Reductase from Alcaligenes faecalis S-6: Roles of Two Copper Atoms in Nitrite Reduction. Biochemistry 1994, 33, 5246–5252. [Google Scholar] [CrossRef]
- Ellis, M.J.; Dodd, F.E.; Sawers, G.; Eady, R.R.; Hasnain, S.S. Atomic Resolution Structures of Native Copper Nitrite Reductase from Alcaligenes xylosoxidans and the Active Site Mutant Asp92Glu. J. Mol. Biol. 2003, 328, 429–438. [Google Scholar] [CrossRef]
- Hedison, T.M.; Shenoy, R.T.; Iorgu, A.I.; Heyes, D.J.; Fisher, K.; Wright, G.S.A.; Hay, S.; Eady, R.R.; Antonyuk, S.V.; Hasnain, S.S.; et al. Unexpected Roles of a Tether Harboring a Tyrosine Gatekeeper Residue in Modular Nitrite Reductase Catalysis. ACS Catal. 2019, 9, 6087–6099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kataoka, K.; Furusawa, H.; Takagi, K.; Yamaguchi, K.; Suzuki, S. Functional Analysis of Conserved Aspartate and Histidine Residues Located Around the Type 2 Copper Site of Copper-Containing Nitri Reductase. J. Biochem. 2000, 127, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Hodak, M.; Bernholc, J. Enzymatic Mechanism of Copper-Containing Nitrite Reductase. Biochemistry 2015, 54, 1233–1242. [Google Scholar] [CrossRef] [PubMed]
- Lintuluoto, M.; Lintuluoto, J.M. Intra-Electron Transfer Induced by Protonation in Copper-Containing Nitrite Reductase. Metallomics 2018, 10, 565–578. [Google Scholar] [CrossRef]
- Hallin, S.; Philippot, L.; Löffler, F.E.; Sanford, R.A.; Jones, C.M. Genomics and Ecology of Novel N2O-Reducing Microorganisms. Trends Microbiol. 2018, 26, 43–55. [Google Scholar] [CrossRef]
- Hein, S.; Simon, J. Chapter Four—Bacterial nitrous oxide respiration: Electron transport chains and copper transfer reactions. In Advances in Microbial Physiology; Poole, R.K., Ed.; Academic Press: Cambridge, MA, USA, 2019; Volume 75, pp. 137–175. [Google Scholar]
- Pauleta, S.R.; Carreira, C.; Moura, I. Chapter 7. Insights into Nitrous Oxide Reductase. In Metallobiology; Moura, I., Moura, J.J.G., Pauleta, S.R., Maia, L.B., Eds.; Royal Society of Chemistry: Cambridge, UK, 2016; pp. 141–169. ISBN 978-1-78262-334-2. [Google Scholar]
- Simon, J.; Einsle, O.; Kroneck, P.M.H.; Zumft, W.G. The Unprecedented Nos Gene Cluster of Wolinella Succinogenes Encodes a Novel Respiratory Electron Transfer Pathway to Cytochrome c Nitrous Oxide Reductase. 1054. FEBS Lett. 2004, 569, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Dell’Acqua, S.; Moura, I.; Moura, J.J.G.; Pauleta, S.R. The Electron Transfer Complex between Nitrous Oxide Reductase and Its Electron Donors. JBIC J. Biol. Inorg. Chem. 2011, 16, 1241–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, K.; Tegoni, M.; Prudêncio, M.; Pereira, A.S.; Besson, S.; Moura, J.J.; Moura, I.; Cambillau, C. A Novel Type of Catalytic Copper Cluster in Nitrous Oxide Reductase. Nat. Struct. Biol. 2000, 7, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, M.J.; Lafay, B.; Christen, R.; Fernandez, L.; Acquaviva, M.; Bonin, P.; Bertrand, J.-C. Marinobacter hydrocarbonoclasticus Gen. Nov., Sp. Nov., a New, Extremely Halotolerant, Hydrocarbon-Degrading Marine Bacterium. Int. J. Syst. Evol. Microbiol. 1992, 42, 568–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graziano, G.; Merlino, A. Molecular Bases of Protein Halotolerance. Biochim. Biophys. Acta BBA Proteins Proteom. 2014, 1844, 850–858. [Google Scholar] [CrossRef]
- Pauleta, S.R.; Dell’Acqua, S.; Moura, I. Nitrous Oxide Reductase. Coord. Chem. Rev. 2013, 257, 332–349. [Google Scholar] [CrossRef]
- Chen, P.; Gorelsky, S.I.; Ghosh, S.; Solomon, E.I. N2O Reduction by the Μ4-Sulfide-Bridged Tetranuclear CuZ Cluster Active Site. Angew. Chem. Int. Ed. 2004, 43, 4132–4140. [Google Scholar] [CrossRef]
- Chen, P.; Cabrito, I.; Moura, J.J.G.; Moura, I.; Solomon, E.I. Spectroscopic and Electronic Structure Studies of the Μ4-Sulfide Bridged Tetranuclear CuZ Cluster in N2O Reductase: Molecular Insight into the Catalytic Mechanism. 1070. J. Am. Chem. Soc. 2002, 124, 10497–10507. [Google Scholar] [CrossRef]
- Carreira, C.; Pauleta, S.R.; Moura, I. The Catalytic Cycle of Nitrous Oxide Reductase—The Enzyme That Catalyzes the Last Step of Denitrification. J. Inorg. Biochem. 2017, 177, 423–434. [Google Scholar] [CrossRef]
- Ghosh, S.; Gorelsky, S.I.; DeBeer George, S.; Chan, J.M.; Cabrito, I.; Dooley, D.M.; Moura, J.J.G.; Moura, I.; Solomon, E.I. Spectroscopic, Computational, and Kinetic Studies of the Μ4-Sulfide-Bridged Tetranuclear CuZ Cluster in N2O Reductase: pH Effect on the Edge Ligand and Its Contribution to Reactivity. J. Am. Chem. Soc. 2007, 129, 3955–3965. [Google Scholar] [CrossRef] [Green Version]
- Johnston, E.M.; Carreira, C.; Dell’Acqua, S.; Dey, S.G.; Pauleta, S.R.; Moura, I.; Solomon, E.I. Spectroscopic Definition of the CuZ° Intermediate in Turnover of Nitrous Oxide Reductase and Molecular Insight into the Catalytic Mechanism. J. Am. Chem. Soc. 2017, 139, 4462–4476. [Google Scholar] [CrossRef] [Green Version]
- Dell’Acqua, S.; Pauleta, S.R.; Moura, J.J.G.; Moura, I. Biochemical Characterization of the Purple Form of Marinobacter hydrocarbonoclasticus Nitrous Oxide Reductase. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 1204–1212. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Perchlorate | Chlorate | Bromate | Iodate | Nitrate | References | |
|---|---|---|---|---|---|---|
| AoPcrAB-Wild type | 6.0 ± 2.1 | 7.4 ± 2.1 | 4.4 ± 3.8 | 11 ± 5.4 | 23 ± 5.2 | [35] |
| AoPcrAB- PcrA W489E | 301 ± 14.1 | 20.5 ± 3.2 | 4.8 ± 2.1 | 1.27 × 105 ± 3.5 × 104 | 1.59 × 104 ± 4.5 × 103 | [35] |
| EcNarGHI | 1060 ± 154 | 113 ± 35 | 2690 ± 502 | N.S | 202 ± 65 | [35,52] |
| HmNarGH | Unquantified detected activity | 2.41 ± 0.16 | Unquantified detected activity | N.S | 0.82 ± 0.14 | [20,51] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernabeu, E.; Miralles-Robledillo, J.M.; Giani, M.; Valdés, E.; Martínez-Espinosa, R.M.; Pire, C. In Silico Analysis of the Enzymes Involved in Haloarchaeal Denitrification. Biomolecules 2021, 11, 1043. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11071043
Bernabeu E, Miralles-Robledillo JM, Giani M, Valdés E, Martínez-Espinosa RM, Pire C. In Silico Analysis of the Enzymes Involved in Haloarchaeal Denitrification. Biomolecules. 2021; 11(7):1043. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11071043
Chicago/Turabian StyleBernabeu, Eric, Jose María Miralles-Robledillo, Micaela Giani, Elena Valdés, Rosa María Martínez-Espinosa, and Carmen Pire. 2021. "In Silico Analysis of the Enzymes Involved in Haloarchaeal Denitrification" Biomolecules 11, no. 7: 1043. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11071043