
Enriched CO2 and Root-Associated Fungi (Mycorrhizae) Yield Inverse Effects on Plant Mass and Root Morphology in Six Asclepias Species



Department of Ecology and Evolutionary Biology, Kansas Biological Survey, 2101 Constant Ave, Lawrence, KS 66045, USA
*
Author to whom correspondence should be addressed.
Plants 2021, 10(11), 2474; https://0-doi-org.brum.beds.ac.uk/10.3390/plants10112474
Submission received: 17 September 2021
/
Revised: 2 November 2021
/
Accepted: 4 November 2021
/
Published: 16 November 2021
(This article belongs to the Special Issue The Effects of Mycorrhizal Symbiosis on Plant Development and Stress Tolerance)
Abstract
:While milkweeds (Asclepias spp.) are important for sustaining biodiversity in marginal ecosystems, CO2 flux may afflict Asclepias species and cause detriment to native communities. Negative CO2-induced effects may be mitigated through mycorrhizal associations. In this study, we sought to determine how mycorrhizae interacts with CO2 to influence Asclepias biomass and root morphology. A broad range of Asclepias species (n = 6) were chosen for this study, including four tap-root species (A. sullivantii, A. syriaca, A. tuberosa, and A. viridis) and two fibrous root species (A. incarnata and A. verticillata). Collectively, the six Asclepias species were manipulated under a 2 × 2 full-factorial design that featured two mycorrhizal levels (−/+ mycorrhizae) and two CO2 levels (ambient and enriched (i.e., 3.5× ambient)). After a duration of 10 months, Asclepias responses were assessed as whole dry weight (i.e., biomass) and relative transportive root. Relative transportive root is the percent difference in the diameter of highest order root (transportive root) versus that of first-order absorptive roots. Results revealed an asymmetrical response, as mycorrhizae increased Asclepias biomass by ~12-fold, while enriched CO2 decreased biomass by about 25%. CO2 did not impact relative transportive roots, but mycorrhizae increased root organ’s response by more than 20%. Interactions with CO2 and mycorrhizae were observed for both biomass and root morphology (i.e., relative transportive root). A gene associated with CO2 fixation (rbcL) revealed that the two fibrous root species formed a phylogenetic clade that was distant from the four tap-root species. The effect of mycorrhizae was most profound in tap-root systems, as mycorrhizae modified the highest order root into tuber-like structures. A strong positive correlation was observed with biomass and relative transportive root. This study elucidates the interplay with roots, mycorrhizae, and CO2, while providing a potential pathway for mycorrhizae to ameliorate CO2 induced effects.
1. Introduction
Plants have a broad range of traits that vary in response to environmental cues [1,2], including atmospheric carbon. CO2-induced responses (i.e., guard cell regulation and stomatal conductance) have long been characterized [3,4], while subsurface responses, particularly those pertaining to roots, have become increasingly controversial [5,6,7,8]. Roots are conduits for carbon flux, as these extensive organs can modulate soil chemistry, structure, and microbes [9,10,11,12]. At the root–soil interface, mycorrhizal associations are sustained through carbon–nutrient exchange [13,14], but a rise in atmospheric carbon may strain root-mediated traits [6,15], and eco-physiological function (i.e., root respiration, longevity, and turnover), which are central to root-mediated C cycling [16,17,18]. CO2 enrichment may increase root hierarchal branching, specific length, stele, cortex, and diameter [19,20], but it is unclear how root morphology varies in response to CO2, especially across root order [17] and system (tap- versus fibrous root system). Roots that correspond to the hyper-fibrous end of the tap- versus fibrous root trait continuum, are less mycorrhizal dependent [21], which can lead to negative interactions (i.e., competition) with mycorrhizal extramatrical hyphae [22]. Unraveling how mycorrhizae and atmospheric carbon may interact with roots can potentially provide insight into CO2-induced changes.
Root morphology and CO2 may interact with mycorrhizae. Upon C fixation in the shoot, carbon is translocated to roots and metabolized by mycorrhizae [23], but among glomeromycotans, CO2 enrichment may not necessarily lead to an increase in arbuscule development [24]. Arbuscules are confined to root cortical cells, and are important for carbon–nutrient exchange in colonized roots. Tap- and fibrous root systems are architecturally different, but have unique advantages. Fibrous roots are conducive toward water infiltration and reducing soil erosion [11,25], while tap-root systems excel at penetrating compacted soil layers [26]. In response to CO2 flux, fibrous and tap-root systems may vary in modes of nutrient foraging that are independent of mycorrhizal networks. Fibrous root systems are premier foragers at soil depths proximal to organic layers [27]. Meanwhile, tap-root systems are effective at exploiting deeper soil horizons that have accumulated leached nutrients (i.e., NO3−, SO42−), minerals [27,28], and ground water [29]. In comparison, fibrous root systems have high absorptive capacity [30,31] and specific root length (SRL) [32].
Root function is a product of root branching order [33], but still, root morphology may be impacted by CO2. An increase in atmospheric CO2 has been reported to improve lateral and tap-root sizes [34]. Tap-roots are of a higher root order in contrast to absorptive roots (i.e., root order 1 and 2), which are of a lower branching order [35]. Lower order roots are more involved in nutrient uptake and mycorrhizal associations. In contrast, higher order roots (i.e., root order > 3) have higher C–N [36], which may suggest a greater response to enriched CO2.
To date, very few studies have addressed the combinatorial effect of CO2 and mycorrhizae on roots, and even fewer studies have addressed these factors in the context of tap- versus fibrous root systems. Here, we address the following questions, (1) are the combinatorial effects of CO2 and mycorrhizae additive or interactive? (2) Does the outcome of CO2 and mycorrhizae differ in tap- versus fibrous root systems? Here, it is hypothesized that (a) the effect of CO2 and mycorrhizae will not be additive, as roots are likely to trade-away excess carbon to mycorrhizae. It is also hypothesized that, (b) the root response to mycorrhizae and CO2 will differ across the tap- versus fibrous root trait continuum, particularly due to difference in root architecture.
2. Material and Methods
2.1. Study System
The aim of this study was to determine ways in which CO2 and mycorrhizae can influence root morphology and plant performance (i.e., whole biomass). Thus, 6 species of milkweed from genus Asclepias were chosen for this experiment (Table 1). Milkweeds are excellent specimens for environmental manipulation (i.e., growth chamber) because they are adapted to a wide range of ecological niches [37], including disturbed, pristine, drought, and wetland habitats (Table 1). In addition, milkweed (hereafter referred to as Asclepias species) are important to marginal ecosystems (i.e., remnant prairie), biodiversity (i.e., monarch butterflies), and conservation interests.
2.2. Source of Asclepias spp. (Milkweed) and Mycorrhizae
Asclepias seeds were sourced from the midwestern region of the U.S.A. The 6 Asclepias spp. were A. sullivantii, A. verticillata (seed source, Prairie Moon Nursery Winona, MN, U.S.A), A. incarnata, A. syriaca, A. tuberosa, and A. viridis (seed source, Missouri Wild Flower Nursery (Jefferson City, MO, U.S.A). The relatedness of each of these species was characterized using a chloroplast DNA sequence that codes for ribulose bisphosphate carboxylase (rbcL), a conserved gene involved in CO2 fixation. Since Asclepias spp. response to (CO2) manipulation is the nature of this study, assessing Asclepias relatedness with respect to rbcL gene was appropriate.
Similar to Asclepias spp., mycorrhizal isolates were also sourced from the midwestern region of the U.S.A. Fungal consortia featured 4 isolates of mycorrhizae derived from Kankakee sands prairie reserve in Indiana, USA. Isolates included several species (Claroideoglomus claroideum, Racocetra fulgida, Funneliformis mosseae, and Claroideoglomus lamellosum) that have comparable levels of colonization [42]. In addition, these species are representative of mycorrhizal phylogenetic diversity [43,44], and include isolates of high and low experimental usage rate [45].
2.3. Preparing Mycorrhizal Inoculate
Mycorrhizae are lab isolates that were first isolated from prairie soil to start lab cultures (Table S1). Identity of spores were previously determined using morphological and phylogenetic species concept [43]. Since then, these isolates have been of standard use in the lab [42,46,47]. Prior to the experiment, fungal cultures were prepared and bulked on sorghum roots for a full growing season under glasshouse conditions [46]. Briefly, after a full growing season, aboveground sorghum tissue was removed, while the belowground soil and mycorrhizal root mix was stored at 4 °C prior to experimental use.
At the time of experimental use, mycorrhizal fungal inocula (fungal spores and sorghum root mix) was used to inoculate in between the top and bottom layers of heat sterile background soil (Kansas clay loam–sand (1:1)). The background soil was mixed with sand, primarily because sand enhances drainage in pots, reduces nutrient levels, and enhances plant’s mycorrhizal functional response. The volume of the added inocula (i.e., fungal spore–root mix) was 50 cm3 (+ mycorrhizae). Meanwhile, sterile inocula void of live mycorrhizae was added to control pots (− mycorrhizae), also at a volume of 50 cm3. This assured that the soil structure and nutrient ratio of all pots was maintained. Finally, Asclepias seedlings were transplanted to cone-tainer pots (Stuewe & Sons Inc., Tangent, Oregon, U.S.A.), which were positive or negative for mycorrhizae.
2.4. Growth Chamber and Atmospheric Conditions
Prior to transplanting milkweed to experimental conditions (i.e., −/+ mycorrhizae and CO2 manipulation), Asclepias spp. were germinated on heat sterile potting soil (Berger bark mix BM 7, www.berger.ca. access date: 9 September 2019) After five weeks, 5-week-old seedlings were transferred to pots that were positive or negative for mycorrhizae. The dimensions of the pots were 6.8 cm in diameter by a depth of 35.56 cm. In a random block design, pots were then placed into 4 Conviron CMP 6050 growth chambers (Controlled Environments Limited, Winnipeg, Manitoba, Canada). Two of the chambers were set to ambient (CO2) at 400 ppm, while the other two chambers were set to 3.75× ambient levels. The 3.75× ambient level (hereafter referred to as enriched CO2) was chosen because geological time records suggest (CO2) > 1500 ppm reduces plant stomatal index [48], which is of detriment to photosynthesis and Earth’s biosphere. Growth chamber settings were 23 °C, 70% humidity (% RH), and a lighting intensity of 1000 unit micromoles. CO2 concentrations were set to 400 ppm (ambient) and 1500 ppm (enriched). Plants were fertilized with nitrogen (0.2 g/L) periodically (average of once a month) at an application volume of 50 mL. Similar to Malik et al. (2016) [46], this (N) was determined and applied by using the atomic mass of N in the NH4NO3 compound, which was weighted to control for N mass. Using this ratio, we determined how much NH4NO3 is needed in 1 L of water to create 0.2 g/L of N.
2.5. Experimental Design
Experimental design featured 6 Asclepias spp. × 2 mycorrhizal levels (+/− mycorrhizae) × 2 CO2 conditions (ambient and 3.75× ambient) × 8 replicates per experimental treatment. In totality, the experiment was 192 observations (i.e., 192 cone-tainers). The experiment was performed under growth chamber conditions in a random block design. For 10 months, 6 Asclepias spp. were grown on −/+ mycorrhizal soil regimes, while under ambient and enriched (3.75× ambient) CO2 conditions. Plants were first germinated on heat sterile potting soil. After 5 weeks, seedlings were transplanted to −/+ mycorrhizal microcosms. The Asclepias–mycorrhizal microcosms were exposed to either ambient or enriched (CO2) for a duration of 10 months (December 2019–October 2020).
2.6. Harvest
After a duration of 10 months, the effects of mycorrhizae and CO2 on Asclepias spp. were observed. Roots were washed and scanned for root morphological responses. Briefly, roots were blot dried and positioned next to a scale, and an EPSON scan was used to make morphological observations. Roots were then stored in 75% ethanol and boiled in KOH, prior to trypan blue staining [49]. Staining and qualitative microscopic examination confirmed the mycorrhizal treatments. Finally, plant performance was assessed as whole dry weight.
2.7. Root Image Analysis
For image analysis, roots were washed and scanned to assess morphological response to CO2 and mycorrhizae. Root morphological response was quantified with ImageJ 1.46r [50], as pixels were calibrated to centimeters to assess diameter. Root diameter was scored at the midpoint of the highest order root (transport root), as well as the midpoint of the lowest order root (absorptive root). Percent difference in transport versus absorptive root diameter (relative transport root) was estimated.
Relative transport root, or percent difference between transport and absorptive root, was appropriate because root diameter has been shown to vary across plant groups [1], including tap- and fibrous root species. Relative transport root is calculated according to Equation 1, where transport (T) and absorptive (A) root diameters are divided by transport root (T) diameter. This value is then multiplied by 100.
2.8. Statistical Analysis for Root Morphology and Biomass
This study was analyzed in R version 4.0.2. Shapiro–Wilk and Levene’s Test, as well as diagnostic plots, were used to assess homogeneity in variance and normality for dependent variables (i.e., relative transport root and biomass). CO2 level, mycorrhizae, species and growth chamber effects were set as predictors, while both relative transportive root and biomass were set as response variables. MANOVA analysis was employed, as relative transport root and biomass were response variables from the same set of plants. Briefly, for MANOVA analysis, relative transport root and biomass were made into a vector, and simultaneous inferences were made by the predictor variables. The analysis was followed by a set of ANOVAs that independently predict biomass and root morphology. Interactions among the predictors were also included in the model, this was critical for determining whether or not growth chamber(s) were a confounding factor. Post-hoc analysis included Step AICc, Tukey HSD, and a priori contrast using the ‘multcomp’ package. The ‘multcomp’ package allowed simultaneous test for general linear hypotheses [51]. Finally, a correlation between relative transport root and biomass was assessed using the ‘rcorr’ and ‘cor.test’ function via ‘Hmisc’ package [52]. Correlations were then examined with 95% CI ellipses, using ‘ggplot 2’. The ellipses enabled graphical analysis for outcome and experimental variables
2.9. Phylogenetic Analysis: Asclepias Relatedness Using CO2 Fixation Gene
Amino acid (AA) sequence for rbcL locus was aligned using Muscle, which assesses distance using kmer distance for unaligned pairs and kimura distance for aligned pairs (Edgar, 2004). Evolutionary analysis was inferred using Maximum Likelihood Method and the JTT matrix-based model [53]. The phylogenetic tree for the 6 Asclepias species was constructed according to the highest log likelihood. This enabled a tree that was drawn to scale with branch lengths measured as the number of substitutions per site. While this analysis involved 6 AA sequences, there were a total of 475 positions in the final dataset. This allowed evolutionary analysis to be conducted with MEGA X [54].
3. Results
3.1. Multivariate Analysis, Qualitative Observations, and Root System
The viability and effectiveness of the inoculum was confirmed after 10 months, as root–mycorrhizal structures (i.e., arbuscules and vesicles) were observed to be present in experimental roots (i.e., + Mycorrhizae), while the presence of these structures was undetected in the control. MANOVA analysis revealed that the combination of root diameter and biomass significantly responded to CO2 (Pillai 1143 = 0.057, p = 0.01) and mycorrhizae (Pillai 1143 = 0.064, p < 0.0001). Additionally, the combination of these traits led to significant interactions with CO2 and mycorrhizae (MANOVA, Pillai 1143 = 0.051, p = 0.023), CO2 and Asclepias species (MANOVA, Pillai 5143 = 0.140, p = 0.020), and mycorrhizae and Asclepias species (MANOVA, Pillai 5143 = 0.613, p < 0.0001). Both fibrous root species and tap-root species were enhanced by mycorrhizae (Figure 1). In tap-root systems, dominant pronounced roots (i.e., higher order transport roots) were likely to be modified into tuber-like structures while in the presence of mycorrhizae (Figure 1). A priori contrast revealed significant biomass difference in species with tap- versus fibrous root systems. The difference was about 7 g (95% CI [−∞, −2.576], p = 0.005, ANOVA, simultaneous tests for GLH, fibrous versus tap-root system). Similarly, root systems differed in relative transport roots by an estimate of 25.7% (95% CI [−∞, −3.459], p = 0.028, ANOVA, simultaneous tests for GLH, fibrous versus tap-root system).
3.2. Biomass Response to Mycorrhizae and CO2
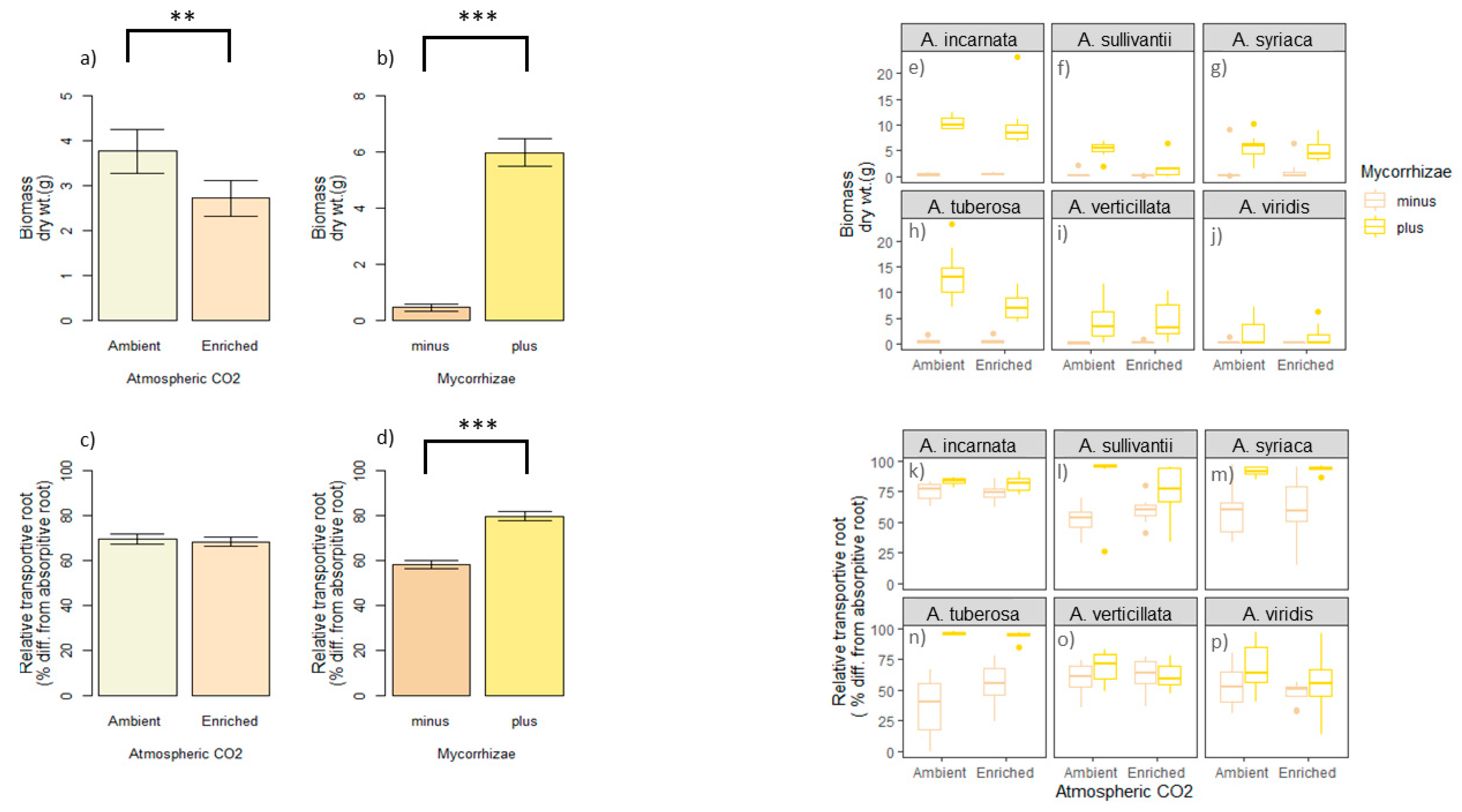
Biomass (i.e., dry weight) was decreased under enriched atmospheric CO2 by ~25% (Figure 2a, ANOVA, F 1143 = 8.525, p = 0.004). In contrast, mycorrhizae significantly improved biomass by ~12-fold (Figure 2b, ANOVA F 1143 = 235.161, p < 0.0001,). Increase in biomass was broadly observed across the six Asclepias species (Figure 2e–j). Interestingly, biomass response was also influenced by CO2 and mycorrhizae interaction (ANOVA F 1143 = 5.690, p = 0.01, Table S2), as well as CO2 and Asclepias species interaction (ANOVA, F 5143 = 8.021, p = 0.03, Table S2).
3.3. Transport Root Response to Mycorrhizae and CO2
With regard to the relative transport root, CO2 did not play a significant role (Figure 2c, ANOVA, F 1143 = 0.228, p = 0.63, Table S3), while mycorrhizae did (Figure 2d, F 1143 = 91.881, p < 0.0001, Table S3). Specifically, mycorrhizae improved relative transport root diameter by about 20% (Figure 2d). The relative transport root was also significantly impacted by a CO2 and mycorrhizae interaction (F 1144 = 4.580, p = 0.034, Table S3), as well as mycorrhizae and Asclepias species interaction (F 1143 = 9.527, p < 0.0001, Table S3). In general, the relative transport root differed across Asclepias spp. (Figure 2k–p, F 5143 = 8.937, p < 0.0001).
3.4. Correlation with Relative Transport and Biomass
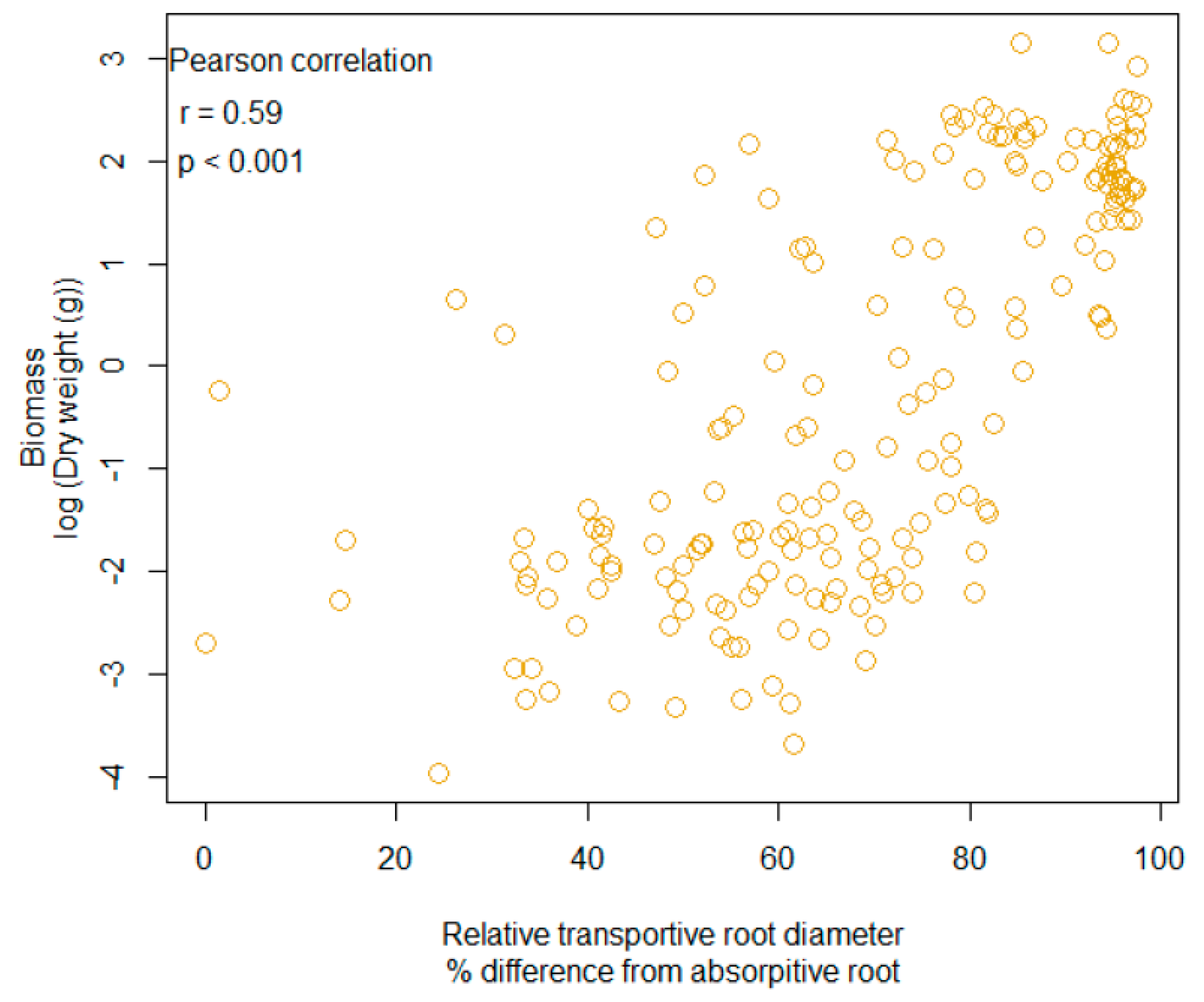
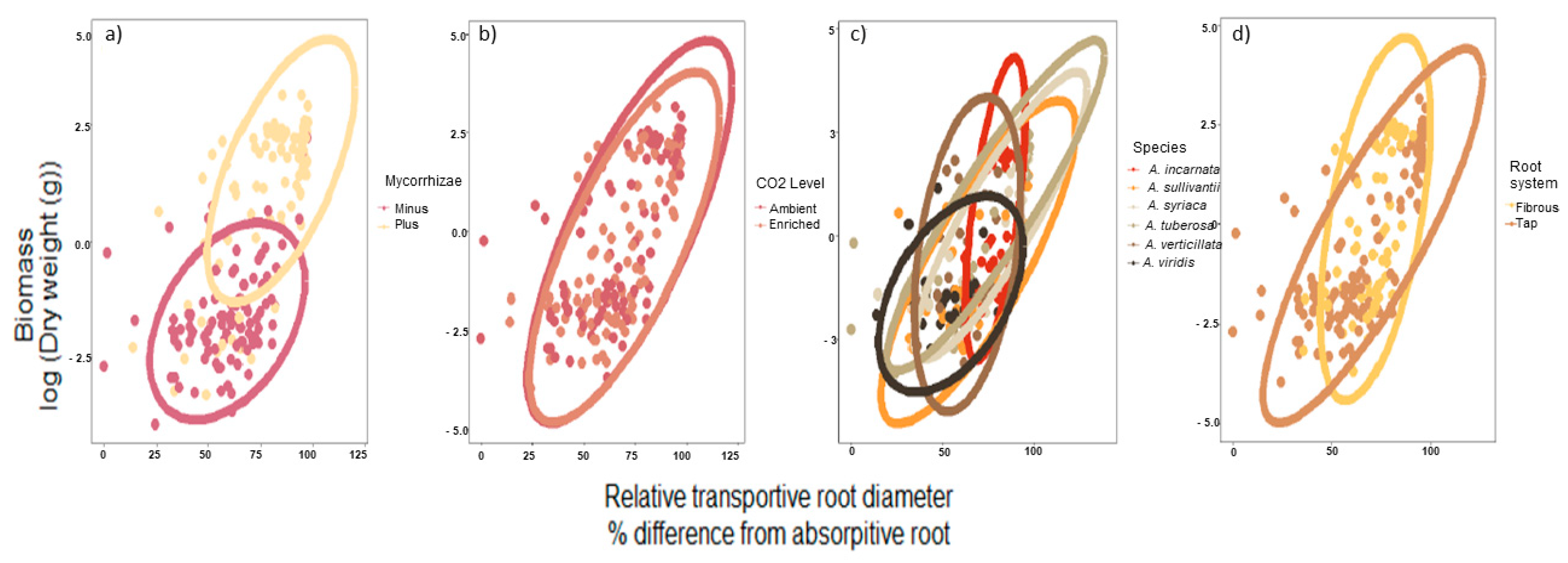
A significant correlation was also detected with biomass and root diameter, as these two responses were positively associated (Pearson correlation, r = 0.59, n = 191, p < 0.001, Figure 3). In addition, 95% CI ellipse revealed that the association with relative transport root and biomass was increasingly positive in the presence of mycorrhizae, which led to a small overlap between + mycorrhizae and − mycorrhizae (Figure 4a). The CO2 regime (i.e., ambient versus enriched) produced a 95% CI ellipse with a high degree of overlap and similar shape. However, the CO2 ellipses were not congruent as the ambient CO2 ellipse was more positive (Figure 4b). As it relates to species, both fibrous root species, A. incarnata and A. verticillata produced ellipses shapes that were unique and not angled at 45 degrees, unlike the tap-root species (Figure 4c). These differences are even more apparent when ellipses are assorted by root system (Figure 4d).
3.5. Asclepias Species Relatedness and Root System
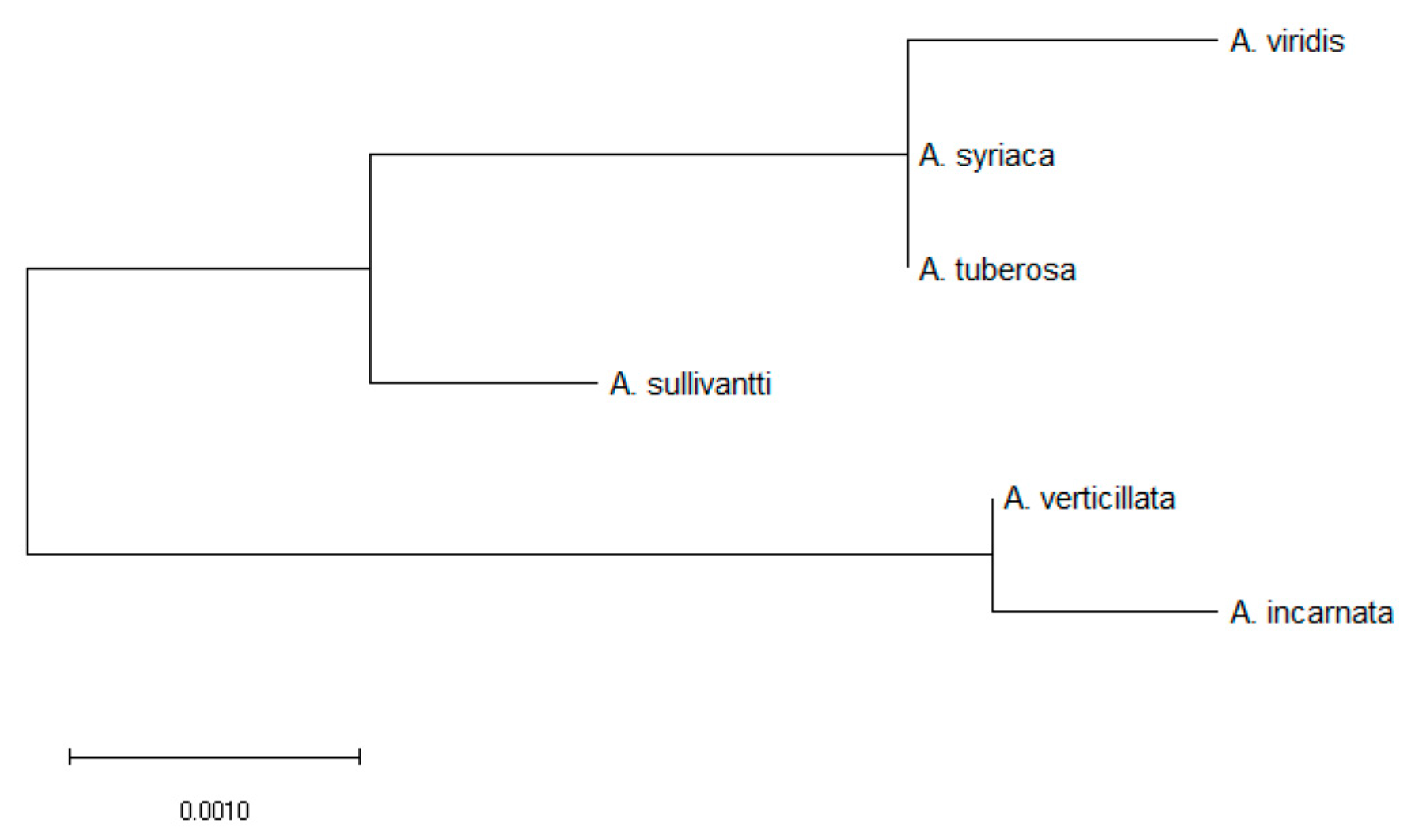
With respect to the rbcL locus (i.e., gene involved in CO2 fixation), a tree was constructed to the highest log likelihood (−1398.00) to examine relatedness of the six Asclepias species. Interestingly, the root system helped explain relatedness. Specifically, the two fibrous root species, A. incarnata and A. verticillata were sister taxa, and an outgroup, that was distant to the four other tap-root species (Figure 5).
3.6. Chamber Effects
Predictors (i.e., CO2, mycorrhizae, Asclepias species) did not interact with CO2 chambers (Tables S2 and S3, p > 0.05). This provides confidence that results were not confounded by the experimental apparatus. In addition, Step AICc revealed that response variables including biomass and relative transport root were best explained when growth chamber interactions were removed from the model.
4. Discussion
Enriched CO2 and mycorrhizae may be additive or opposing forces helping shape plant eco-physiological responses. CO2 flux is relevant to root life span, production, and diameter [55,56,57], as well as ecosystem processes [58,59,60], and plant fitness [61]. Interestingly, the targeted role of roots may be enhanced by mycorrhizae [62], but our findings reveal that enriched CO2 reduced plant biomass by 25% (Figure 2a). We speculate that the mechanism at play may be CO2 regulation of stomatal density, conductance, and aperture [63], as enriched CO2 can lead to stomatal closures, which can reduce carbon capture and net CO2 assimilation [64,65]. Together, this may explain negative CO2-induced effects on Asclepias spp., particularly biomass. Contrarily, mycorrhizae improved biomass by 12-fold while modifying root morphology (Figure 1 and Figure 2b,d). Significant interactions with CO2 and mycorrhizae were detected for biomass and relative transport roots (Tables S2 and S3). This study elucidates potential mechanisms in which CO2 and mycorrhizae may yield asymmetrical outcomes on plant eco-physiology.
4.1. Biomass, CO2 and Mycorrhizae
Carbon is captured from the atmosphere, and used to build biomass and organs (i.e., roots), but this would not be sustainable without root absorption, subsurface foraging, nutrient transport, and organic content storage [66,67,68]. While root functionality can be stimulated by CO2 [69], enriched CO2 can have negative outcomes on fine lateral roots and plant mass in agroecosystems [70,71]. As it relates to Asclepias species, which are essential to specialist herbivores (e.g., monarch butterflies), biodiversity, and prairie ecosystems; enriched CO2 decreased plant biomass by 25% (Figure 2a). These differences were most apparent in five of the six Asclepias species (A. incarnata, A tuberosa, A. sullivantii, A. viridis, A. syriaca, Figure 2e–j (but not Figure 2i)). Fibrous root species, A. verticillata, was the only species in which the combination of mycorrhizae and enriched CO2 was observed to have an additive effect on biomass (Figure 2i). This may be explained by the fact that A. verticillata is considered to have some of the most divergent traits in genus Asclepias [39]. Hence, the disparate thin pointy leaves and very fine shallow grass-like roots makes A. verticillata distinct, and may have a role in explaining A. verticillata’s unique response to mycorrhizae and CO2.
Interestingly, the CO2 effect on biomass and root morphology may be context dependent. At a CO2 level of 650 ppm, plant growth and percent mycorrhizal colonization was reportedly increased [72]. Perhaps suggesting that when CO2 stimulates plant growth, percent mycorrhizal colonization is also stimulated. While percent mycorrhizal colonization was not quantified in this study, it may be the case that since enriched CO2 depressed plant growth (Figure 2a), percent mycorrhizal colonization was also depressed. This may help explain the significant interaction with CO2 and mycorrhizae on biomass (Table S2). However, the outcome of CO2 on plant–mycorrhizal nutrient exchange may be species or cultivar specific [24].
4.2. CO2–Mycorrhizae Interaction and Root Response
Although elevated CO2 can lead to roots of high tissue density [73], root morphology has also been found to be associated with aboveground traits [1]. However, the precise mechanism as to how aboveground environmental cues affect belowground physiology remains obscure. In the present study, CO2 did not influence root morphology (Figure 2c), as these findings differed from a recent study [69]. It may be the case that CO2 can indirectly affect root morphology by increasing mycorrhizal root length colonization [74]. Interestingly, this hypothesis would rely on root systems that are highly responsive to mycorrhizae, as well as atmospheric–edaphic environment interactions.
Irrespective of the main effect of CO2, we observed an interaction with CO2 and mycorrhizae on root response (p = 0.03, Table S3). This may provide insight as to why the effect of plus mycorrhizal treatment depended on the CO2 level in A. verticillata roots. Specifically, mycorrhizae and enriched CO2 depressed A. verticillata’s relative transport root (Figure 2o). This sort of interaction was not observed in A. incarnata, the other fibrous root species. Differences among these two fibrous root species (i.e., A. verticillata and A. incarnata) may be due to divergent life histories and localized adaptation. In particular, A. verticillata thrives in xeric soils, while A. incarnata thrives in hydric soils (Table 1). Despite this, A. incarnata and A. verticillata (two fibrous root species), were more closely related than the other four tap-root species.
Tap- versus fibrous root systems may have been a good predictor of Asclepias spp. relatedness, as this observation does not appear to be confounded by rhizome trait(s). Hence, when rhizome traits were mapped onto a pre-existing phylogeny [75,76], it was determined that Asclepias spp. relatedness did not assort according to rhizome morphology. However, as it relates to the present study, Asclepias spp. assort according to the root system (Figure 5), as relatedness was characterized using the rbcL locus (Table S4)
4.3. Mycorrhizae, Root Modification, and Carbon Storage
Root morphology can vary across plant groups (i.e., lilies, forbes, grasses) of the same community [1]. This may be explained by root metabolomics [77], which have been shown to be altered by mycorrhizae [78]. Despite this, mycorrhizal implications may depend on the root system and morphology [21]. The outcome of mycorrhizae on root morphology was most exaggerated in tap-root systems (Figure S1), irrespective of [CO2]. In most cases, tap-roots were modified into tuber-like structures (Figure 1), perhaps re-iterating the notion that tap-root systems are more likely to be mycorrhizal responsive. In contrast, fibrous root systems may antagonize mycorrhizae, as fibrous root systems have been shown to have negative effects on mycorrhizal hyphal length [22]. In addition, mycorrhizal dependence has been reported to decrease with the increase in root fibrousness [21].
As it relates to the ecosystem function, mycorrhizae may use different mechanisms to promote carbon storage depending on the plant root system. Our findings suggest that in tap-root systems, mycorrhizae may directly promote root carbon storage by helping to modify tap-roots into tuber-like structures (Figure 1). However, mycorrhizae may complement fine roots by effectively promoting microaggregate stabilization [22], which can affect C sequestration and nutrient cycling [79,80,81].
5. Conclusions
Mycorrhizae enhanced relative transportive root and biomass, while enriched CO2 had the opposite effect. Interactions with CO2 and mycorrhizae were observed for both biomass and relative transport root. The findings of this study shed insight on how CO2 and mycorrhizae may interact and influence plant eco-physiology. In particular excess CO2 (i.e., 1500 ppm) can depress plant productivity, while mycorrhizae can act as a countering force to improve productivity.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/plants10112474/s1, Table S1. Code of Isolates. Table S2. Biomass explained by CO2, mycorrhizae, species, and growth chamber. Significant codes are for alpha thresholds. Table S3. Root morph. explained by CO2, mycorrhizae, species and growth chamber. Significant codes are for alpha thresholds. Figure S1 The effect of CO2 and mycorrhizae on fibrous root systems. Irrespective of CO2 regime, +mycorrhizae yielded a larger effect on tap-root systems. Table S4. GenBank Accession table.
Author Contributions
R.J.M. and J.D.B. conceived the ideas and concepts. R.J.M. performed the experiment, made observations, handled the data, performed analysis and wrote the manuscript. J.D.B. provided resources and materials, and helped with final edits. All authors have read and agreed to the published version of the manuscript.
Funding
Funding support provided by NSF-PRFB Award #190242 and NSF Award #2027458.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data shared upon request.
Acknowledgments
Special thanks to Suzanne M. Fleishman for helpful discussions. In addition, thank you to the reviewers at Plants.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhou, M.; Bai, W.; Li, Q.; Guo, Y.; Zhang, W. Root anatomical traits determined leaf-level physiology and responses to precipitation change of herbaceous species in a temperate steppe. New Phytol. 2021, 229, 1481–1491. [Google Scholar] [CrossRef] [PubMed]
- Henn, J.J.; Buzzard, V.; Enquist, B.J.; Halbritter, A.H.; Klanderud, K.; Maitner, B.S.; Michaletz, S.T.; Pötsch, C.; Seltzer, L.; Telford, R.J. Intraspecific trait variation and phenotypic plasticity mediate alpine plant species response to climate change. Front. Plant. Sci. 2018, 9, 1548. [Google Scholar] [CrossRef] [PubMed]
- Assmann, S. The cellular basis of guard cell sensing of rising CO2. Plant Cell Environ. 1999, 22, 629–637. [Google Scholar] [CrossRef] [Green Version]
- Purcell, C.; Batke, S.; Yiotis, C.; Caballero, R.; Soh, W.; Murray, M.; Mcelwain, J.C. Increasing stomatal conductance in response to rising atmospheric CO2. Ann. Bot. 2018, 121, 1137–1149. [Google Scholar] [CrossRef] [Green Version]
- Zak, D.; Pregitzer, K.S.; King, J.S.; Holmes, W.E. Elevated atmospheric CO2, fine roots and the response of soil microorganisms: A Review and hypothesis. New Phytol. 2000, 147, 201–222. [Google Scholar] [CrossRef] [Green Version]
- Drigo, B.; Pijl, A.S.; Duyts, H.; Kielak, A.M.; Gamper, H.A.; Houtekamer, M.J.; Boschker, H.T.; Bodelier, P.L.; Whiteley, A.S.; Van veen, J.A. Shifting carbon flow from roots into associated microbial communities in response to elevated atmospheric CO2. Proc. Natl. Acad. Sci. USA 2010, 107, 10938–10942. [Google Scholar] [CrossRef] [Green Version]
- Nelson, L.; Blumenthal, D.M.; Williams, D.G.; Pendall, E. Digging into the roots of belowground carbon cycling following seven years of Prairie Heating and CO2 Enrichment (PHACE), Wyoming USA. Soil Biol. Biochem. 2017, 115, 169–177. [Google Scholar] [CrossRef]
- Malik, R.J. No “Gadgil effect”: Temperate tree roots and soil lithology are effective predictors of wood decomposition. For. Pathol. 2019, 49, e12506. [Google Scholar] [CrossRef]
- Qadir, M.; Noble, A.; Oster, J.; Schubert, S.; Ghafoor, A. Driving forces for sodium removal during phytoremediation of calcareous sodic and saline–sodic soils: A review. Soil Use Manag. 2005, 21, 173–180. [Google Scholar] [CrossRef]
- Brady, N.C.; Weil, R.R. The Nature and Properties of Soils; Prentice Hall: Upper Saddle River, NJ, USA, 2008. [Google Scholar]
- Li, C.; Pan, C. The relative importance of different grass components in controlling runoff and erosion on a hillslope under simulated rainfall. J. Hydrol. 2018, 558, 90–103. [Google Scholar] [CrossRef]
- Oldroyd, G.E.D. Speak, friend, and enter: Signalling systems that promote beneficial symbiotic associations in plants. Nat. Rev. Microbiol. 2013, 11, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Shi, J.; Xie, Q.; Jiang, Y.; Yu, N.; Wang, E. Nutrient Exchange and Regulation in Arbuscular Mycorrhizal Symbiosis. Mol. Plant 2017, 10, 1147–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakobsen, I.; Rosendahl, L. Carbon flow into soil and external hyphae from roots of mycorrhizal cucumber plants. New Phytol. 1990, 115, 77–83. [Google Scholar] [CrossRef]
- Treseder, K.K.; Allen, M.F. Mycorrhizal fungi have a potential role in soil carbon storage under elevated CO2 and nitrogen deposition. New Phytol. 2000, 147, 189–200. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Fujita, S.; Nakaji, T.; Watanabe, M.; Satoh, F.; Koike, T. Fine root turnover of Japanese white birch (Betula platyphylla var. japonica) grown under elevated CO2 in northern Japan. Trees 2016, 30, 363–374. [Google Scholar] [CrossRef]
- Cohen, I.; Rapaport, T.; Berger, R.T.; Rachmilevitch, S. The effects of elevated CO2 and nitrogen nutrition on root dynamics. Plant Sci. 2018, 272, 294–300. [Google Scholar] [CrossRef]
- Suseela, V.; Tharayil, N.; Pendall, E.; Rao, A.M. Warming and elevated CO2 alter the suberin chemistry in roots of photosynthetically divergent grass species. AoB Plants 2017, 9, plx041. [Google Scholar] [CrossRef]
- Eissenstat, D.; Wells, C.; Yanai, R.; Whitbeck, J. Building roots in a changing environment: Implications for root longevity. New Phytol. 2000, 147, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Rogers, H.H.; Peterson, C.M.; McCrimmon, J.N.; Cure, J.D. Response of plant roots to elevated atmospheric carbon dioxide. Plant Cell Environ. 1992, 15, 749–752. [Google Scholar] [CrossRef]
- Hetrick, B.; Wilson, G.; Todd, T. Relationships of mycorrhizal symbiosis, rooting strategy, and phenology among tallgrass prairie forbs. Can. J. Bot. 1992, 70, 1521–1528. [Google Scholar] [CrossRef]
- Jastrow, J.; Miller, R.; Lussenhop, J. Contributions of interacting biological mechanisms to soil aggregate stabilization in restored prairie. Soil Biol. Biochem. 1998, 30, 905–916. [Google Scholar] [CrossRef]
- Bécard, G.; Piché, Y. Fungal growth stimulation by CO2 and root exudates in vesicular-arbuscular mycorrhizal symbiosis. Appl. Environ. Microbiol. 1989, 55, 2320–2325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thirkell, T.J.; Pastok, D.; Field, K.J. Carbon for nutrient exchange between arbuscular mycorrhizal fungi and wheat varies according to cultivar and changes in atmospheric carbon dioxide concentration. Glob. Chang. Biol. 2020, 26, 1725–1738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Wang, Z.; Guo, Q.; Lian, J.; Chen, L. Increase and Spatial Variation in Soil Infiltration Rates Associated with Fibrous and Tap Tree Roots. Water 2019, 11, 1700. [Google Scholar] [CrossRef] [Green Version]
- Hamza, M.; Anderson, W. Soil compaction in cropping systems: A review of the nature, causes and possible solutions. Soil Tillage Res. 2005, 82, 121–145. [Google Scholar] [CrossRef]
- Giehl, R.F.; Von Wirén, N. Root nutrient foraging. Plant. Physiol. 2014, 166, 509–517. [Google Scholar] [CrossRef] [Green Version]
- Jobbagy, E.G.; Jackson, R.B. The distribution of soil nutrients with depth: Global patterns and the imprint of plants. Biogeochemistry 2001, 53, 51–77. [Google Scholar] [CrossRef]
- Wilcox, B.P.; Owens, M.K.; Dugas, W.A.; Ueckert, D.N.; Hart, C.R. Shrubs, streamflow, and the paradox of scale. Hydrol. Process. An. Int. J. 2006, 20, 3245–3259. [Google Scholar] [CrossRef]
- Himmelbauer, M.L.; Puschenreiter, M.; Schnepf, A.; Loiskandl, W.; Wenzel, W.W. Root morphology of Thlaspi goesingense Hálácsy grown on a serpentine soil. J. Plant. Nutr. Soil Sci. 2005, 168, 138–144. [Google Scholar] [CrossRef]
- Leonard, E.D.; Monaco, T.A.; Stark, J.M.; Ryel, R.J. Invasive forb, annual grass, and exotic shrub competition with three sagebrush-steppe growth forms: Acquisition of a spring 15N tracer. Invasive Plant. Sci. Manag. 2008, 1, 168–177. [Google Scholar] [CrossRef]
- Eissenstat, D.M. Costs and benefits of constructing roots of small diameter. J. Plant Nutr. 1992, 15, 763–782. [Google Scholar] [CrossRef]
- Long, Y.; Kong, D.; Chen, Z.; Zeng, H. Variation of the linkage of root function with root branch order. PLoS ONE 2013, 8, e57153. [Google Scholar]
- Prior, S.A.; Rogers, H.H.; Runion, G.B.; Kimball, B.A.; Mauney, J.R.; Lewin, K.F.; Nagy, J.; Hendrey, G.R. Free-Air Carbon Dioxide Enrichment of Cotton: Root Morphological Characteristics. J. Environ. Qual. 1995, 24, 678–683. [Google Scholar] [CrossRef]
- King, W.L.; Yates, C.F.; Guo, J.; Fleishman, S.M.; Trexler, R.V.; Centinari, M.; Bell, T.H.; Eissenstat, D.M. The hierarchy of root branching order determines bacterial composition, microbial carrying capacity and microbial filtering. Commun. Biol. 2021, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lavely, E.K.; Chen, W.; Peterson, K.A.; Klodd, A.E.; Volder, A.; Marini, R.P.; Eissenstat, D.M. On characterizing root function in perennial horticultural crops. Am. J. Bot. 2020, 107, 1214–1224. [Google Scholar] [CrossRef] [PubMed]
- Woodson, R.E. The North American Species of Asclepias L. Ann. Mo. Bot. Gard. 1954, 41, 1–211. [Google Scholar]
- Price, P.W.; Wilson, M.F. Abundance of Herbivores on Six Milkweed Species in Illinois. Am. Midl. Nat. 1979, 101, 76–86. [Google Scholar] [CrossRef]
- Wilbur, H.M. Life History Evolution in Seven Milkweeds of the Genus Asclepias. J. Ecol. 1976, 64, 223–240. [Google Scholar] [CrossRef]
- Lucansky, T.W.; Clough, K.T. Comparative Anatomy and Morphology of Asclepias perennis and Asclepias tuberosa Subspecies Rolfsii. Int. J. Plant Sci. 1986, 147, 290–301. [Google Scholar] [CrossRef]
- Ali, J.G.; Agrawal, A.A. Trade-offs and tritrophic consequences of host shifts in specialized root herbivores. Funct. Ecol. 2017, 31, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Vogelsang, K.M.; Reynolds, H.L.; Bever, J.D. Mycorrhizal fungal identity and richness determine the diversity and productivity of a tallgrass prairie system. New Phytol. 2006, 172, 554–562. [Google Scholar] [CrossRef] [PubMed]
- House, G.L.; Ekanayake, S.; Ruan, Y.; Schütte, U.M.; Kaonongbua, W.; Fox, G.; Ye, Y.; Bever, J.D. Phylogenetically structured differences in rRNA gene sequence variation among species of arbuscular mycorrhizal fungi and their implications for sequence clustering. Appl. Environ. Microbiol. 2016, 82, 4921–4930. [Google Scholar] [CrossRef] [Green Version]
- Krüger, M.; Krüger, C.; Walker, C.; Stockinger, H.; Schüßler, A. Phylogenetic reference data for systematics and phylotaxonomy of arbuscular mycorrhizal fungi from phylum to species level. New Phytol. 2012, 193, 970–984. [Google Scholar] [CrossRef]
- Malik, R.J. Recent Trend: Is the Role of Arbuscular Mycorrhizal Fungi in Plant-Enemies Performance Biased by Taxon Usage? Am. Midl. Nat. 2018, 180, 306–312. [Google Scholar] [CrossRef]
- Malik, R.J.; Dixon, M.H.; Bever, J.D. Mycorrhizal composition can predict foliar pathogen colonization in soybean. Biol. Control. 2016, 103, 46–53. [Google Scholar] [CrossRef]
- Malik, R.J.; Ali, J.; Bever, J.D. Mycorrhizal composition influences plant anatomical defense and impacts herbivore growth and survival in a life-stage dependent manner. Pedobiologia 2018, 66, 29–35. [Google Scholar] [CrossRef]
- Retallack, G.J.; Conde, G.D. Deep time perspective on rising atmospheric CO2. Glob. Planet. Chang. 2020, 189, 103177. [Google Scholar] [CrossRef]
- Ambler, J.R.; Young, J. Techniques for determining root length infected by vesicular-arbuscular mycorrhizae. Soil Sci. Soc. Am. J. 1977, 41, 551–556. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. J. Math. Methods Biosci. 2008, 50, 346–363. [Google Scholar] [CrossRef] [Green Version]
- Harrell, F.E., Jr.; Harrell, M.F.E., Jr. Package ‘hmisc’. CRAN2018 2019. 2019, pp. 235–236. Available online: https://cran.uib.no/web/packages/Hmisc/Hmisc.pdf (accessed on 20 October 2021).
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Bioinformatics 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Tingey, D.T.; Phillips, D.L.; Johnson, M.G. Elevated CO2 and conifer roots: Effects on growth, life span and turnover. New Phytol. 2000, 147, 87–103. [Google Scholar] [CrossRef]
- Burton, A.; Pregitzer, K.; Hendrick, R. Relationships between fine root dynamics and nitrogen availability in Michigan northern hardwood forests. Oecologia 2000, 125, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Lukac, M.; Calfapietra, C.; Godbold, D.L. Production, turnover and mycorrhizal colonization of root systems of three Populus species grown under elevated CO2 (POPFACE). Glob. Chang. Biol. 2003, 9, 838–848. [Google Scholar] [CrossRef]
- De Graaff, M.-A.; Six, J.; Jastrow, J.D.; Schadt, C.W.; Wullschleger, S.D. Variation in root architecture among switchgrass cultivars impacts root decomposition rates. Soil Biol. Biochem. 2013, 58, 198–206. [Google Scholar] [CrossRef]
- Rabot, E.; Wiesmeier, M.; Schlüter, S.; Vogel, H.-J. Soil structure as an indicator of soil functions: A review. Geoderma 2018, 314, 122–137. [Google Scholar] [CrossRef]
- Kumar, R.; Rawat, K.S.; Singh, J.; Singh, A.; Rai, A. Soil aggregation dynamics and carbon sequestration. J. Appl. Nat. Sci. 2013, 5, 250–267. [Google Scholar] [CrossRef]
- Sanz-Saez, A.; Lopez, U.P.; Del-Canto, A.; Ortiz-Barredo, A.; Petite, A.M.; Aranjuelo, I.; Muñoz-Rueda, A.; Lacuesta, M. Changes in environmental CO2 concentration can modify Rhizobium-soybean specificity and condition plant fitness and productivity. Environ. Exp. Bot. 2019, 162, 133–143. [Google Scholar] [CrossRef]
- Wilson, G.W.T.; Rice, C.W.; Rillig, M.C.; Springer, A.; Hartnett, D.C. Soil aggregation and carbon sequestration are tightly correlated with the abundance of arbuscular mycorrhizal fungi: Results from long-term field experiments. Ecol. Lett. 2009, 12, 452–461. [Google Scholar] [CrossRef]
- Medlyn, B.E.; Barton, C.; Broadmeadow, M.; Ceulemans, R.; De Angelis, P.; Forstreuter, M.; Freeman, M.; Jackson, S.; Kellomäki, S.; Laitat, E. Stomatal conductance of forest species after long-term exposure to elevated CO2 concentration: A synthesis. New Phytol. 2001, 149, 247–264. [Google Scholar] [CrossRef]
- Woodward, F.I. Stomatal numbers are sensitive to increases in CO2 from pre-industrial levels. Nature 1987, 327, 617–618. [Google Scholar] [CrossRef]
- Dubeaux, G.; Hsu, P.-K.; O Ceciliato, P.H.; Swink, K.J.; Rappel, W.-J.; Schroeder, J. Deep dive into CO2-dependent molecular mechanisms driving stomatal responses in plants. Plant Physiol. 2021, kiab342. [Google Scholar] [CrossRef]
- Zhang, J.; Jiao, X.; Du, Q.; Song, X.; Ding, J.; Li, J. Effects of Vapor Pressure Deficit and Potassium Supply on Root Morphology, Potassium Uptake, and Biomass Allocation of Tomato Seedlings. J. Plant Growth Regul. 2021, 40, 509–518. [Google Scholar] [CrossRef]
- Li, H.; Si, B.; Ma, X.; Wu, P. Deep soil water extraction by apple sequesters organic carbon via root biomass rather than altering soil organic carbon content. Sci. Total. Environ. 2019, 670, 662–671. [Google Scholar] [CrossRef] [PubMed]
- Fleishman, S.M.; Bock, H.W.; Eissenstat, D.M.; Centinari, M. Undervine groundcover substantially increases shallow but not deep soil carbon in a temperate vineyard. Agric. Ecosyst. Environ. 2021, 313, 107362. [Google Scholar] [CrossRef]
- Nie, M.; Lu, M.; Bell, J.; Raut, S.; Pendall, E. Altered root traits due to elevated CO2: A meta-analysis. Glob. Ecol. Biogeogr. 2013, 22, 1095–1105. [Google Scholar] [CrossRef]
- Han, Y.; Zhang, X.; Ma, X. Fine Root Length of Maize Decreases in Response to Elevated CO2 Levels in Soil. Appl. Sci. 2020, 10, 968. [Google Scholar] [CrossRef] [Green Version]
- Lake, J.A.; Steven, M.D.; Smith, K.L.; Lomax, B.H. Plant responses to elevated CO2 levels in soils: Distinct CO2 and O2 -depletion effects. Int. J. Greenh. Gas Control. 2017, 64, 333–339. [Google Scholar] [CrossRef]
- Staddon, P.; Fitter, A.H.; Graves, J.D. Effect of elevated atmospheric CO2 on mycorrhizal colonization, external mycorrhizal hyphal production and phosphorus inflow in Plantago lanceolata and Trifolium repens in association with the arbuscular mycorrhizal fungus Glomus mosseae. Glob. Chang. Biol. 1999, 5, 347–358. [Google Scholar] [CrossRef]
- Hodge, A. The plastic plant: Root responses to heterogeneous supplies of nutrients. New Phytol. 2004, 162, 9–24. [Google Scholar] [CrossRef]
- Morgan, J.; Knight, W.; Dudley, L.; Hunt, H. Enhanced root system C-sink activity, water relations and aspects of nutrient acquisition in mycotrophic Bouteloua gracilis subjected to CO2 enrichment. In Belowground Responses to Rising Atmospheric CO2: Implications for Plants, Soil Biota, and Ecosystem Processes; Springer: Berlin/Heidelberg, Germany, 1994. [Google Scholar]
- Pellissier, L.; Litsios, G.; Fishbein, M.; Salamin, N.; Agrawal, A.A.; Rasmann, S. Different rates of defense evolution and niche preferences in clonal and nonclonal milkweeds (Asclepias spp.). New Phytol. 2016, 209, 1230–1239. [Google Scholar] [CrossRef] [PubMed]
- Fishbein, M.; Chuba, D.; Ellison, C.; Mason-Gamer, R.J.; Lynch, S.P. Phylogenetic Relationships of Asclepias (Apocynaceae) Inferred from Non-coding Chloroplast DNA Sequences. Syst. Bot. 2011, 36, 1008–1023. [Google Scholar] [CrossRef] [Green Version]
- Shtark, O.; Puzanskiy, R.; Avdeeva, G.; Yemelyanov, V.; Shavarda, A.; Romanyuk, D.; Kliukova, M.; Kirpichnikova, A.; Tikhonovich, I.; Zhukov, V. Metabolic Alterations in Pisum sativum Roots during Plant Growth and Arbuscular Mycorrhiza Development. Plants 2021, 10, 1033. [Google Scholar] [CrossRef] [PubMed]
- Laparre, J.; Malbreil, M.; Letisse, F.; Portais, J.C.; Roux, C.; Bécard, G.; Puech-pagès, V. Combining metabolomics and gene expression analysis reveals that propionyl-and butyryl-carnitines are involved in late stages of arbuscular mycorrhizal symbiosis. Mol. Plant. 2014, 7, 554–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mustafa, A.; Minggang, X.; Shah, S.A.A.; Abrar, M.M.; Nan, S.; Baoren, W.; Zejiang, C.; Saeed, Q.; Naveed, M.; Mehmood, K. Soil aggregation and soil aggregate stability regulate organic carbon and nitrogen storage in a red soil of southern China. J. Environ. Manag. 2020, 270, 110894. [Google Scholar] [CrossRef] [PubMed]
- Bodner, G.; Mentler, A.; Keiblinger, K. Plant Roots for Sustainable Soil Structure Management in Cropping Systems. Root Syst. Sustain. Agric. Intensif. 2021, 45–90. [Google Scholar]
- Lu, M.; Yang, M.; Yang, Y.; Wang, D.; Sheng, L. Soil carbon and nutrient sequestration linking to soil aggregate in a temperate fen in Northeast China. Ecol. Indic. 2019, 98, 869–887. [Google Scholar] [CrossRef]
Figure 1.
Imagery shows a subset of plants from the experiment. The top panel represents roots with mycorrhizae, the bottom panel represent roots void of mycorrhizae. Mycorrhizae had pronounced effects on root systems. In tap-root systems, the dominant root was more likely to be modified into a tuber-like structure.
Figure 1.
Imagery shows a subset of plants from the experiment. The top panel represents roots with mycorrhizae, the bottom panel represent roots void of mycorrhizae. Mycorrhizae had pronounced effects on root systems. In tap-root systems, the dominant root was more likely to be modified into a tuber-like structure.

Figure 2.
The outcome of mycorrhizae and CO2 (predictors) on explanatory variables (i.e., relative transportive root and biomass). Relative transportive root response is the percent difference in transportive root versus absorptive root diameter. Biomass response is dry weight in grams. The main effect of predictors on biomass are shown in (a,b). The main effect of the predictors on root response are shown in (c,d). Asclepias spp. biomass response to CO2 and mycorrhizae (i.e., predictors) are shown in (e–j)). Asclepias spp. root response to CO2 and mycorrhizae (i.e., predictors) are shown in (k–p). Error bars are standard error in (a–d). Significant codes: ‘***’ p < 0; ‘**’ p < 0.001.
Figure 2.
The outcome of mycorrhizae and CO2 (predictors) on explanatory variables (i.e., relative transportive root and biomass). Relative transportive root response is the percent difference in transportive root versus absorptive root diameter. Biomass response is dry weight in grams. The main effect of predictors on biomass are shown in (a,b). The main effect of the predictors on root response are shown in (c,d). Asclepias spp. biomass response to CO2 and mycorrhizae (i.e., predictors) are shown in (e–j)). Asclepias spp. root response to CO2 and mycorrhizae (i.e., predictors) are shown in (k–p). Error bars are standard error in (a–d). Significant codes: ‘***’ p < 0; ‘**’ p < 0.001.

Figure 3.
The relationship with biomass and relative root diameter. Each point represents an individual plant from the experiment.
Figure 3.
The relationship with biomass and relative root diameter. Each point represents an individual plant from the experiment.

Figure 4.
The relationship with biomass and relative transportive root across experimental factors. This relationship is decoupled by factor (a) mycorrhizae; (b) CO2 regime; (c) species; (d) root systems. Ellipses represent 95% CI for the levels in each of the factors described.
Figure 4.
The relationship with biomass and relative transportive root across experimental factors. This relationship is decoupled by factor (a) mycorrhizae; (b) CO2 regime; (c) species; (d) root systems. Ellipses represent 95% CI for the levels in each of the factors described.

Figure 5.
Provided here is a maximum likelihood tree for 6 the Asclepias species. This tree was constructed using highest log likelihood (−1398.00) for the rbcL amino acid sequence (rbcL is a gene involved in CO2 fixation). According to this tree, fibrous root species, A. verticillata and A. incarnata, are distant from the 4 tap-root species.
Figure 5.
Provided here is a maximum likelihood tree for 6 the Asclepias species. This tree was constructed using highest log likelihood (−1398.00) for the rbcL amino acid sequence (rbcL is a gene involved in CO2 fixation). According to this tree, fibrous root species, A. verticillata and A. incarnata, are distant from the 4 tap-root species.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Root system, milkweed, and qualitative observations of the 6 Asclepias species.
| Root System | Species | Common Name | Niche | Experimental Observation |
|---|---|---|---|---|
| Fibrous |
| Swamp milkweed | Prefers moist wet soils (wildflower.org), and is found in standing water several months of the year [38]. In addition, many stems arise from a single root stock [38]. | Fine roots became increasingly coarse when colonized with mycorrhizae. |
| Whorled milkweed | Has divergent traits including shallow fine roots that are 5–10 cm deep [39]. Additionally, prefers xeric soils (Missouri botanical garden.org), and grows extensive rhizome systems [38]. | Very dense fine roots that tightly aggregated the soil together when inoculated with mycorrhizae. | |
| Tap |
| Common milkweed | Prefers well-drained loamy soils (wildflower.org). Grows large extensive rhizome systems while adjacent stems may belong to the same or different clones [38]. | Primary/dominant root is modified into tuber-like structures when colonized with mycorrhizae. |
| Prairie milkweed | Prefers wet moist soils(wildflower.org). Stems arise from fleshy rhizomes, as vegetative reproduction is common [38]. | Primary/dominant root is modified into tuber-like structures when colonized with mycorrhizae. | |
| Butterfly milkweed | Thick woody orange-brown tap-root that serves for C storage, attachment, and perennation. This species produces smaller lateral roots [40]. Many stems arise from a single root stock [38] | Primary/dominant root is modified into tuber-like structures when colonized with mycorrhizae. | |
| Spider milkweed | Prefers moist soils (wildflower.org). Root cardenolides in this species are four times higher than those of common milkweed [41]. | Primary/dominant root is modified into tuber-like structures, but, in some cases, the tuber-like root was less elongated and more gall-like when colonized with mycorrhizae. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Malik, R.J.; Bever, J.D. Enriched CO2 and Root-Associated Fungi (Mycorrhizae) Yield Inverse Effects on Plant Mass and Root Morphology in Six Asclepias Species. Plants 2021, 10, 2474. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10112474
AMA Style
Malik RJ, Bever JD. Enriched CO2 and Root-Associated Fungi (Mycorrhizae) Yield Inverse Effects on Plant Mass and Root Morphology in Six Asclepias Species. Plants. 2021; 10(11):2474. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10112474
Chicago/Turabian StyleMalik, Rondy J., and James D. Bever. 2021. "Enriched CO2 and Root-Associated Fungi (Mycorrhizae) Yield Inverse Effects on Plant Mass and Root Morphology in Six Asclepias Species" Plants 10, no. 11: 2474. https://0-doi-org.brum.beds.ac.uk/10.3390/plants10112474
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.