
The Therapeutic Potential of Secreted Factors from Dental Pulp Stem Cells for Various Diseases

 , ,
, ,
Abstract
:1. Background
2. Cytokines Contained in DPSC-CM
3. Cerebrovascular Disease
4. Heart Disease
5. Lung Disease
6. Liver Disease
7. Eye Disease
8. Neurological Disorder
9. Autoimmune Disease
10. Bone and Cartilage Disease
11. Metabolic Disease
12. Dental Diseases
13. Relationship between DPSC-CM and Exosomes
14. Current Limitation and Perspective
15. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamura, T. Differentiation of pulpal cells and inductive influences of various matrices with reference to pulpal wound healing. J. Dent. Res. 1985, 64, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Mattei, V.; Martellucci, S.; Pulcini, F.; Santilli, F.; Sorice, M.; Delle Monache, S. Regenerative Potential of DPSCs and Revascularization: Direct, Paracrine or Autocrine Effect? Stem Cell Rev. Rep. 2021, 17, 1635–1646. [Google Scholar] [CrossRef] [PubMed]
- Gang, E.J.; Bosnakovski, D.; Figueiredo, C.A.; Visser, J.W.; Perlingeiro, R.C. SSEA-4 identifies mesenchymal stem cells from bone marrow. Blood 2007, 109, 1743–1751. [Google Scholar] [CrossRef] [PubMed]
- Martinez, C.; Hofmann, T.J.; Marino, R.; Dominici, M.; Horwitz, E.M. Human bone marrow mesenchymal stromal cells express the neural ganglioside GD2: A novel surface marker for the identification of MSCs. Blood 2007, 109, 4245–4248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakopoulou, A.; Apatzidou, D.; Aggelidou, E.; Gousopoulou, E.; Leyhausen, G.; Volk, J.; Kritis, A.; Koidis, P.; Geurtsen, W. Isolation and prolonged expansion of oral mesenchymal stem cells under clinical-grade, GMP-compliant conditions differentially affects “stemness” properties. Stem Cell Res. Ther. 2017, 8, 247. [Google Scholar] [CrossRef]
- Qiu, Z.; Lin, S.; Hu, X.; Zeng, J.; Xiao, T.; Ke, Z.; Lv, H. Involvement of miR-146a-5p/neurogenic locus notch homolog protein 1 in the proliferation and differentiation of STRO-1(+) human dental pulp stem cells. Eur. J. Oral. Sci. 2019, 127, 294–303. [Google Scholar] [CrossRef]
- Petrenko, Y.; Vackova, I.; Kekulova, K.; Chudickova, M.; Koci, Z.; Turnovcova, K.; Kupcova Skalnikova, H.; Vodicka, P.; Kubinova, S. A Comparative Analysis of Multipotent Mesenchymal Stromal Cells derived from Different Sources, with a Focus on Neuroregenerative Potential. Sci. Rep. 2020, 10, 4290. [Google Scholar] [CrossRef]
- Ouchi, T.; Nakagawa, T. Mesenchymal stem cell-based tissue regeneration therapies for periodontitis. Regen. Ther. 2020, 14, 72–78. [Google Scholar] [CrossRef]
- Jo, H.; Brito, S.; Kwak, B.M.; Park, S.; Lee, M.G.; Bin, B.H. Applications of Mesenchymal Stem Cells in Skin Regeneration and Rejuvenation. Int. J. Mol. Sci. 2021, 22, 2410. [Google Scholar] [CrossRef]
- Ulpiano, C.; da Silva, C.L.; Monteiro, G.A. Mesenchymal Stromal Cells (MSCs): A Promising Tool for Cell-Based Angiogenic Therapy. Curr. Gene 2021, 21, 382–405. [Google Scholar] [CrossRef] [PubMed]
- Das, M.; Mayilsamy, K.; Mohapatra, S.S.; Mohapatra, S. Mesenchymal stem cell therapy for the treatment of traumatic brain injury: Progress and prospects. Rev. Neurosci. 2019, 30, 839–855. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Ji, C.; Lu, L. Mesenchymal stem cell therapy for liver fibrosis/cirrhosis. Ann. Transl. Med. 2020, 8, 562. [Google Scholar] [CrossRef] [PubMed]
- Sandona, M.; Di Pietro, L.; Esposito, F.; Ventura, A.; Silini, A.R.; Parolini, O.; Saccone, V. Mesenchymal Stromal Cells and Their Secretome: New Therapeutic Perspectives for Skeletal Muscle Regeneration. Front. Bioeng. Biotechnol. 2021, 9, 652970. [Google Scholar] [CrossRef]
- Van Nguyen, T.T.; Vu, N.B.; Van Pham, P. Mesenchymal Stem Cell Transplantation for Ischemic Diseases: Mechanisms and Challenges. Tissue Eng. Regen. Med. 2021, 18, 587–611. [Google Scholar] [CrossRef]
- Gotherstrom, C.; Walther-Jallow, L. Stem Cell Therapy as a Treatment for Osteogenesis Imperfecta. Curr. Osteoporos. Rep. 2020, 18, 337–343. [Google Scholar] [CrossRef]
- Young, F.; Sloan, A.; Song, B. Dental pulp stem cells and their potential roles in central nervous system regeneration and repair. J. Neurosci. Res. 2013, 91, 1383–1393. [Google Scholar] [CrossRef]
- Ahmed Nel, M.; Murakami, M.; Hirose, Y.; Nakashima, M. Therapeutic Potential of Dental Pulp Stem Cell Secretome for Alzheimer’s Disease Treatment: An In Vitro Study. Stem Cells Int. 2016, 2016, 8102478. [Google Scholar] [CrossRef]
- Kim, H.J.; Cho, Y.A.; Lee, Y.M.; Lee, S.Y.; Bae, W.J.; Kim, E.C. PIN1 Suppresses the Hepatic Differentiation of Pulp Stem Cells via Wnt3a. J. Dent. Res. 2016, 95, 1415–1424. [Google Scholar] [CrossRef]
- Mead, B.; Logan, A.; Berry, M.; Leadbeater, W.; Scheven, B.A. Concise Review: Dental Pulp Stem Cells: A Novel Cell Therapy for Retinal and Central Nervous System Repair. Stem Cells 2017, 35, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Nicola, F.D.C.; Marques, M.R.; Odorcyk, F.; Arcego, D.M.; Petenuzzo, L.; Aristimunha, D.; Vizuete, A.; Sanches, E.F.; Pereira, D.P.; Maurmann, N.; et al. Neuroprotector effect of stem cells from human exfoliated deciduous teeth transplanted after traumatic spinal cord injury involves inhibition of early neuronal apoptosis. Brain Res. 2017, 1663, 95–105. [Google Scholar] [CrossRef]
- Ide, C.; Nakai, Y.; Nakano, N.; Seo, T.B.; Yamada, Y.; Endo, K.; Noda, T.; Saito, F.; Suzuki, Y.; Fukushima, M.; et al. Bone marrow stromal cell transplantation for treatment of sub-acute spinal cord injury in the rat. Brain Res. 2010, 1332, 32–47. [Google Scholar] [CrossRef]
- Osugi, M.; Katagiri, W.; Yoshimi, R.; Inukai, T.; Hibi, H.; Ueda, M. Conditioned media from mesenchymal stem cells enhanced bone regeneration in rat calvarial bone defects. Tissue Eng. Part A 2012, 18, 1479–1489. [Google Scholar] [CrossRef] [Green Version]
- Inukai, T.; Katagiri, W.; Yoshimi, R.; Osugi, M.; Kawai, T.; Hibi, H.; Ueda, M. Novel application of stem cell-derived factors for periodontal regeneration. Biochem. Biophys. Res. Commun. 2013, 430, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Katagiri, W.; Osugi, M.; Sugimura, Y.; Hibi, H.; Ueda, M. Secretomes from bone marrow-derived mesenchymal stromal cells enhance periodontal tissue regeneration. Cytotherapy 2015, 17, 369–381. [Google Scholar] [CrossRef]
- Ogata, K.; Katagiri, W.; Osugi, M.; Kawai, T.; Sugimura, Y.; Hibi, H.; Nakamura, S.; Ueda, M. Evaluation of the therapeutic effects of conditioned media from mesenchymal stem cells in a rat bisphosphonate-related osteonecrosis of the jaw-like model. Bone 2015, 74, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Sugimura-Wakayama, Y.; Katagiri, W.; Osugi, M.; Kawai, T.; Ogata, K.; Sakaguchi, K.; Hibi, H. Peripheral Nerve Regeneration by Secretomes of Stem Cells from Human Exfoliated Deciduous Teeth. Stem Cells Dev. 2015. [Google Scholar] [CrossRef]
- Yamada, Y.; Nakamura-Yamada, S.; Umemura-Kubota, E.; Baba, S. Diagnostic Cytokines and Comparative Analysis Secreted from Exfoliated Deciduous Teeth, Dental Pulp, and Bone Marrow Derived Mesenchymal Stem Cells for Functional Cell-Based Therapy. Int. J. Mol. Sci. 2019, 20, 5900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bousnaki, M.; Bakopoulou, A.; Pich, A.; Papachristou, E.; Kritis, A.; Koidis, P. Mapping the Secretome of Dental Pulp Stem Cells Under Variable Microenvironmental Conditions. Stem Cell Rev. Rep. 2021, 18, 1372–1407. [Google Scholar] [CrossRef] [PubMed]
- Matsumura-Kawashima, M.; Ogata, K.; Moriyama, M.; Murakami, Y.; Kawado, T.; Nakamura, S. Secreted factors from dental pulp stem cells improve Sjogren’s syndrome via regulatory T cell-mediated immunosuppression. Stem Cell Res. Ther. 2021, 12, 182. [Google Scholar] [CrossRef]
- Ogata, K.; Matsumura-Kawashima, M.; Moriyama, M.; Kawado, T.; Nakamura, S. Dental pulp-derived stem cell-conditioned media attenuates secondary Sjogren’s syndrome via suppression of inflammatory cytokines in the submandibular glands. Regen. Ther. 2021, 16, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T.; Sugiyama, M.; Hattori, H.; Wakita, H.; Wakabayashi, T.; Ueda, M. Stem cells from human exfoliated deciduous tooth-derived conditioned medium enhance recovery of focal cerebral ischemia in rats. Tissue Eng. Part A 2013, 19, 24–29. [Google Scholar] [CrossRef] [Green Version]
- Yamagata, M.; Yamamoto, A.; Kako, E.; Kaneko, N.; Matsubara, K.; Sakai, K.; Sawamoto, K.; Ueda, M. Human dental pulp-derived stem cells protect against hypoxic-ischemic brain injury in neonatal mice. Stroke 2013, 44, 551–554. [Google Scholar] [CrossRef]
- Yamamoto, A.; Sakai, K.; Matsubara, K.; Kano, F.; Ueda, M. Multifaceted neuro-regenerative activities of human dental pulp stem cells for functional recovery after spinal cord injury. Neurosci. Res. 2014, 78, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.F.; Chen, K.W.; Chien, Y.; Lai, Y.H.; Hsieh, S.T.; Ma, H.Y.; Wang, K.C.; Shiau, C.Y. Dental Pulp Stem Cell-Derived Factors Alleviate Subarachnoid Hemorrhage-Induced Neuroinflammation and Ischemic Neurological Deficits. Int. J. Mol. Sci. 2019, 20, 3747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakai, K.; Yamamoto, A.; Matsubara, K.; Nakamura, S.; Naruse, M.; Yamagata, M.; Sakamoto, K.; Tauchi, R.; Wakao, N.; Imagama, S.; et al. Human dental pulp-derived stem cells promote locomotor recovery after complete transection of the rat spinal cord by multiple neuro-regenerative mechanisms. J. Clin. Investig. 2012, 122, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Aliaghaei, A.; Boroujeni, M.E.; Ahmadi, H.; Bayat, A.H.; Tavirani, M.R.; Abdollahifar, M.A.; Pooyafar, M.H.; Mansouri, V. Dental pulp stem cell transplantation ameliorates motor function and prevents cerebellar atrophy in rat model of cerebellar ataxia. Cell Tissue Res. 2019, 376, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Willis, C.M.; Nicaise, A.M.; Peruzzotti-Jametti, L.; Pluchino, S. The neural stem cell secretome and its role in brain repair. Brain Res. 2020, 1729, 146615. [Google Scholar] [CrossRef]
- Luzuriaga, J.; Polo, Y.; Pastor-Alonso, O.; Pardo-Rodriguez, B.; Larranaga, A.; Unda, F.; Sarasua, J.R.; Pineda, J.R.; Ibarretxe, G. Advances and Perspectives in Dental Pulp Stem Cell Based Neuroregeneration Therapies. Int. J. Mol. Sci. 2021, 22, 3546. [Google Scholar] [CrossRef]
- Iadecola, C.; Buckwalter, M.S.; Anrather, J. Immune responses to stroke: Mechanisms, modulation, and therapeutic potential. J. Clin. Investig. 2020, 130, 2777–2788. [Google Scholar] [CrossRef]
- Kawabori, M.; Shichinohe, H.; Kuroda, S.; Houkin, K. Clinical Trials of Stem Cell Therapy for Cerebral Ischemic Stroke. Int. J. Mol. Sci. 2020, 21, 7380. [Google Scholar] [CrossRef]
- Nie, G.; Wen, X.; Liang, X.; Zhao, H.; Li, Y.; Lu, J. Additional evidence supports association of common genetic variants in MMP3 and TIMP2 with increased risk of chronic Achilles tendinopathy susceptibility. J. Sci. Med. Sport 2019, 22, 1074–1078. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.H.; Cai, Q.; Hu, Z.; Patel, M.; Bard, J.; Jamison, J.; Coling, D. Metalloproteinases and their associated genes contribute to the functional integrity and noise-induced damage in the cochlear sensory epithelium. J. Neurosci. Off. J. Soc. Neurosci. 2012, 32, 14927–14941. [Google Scholar] [CrossRef] [PubMed]
- Markwerth, P.; Bajanowski, T.; Tzimas, I.; Dettmeyer, R. Sudden cardiac death-update. Int. J. Leg. Med. 2021, 135, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Heusch, G. Myocardial ischaemia-reperfusion injury and cardioprotection in perspective. Nat. Rev. Cardiol. 2020, 17, 773–789. [Google Scholar] [CrossRef]
- Yamaguchi, S.; Shibata, R.; Yamamoto, N.; Nishikawa, M.; Hibi, H.; Tanigawa, T.; Ueda, M.; Murohara, T.; Yamamoto, A. Dental pulp-derived stem cell conditioned medium reduces cardiac injury following ischemia-reperfusion. Sci. Rep. 2015, 5, 16295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sowa, K.; Nito, C.; Nakajima, M.; Suda, S.; Nishiyama, Y.; Sakamoto, Y.; Nitahara-Kasahara, Y.; Nakamura-Takahashi, A.; Ueda, M.; Kimura, K.; et al. Impact of Dental Pulp Stem Cells Overexpressing Hepatocyte Growth Factor after Cerebral Ischemia/Reperfusion in Rats. Mol. Methods Clin. Dev. 2018, 10, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Cheng, P.; Li, S.; Chen, H. Macrophages in Lung Injury, Repair, and Fibrosis. Cells 2021, 10, 436. [Google Scholar] [CrossRef]
- He, Y.Q.; Zhou, C.C.; Yu, L.Y.; Wang, L.; Deng, J.L.; Tao, Y.L.; Zhang, F.; Chen, W.S. Natural product derived phytochemicals in managing acute lung injury by multiple mechanisms. Pharm. Res. 2021, 163, 105224. [Google Scholar] [CrossRef]
- Murray, P.J.; Wynn, T.A. Protective and pathogenic functions of macrophage subsets. Nat. Rev. Immunol. 2011, 11, 723–737. [Google Scholar] [CrossRef]
- Feng, Y.; Xu, Q.; Yang, Y.; Shi, W.; Meng, W.; Zhang, H.; He, X.; Sun, M.; Chen, Y.; Zhao, J.; et al. The therapeutic effects of bone marrow-derived mesenchymal stromal cells in the acute lung injury induced by sulfur mustard. Stem Cell Res. Ther. 2019, 10, 90. [Google Scholar] [CrossRef] [PubMed]
- Ross, E.A.; Devitt, A.; Johnson, J.R. Macrophages: The Good, the Bad, and the Gluttony. Front. Immunol. 2021, 12, 708186. [Google Scholar] [CrossRef]
- Wakayama, H.; Hashimoto, N.; Matsushita, Y.; Matsubara, K.; Yamamoto, N.; Hasegawa, Y.; Ueda, M.; Yamamoto, A. Factors secreted from dental pulp stem cells show multifaceted benefits for treating acute lung injury in mice. Cytotherapy 2015, 17, 1119–1129. [Google Scholar] [CrossRef] [PubMed]
- Ionescu, L.; Byrne, R.N.; van Haaften, T.; Vadivel, A.; Alphonse, R.S.; Rey-Parra, G.J.; Weissmann, G.; Hall, A.; Eaton, F.; Thebaud, B. Stem cell conditioned medium improves acute lung injury in mice: In vivo evidence for stem cell paracrine action. Am. J. Physiol. Lung. Cell Mol. Physiol. 2012, 303, L967–L977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stravitz, R.T.; Lee, W.M. Acute liver failure. Lancet 2019, 394, 869–881. [Google Scholar] [CrossRef]
- Miyajima, A.; Tanaka, M.; Itoh, T. Stem/progenitor cells in liver development, homeostasis, regeneration, and reprogramming. Cell Stem Cell 2014, 14, 561–574. [Google Scholar] [CrossRef] [Green Version]
- Michalopoulos, G.K.; Bhushan, B. Liver regeneration: Biological and pathological mechanisms and implications. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 40–55. [Google Scholar] [CrossRef]
- Kisseleva, T.; Brenner, D. Molecular and cellular mechanisms of liver fibrosis and its regression. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 151–166. [Google Scholar] [CrossRef]
- Hu, C.; Zhao, L.; Zhang, L.; Bao, Q.; Li, L. Mesenchymal stem cell-based cell-free strategies: Safe and effective treatments for liver injury. Stem Cell Res. Ther. 2020, 11, 377. [Google Scholar] [CrossRef]
- Chinnici, C.M.; Russelli, G.; Bulati, M.; Miceli, V.; Gallo, A.; Busa, R.; Tinnirello, R.; Conaldi, P.G.; Iannolo, G. Mesenchymal stromal cell secretome in liver failure: Perspectives on COVID-19 infection treatment. World J. Gastroenterol. 2021, 27, 1905–1919. [Google Scholar] [CrossRef]
- Hirata, M.; Ishigami, M.; Matsushita, Y.; Ito, T.; Hattori, H.; Hibi, H.; Goto, H.; Ueda, M.; Yamamoto, A. Multifaceted Therapeutic Benefits of Factors Derived from Dental Pulp Stem Cells for Mouse Liver Fibrosis. Stem Cells Transl. Med. 2016, 5, 1416–1424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsushita, Y.; Ishigami, M.; Matsubara, K.; Kondo, M.; Wakayama, H.; Goto, H.; Ueda, M.; Yamamoto, A. Multifaceted therapeutic benefits of factors derived from stem cells from human exfoliated deciduous teeth for acute liver failure in rats. J. Tissue Eng. Regen. Med. 2017, 11, 1888–1896. [Google Scholar] [CrossRef] [PubMed]
- Okano, J.; Shiota, G.; Matsumoto, K.; Yasui, S.; Kurimasa, A.; Hisatome, I.; Steinberg, P.; Murawaki, Y. Hepatocyte growth factor exerts a proliferative effect on oval cells through the PI3K/AKT signaling pathway. Biochem. Biophys. Res. Commun. 2003, 309, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Moumen, A.; Ieraci, A.; Patane, S.; Sole, C.; Comella, J.X.; Dono, R.; Maina, F. Met signals hepatocyte survival by preventing Fas-triggered FLIP degradation in a PI3k-Akt-dependent manner. Hepatology 2007, 45, 1210–1217. [Google Scholar] [CrossRef]
- Hu, B.; Colletti, L.M. Stem cell factor and c-kit are involved in hepatic recovery after acetaminophen-induced liver injury in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G45–G53. [Google Scholar] [CrossRef]
- Leu, J.I.; Crissey, M.A.; Taub, R. Massive hepatic apoptosis associated with TGF-beta1 activation after Fas ligand treatment of IGF binding protein-1-deficient mice. J. Clin. Investig. 2003, 111, 129–139. [Google Scholar] [CrossRef] [Green Version]
- Duarte, S.; Hamada, T.; Kuriyama, N.; Busuttil, R.W.; Coito, A.J. TIMP-1 deficiency leads to lethal partial hepatic ischemia and reperfusion injury. Hepatology 2012, 56, 1074–1085. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Xu, Z. Mechanisms of action of angiogenin. Acta Biochim. Biophys. Sin. 2008, 40, 619–624. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Tang, S.; Liao, J.; Liu, M.; Lin, Y. VEGF165 gene-modified human umbilical cord blood mesenchymal stem cells protect against acute liver failure in rats. J. Gene Med. 2021, 23, e3369. [Google Scholar] [CrossRef]
- Lin, Y.; Dong, M.Q.; Liu, Z.M.; Xu, M.; Huang, Z.H.; Liu, H.J.; Gao, Y.; Zhou, W.J. A strategy of vascular-targeted therapy for liver fibrosis. Hepatology 2021. [Google Scholar] [CrossRef]
- Roca, H.; Varsos, Z.S.; Sud, S.; Craig, M.J.; Ying, C.; Pienta, K.J. CCL2 and interleukin-6 promote survival of human CD11b+ peripheral blood mononuclear cells and induce M2-type macrophage polarization. J. Biol. Chem. 2009, 284, 34342–34354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newton, F.; Megaw, R. Mechanisms of Photoreceptor Death in Retinitis Pigmentosa. Genes 2020, 11, 1120. [Google Scholar] [CrossRef] [PubMed]
- Sahel, J.A.; Boulanger-Scemama, E.; Pagot, C.; Arleo, A.; Galluppi, F.; Martel, J.N.; Esposti, S.D.; Delaux, A.; de Saint Aubert, J.B.; de Montleau, C.; et al. Partial recovery of visual function in a blind patient after optogenetic therapy. Nat. Med. 2021, 27, 1223–1229. [Google Scholar] [CrossRef] [PubMed]
- Li, X.X.; Yuan, X.J.; Zhai, Y.; Yu, S.; Jia, R.X.; Yang, L.P.; Ma, Z.Z.; Zhao, Y.M.; Wang, Y.X.; Ge, L.H. Treatment with Stem Cells from Human Exfoliated Deciduous Teeth and Their Derived Conditioned Medium Improves Retinal Visual Function and Delays the Degeneration of Photoreceptors. Stem Cells Dev. 2019, 28, 1514–1526. [Google Scholar] [CrossRef]
- Bray, A.F.; Cevallos, R.R.; Gazarian, K.; Lamas, M. Human dental pulp stem cells respond to cues from the rat retina and differentiate to express the retinal neuronal marker rhodopsin. Neuroscience 2014, 280, 142–155. [Google Scholar] [CrossRef] [PubMed]
- Zrzavy, T.; Schwaiger, C.; Wimmer, I.; Berger, T.; Bauer, J.; Butovsky, O.; Schwab, J.M.; Lassmann, H.; Hoftberger, R. Acute and non-resolving inflammation associate with oxidative injury after human spinal cord injury. Brain 2021, 144, 144–161. [Google Scholar] [CrossRef]
- Devanney, N.A.; Stewart, A.N.; Gensel, J.C. Microglia and macrophage metabolism in CNS injury and disease: The role of immunometabolism in neurodegeneration and neurotrauma. Exp. Neurol. 2020, 329, 113310. [Google Scholar] [CrossRef]
- Tan, G.; Qian, D.; Li, L.; Wang, X.; Zhou, Z.; Liu, W.; Zhu, T.; Ning, B.; Qian, Y.; Zhou, W.; et al. Parthenolide promotes the repair of spinal cord injury by modulating M1/M2 polarization via the NF-kappaB and STAT 1/3 signaling pathway. Cell Death Discov. 2020, 6, 97. [Google Scholar] [CrossRef]
- Kobashi, S.; Terashima, T.; Katagi, M.; Nakae, Y.; Okano, J.; Suzuki, Y.; Urushitani, M.; Kojima, H. Transplantation of M2-Deviated Microglia Promotes Recovery of Motor Function after Spinal Cord Injury in Mice. Mol. Ther. 2020, 28, 254–265. [Google Scholar] [CrossRef]
- Zheng, Q.; Zhang, J.; Zuo, X.; Sun, J.; Liang, Z.; Hu, X.; Wang, Z.; Li, K.; Song, J.; Ding, T.; et al. Photobiomodulation Promotes Neuronal Axon Regeneration After Oxidative Stress and Induces a Change in Polarization from M1 to M2 in Macrophages via Stimulation of CCL2 in Neurons: Relevance to Spinal Cord Injury. J. Mol. Neurosci. 2021, 71, 1290–1300. [Google Scholar] [CrossRef]
- Zhang, J.; Lu, X.; Feng, G.; Gu, Z.; Sun, Y.; Bao, G.; Xu, G.; Lu, Y.; Chen, J.; Xu, L.; et al. Chitosan scaffolds induce human dental pulp stem cells to neural differentiation: Potential roles for spinal cord injury therapy. Cell Tissue Res. 2016, 366, 129–142. [Google Scholar] [CrossRef] [PubMed]
- Asadi-Golshan, R.; Razban, V.; Mirzaei, E.; Rahmanian, A.; Khajeh, S.; Mostafavi-Pour, Z.; Dehghani, F. Sensory and Motor Behavior Evidences Supporting the Usefulness of Conditioned Medium from Dental Pulp-Derived Stem Cells in Spinal Cord Injury in Rats. Asian Spine J. 2018, 12, 785–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsubara, K.; Matsushita, Y.; Sakai, K.; Kano, F.; Kondo, M.; Noda, M.; Hashimoto, N.; Imagama, S.; Ishiguro, N.; Suzumura, A.; et al. Secreted ectodomain of sialic acid-binding Ig-like lectin-9 and monocyte chemoattractant protein-1 promote recovery after rat spinal cord injury by altering macrophage polarity. J. Neurosci. Off. J. Soc. Neurosci. 2015, 35, 2452–2464. [Google Scholar] [CrossRef] [Green Version]
- Kano, F.; Matsubara, K.; Ueda, M.; Hibi, H.; Yamamoto, A. Secreted Ectodomain of Sialic Acid-Binding Ig-Like Lectin-9 and Monocyte Chemoattractant Protein-1 Synergistically Regenerate Transected Rat Peripheral Nerves by Altering Macrophage Polarity. Stem Cells 2017, 35, 641–653. [Google Scholar] [CrossRef] [PubMed]
- Pajares, M.; Rojo, A.I.; Manda, G.; Bosca, L.; Cuadrado, A. Inflammation in Parkinson’s Disease: Mechanisms and Therapeutic Implications. Cells 2020, 9, 1687. [Google Scholar] [CrossRef]
- Bloem, B.R.; Okun, M.S.; Klein, C. Parkinson’s disease. Lancet 2021, 397, 2284–2303. [Google Scholar] [CrossRef]
- Chen, Y.R.; Lai, P.L.; Chien, Y.; Lee, P.H.; Lai, Y.H.; Ma, H.I.; Shiau, C.Y.; Wang, K.C. Improvement of Impaired Motor Functions by Human Dental Exfoliated Deciduous Teeth Stem Cell-Derived Factors in a Rat Model of Parkinson’s Disease. Int. J. Mol. Sci. 2020, 21, 3807. [Google Scholar] [CrossRef]
- Abeysinghe, A.; Deshapriya, R.; Udawatte, C. Alzheimer’s disease; A review of the pathophysiological basis and therapeutic interventions. Life Sci. 2020, 256, 117996. [Google Scholar] [CrossRef]
- Sengoku, R. Aging and Alzheimer’s disease pathology. Neuropathology 2020, 40, 22–29. [Google Scholar] [CrossRef]
- Lei, P.; Ayton, S.; Bush, A.I. The essential elements of Alzheimer’s disease. J. Biol. Chem. 2021, 296, 100105. [Google Scholar] [CrossRef]
- Scheltens, P.; De Strooper, B.; Kivipelto, M.; Holstege, H.; Chetelat, G.; Teunissen, C.E.; Cummings, J.; van der Flier, W.M. Alzheimer’s disease. Lancet 2021, 397, 1577–1590. [Google Scholar] [CrossRef]
- Mita, T.; Furukawa-Hibi, Y.; Takeuchi, H.; Hattori, H.; Yamada, K.; Hibi, H.; Ueda, M.; Yamamoto, A. Conditioned medium from the stem cells of human dental pulp improves cognitive function in a mouse model of Alzheimer’s disease. Behav. Brain Res. 2015, 293, 189–197. [Google Scholar] [CrossRef]
- Norris, S.P.; Likanje, M.N.; Andrews, J.A. Amyotrophic lateral sclerosis: Update on clinical management. Curr. Opin. Neurol. 2020, 33, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zuzzio, K.; Walker, C.L. Systemic Dental Pulp Stem Cell Secretome Therapy in a Mouse Model of Amyotrophic Lateral Sclerosis. Brain Sci. 2019, 9, 165. [Google Scholar] [CrossRef] [Green Version]
- Asadi-Golshan, R.; Razban, V.; Mirzaei, E.; Rahmanian, A.; Khajeh, S.; Mostafavi-Pour, Z.; Dehghani, F. Efficacy of dental pulp-derived stem cells conditioned medium loaded in collagen hydrogel in spinal cord injury in rats: Stereological evidence. J. Chem. Neuroanat. 2021, 116, 101978. [Google Scholar] [CrossRef] [PubMed]
- Shimojima, C.; Takeuchi, H.; Jin, S.; Parajuli, B.; Hattori, H.; Suzumura, A.; Hibi, H.; Ueda, M.; Yamamoto, A. Conditioned Medium from the Stem Cells of Human Exfoliated Deciduous Teeth Ameliorates Experimental Autoimmune Encephalomyelitis. J. Immunol. 2016, 196, 4164–4171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Liu, F.; Zhou, Y.; Jin, B.; Sun, Q.; Guo, S. Immunosuppressive Property of MSCs Mediated by Cell Surface Receptors. Front. Immunol. 2020, 11, 1076. [Google Scholar] [CrossRef]
- Kiselevskii, M.V.; Vlasenko, R.Y.; Stepanyan, N.G.; Shubina, I.Z.; Sitdikova, S.M.; Kirgizov, K.I.; Varfolomeeva, S.R. Secretome of Mesenchymal Bone Marrow Stem Cells: Is It Immunosuppressive or Proinflammatory? Bull. Exp. Biol. Med. 2021, 172, 250–253. [Google Scholar] [CrossRef]
- Krampera, M.; Glennie, S.; Dyson, J.; Scott, D.; Laylor, R.; Simpson, E.; Dazzi, F. Bone marrow mesenchymal stem cells inhibit the response of naive and memory antigen-specific T cells to their cognate peptide. Blood 2003, 101, 3722–3729. [Google Scholar] [CrossRef]
- Sonoyama, W.; Liu, Y.; Yamaza, T.; Tuan, R.S.; Wang, S.; Shi, S.; Huang, G.T. Characterization of the apical papilla and its residing stem cells from human immature permanent teeth: A pilot study. J. Endod. 2008, 34, 166–171. [Google Scholar] [CrossRef] [Green Version]
- Wada, N.; Menicanin, D.; Shi, S.; Bartold, P.M.; Gronthos, S. Immunomodulatory properties of human periodontal ligament stem cells. J. Cell. Physiol. 2009, 219, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Medzhitov, R. Toll-like receptor control of the adaptive immune responses. Nat. Immunol. 2004, 5, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.; Bao, L.; Gu, Z.; Zhou, Q.; Liang, Y.; Zheng, Y.; Xu, Y.; Zhang, X.; Feng, X. Comparison of immunomodulatory properties of exosomes derived from bone marrow mesenchymal stem cells and dental pulp stem cells. Immunol. Res. 2019, 67, 432–442. [Google Scholar] [CrossRef] [PubMed]
- Lo Monaco, M.; Merckx, G.; Ratajczak, J.; Gervois, P.; Hilkens, P.; Clegg, P.; Bronckaers, A.; Vandeweerd, J.M.; Lambrichts, I. Stem Cells for Cartilage Repair: Preclinical Studies and Insights in Translational Animal Models and Outcome Measures. Stem Cells Int. 2018, 2018, 9079538. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Li, Y. A Molecular Cascade Underlying Articular Cartilage Degeneration. Curr. Drug Targets 2020, 21, 838–848. [Google Scholar] [CrossRef]
- Krych, A.J.; Saris, D.B.F.; Stuart, M.J.; Hacken, B. Cartilage Injury in the Knee: Assessment and Treatment Options. J. Am. Acad. Orthop. Surg 2020, 28, 914–922. [Google Scholar] [CrossRef]
- Andersson, J.K.; Hagert, E.; Brittberg, M. Cartilage Injuries and Posttraumatic Osteoarthritis in the Wrist: A Review. Cartilage 2021, 13, 156S–168S. [Google Scholar] [CrossRef]
- Cole, B.J.; Redondo, M.L.; Cotter, E.J. Articular Cartilage Injuries of the Knee: Patient Health Literacy, Expectations for Management, and Clinical Outcomes. Cartilage 2021, 12, 139–145. [Google Scholar] [CrossRef]
- Hu, Y.; Chen, X.; Wang, S.; Jing, Y.; Su, J. Subchondral bone microenvironment in osteoarthritis and pain. Bone Res. 2021, 9, 20. [Google Scholar] [CrossRef]
- van den Bosch, M.H.J. Osteoarthritis year in review 2020: Biology. Osteoarthr. Cartil. 2021, 29, 143–150. [Google Scholar] [CrossRef]
- Aydin, M.; Yorubulut, M.; Basarir, K.; Arikan, M.; Binnet, M.S. Matrix induced autologous chondrocyte implantation in the knee: Comparison between osteochondritis dissecans and osteonecrosis and effect of chondrocyte thickness on prognosis. Acta Orthop. Traumatol. Turc. 2020, 54, 66–73. [Google Scholar] [CrossRef]
- Niemeyer, P.; Albrecht, D.; Andereya, S.; Angele, P.; Ateschrang, A.; Aurich, M.; Baumann, M.; Bosch, U.; Erggelet, C.; Fickert, S.; et al. Autologous chondrocyte implantation (ACI) for cartilage defects of the knee: A guideline by the working group “Clinical Tissue Regeneration” of the German Society of Orthopaedics and Trauma (DGOU). Knee 2016, 23, 426–435. [Google Scholar] [CrossRef] [Green Version]
- Pareek, A.; Carey, J.L.; Reardon, P.J.; Peterson, L.; Stuart, M.J.; Krych, A.J. Long-Term Outcomes after Autologous Chondrocyte Implantation: A Systematic Review at Mean Follow-Up of 11.4 Years. Cartilage 2016, 7, 298–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aae, T.F.; Randsborg, P.H.; Luras, H.; Aroen, A.; Lian, O.B. Microfracture is more cost-effective than autologous chondrocyte implantation: A review of level 1 and level 2 studies with 5 year follow-up. Knee Surg. Sports Traumatol. Arthrosc. 2018, 26, 1044–1052. [Google Scholar] [CrossRef] [Green Version]
- Lo Monaco, M.; Gervois, P.; Beaumont, J.; Clegg, P.; Bronckaers, A.; Vandeweerd, J.M.; Lambrichts, I. Therapeutic Potential of Dental Pulp Stem Cells and Leukocyte- and Platelet-Rich Fibrin for Osteoarthritis. Cells 2020, 9, 980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acri, T.M.; Shin, K.; Seol, D.; Laird, N.Z.; Song, I.; Geary, S.M.; Chakka, J.L.; Martin, J.A.; Salem, A.K. Tissue Engineering for the Temporomandibular Joint. Adv. Healthc. Mater. 2019, 8, e1801236. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wu, J.S.; Tang, Y.L.; Tang, Y.J.; Fei, W.; Liang, X.H. Multiple Treatment Meta-Analysis of Intra-Articular Injection for Temporomandibular Osteoarthritis. J. Oral Maxillofac. Surg. 2020, 78, 373.e1–373.e18. [Google Scholar] [CrossRef]
- Fang, L.; Ye, Y.; Tan, X.; Huang, L.; He, Y. Overloading stress-induced progressive degeneration and self-repair in condylar cartilage. Ann. N. Y. Acad. Sci. 2021, 1503, 72–87. [Google Scholar] [CrossRef]
- Kuroda, S.; Tanimoto, K.; Izawa, T.; Fujihara, S.; Koolstra, J.H.; Tanaka, E. Biomechanical and biochemical characteristics of the mandibular condylar cartilage. Osteoarthr. Cartil. 2009, 17, 1408–1415. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.D.; Zhang, J.N.; Gan, Y.H.; Zhou, Y.H. Current understanding of pathogenesis and treatment of TMJ osteoarthritis. J. Dent. Res. 2015, 94, 666–673. [Google Scholar] [CrossRef]
- Ogasawara, N.; Kano, F.; Hashimoto, N.; Mori, H.; Liu, Y.; Xia, L.; Sakamaki, T.; Hibi, H.; Iwamoto, T.; Tanaka, E.; et al. Factors secreted from dental pulp stem cells show multifaceted benefits for treating experimental temporomandibular joint osteoarthritis. Osteoarthr. Cartil. 2020, 28, 831–841. [Google Scholar] [CrossRef] [PubMed]
- Denzer, K.; Kleijmeer, M.J.; Heijnen, H.F.; Stoorvogel, W.; Geuze, H.J. Exosome: From internal vesicle of the multivesicular body to intercellular signaling device. J. Cell Sci. 2000, 113 Pt 19, 3365–3374. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.W.; Kim, J.W.; Cha, I.H. Development of animal model for Bisphosphonates-related osteonecrosis of the jaw (BRONJ). Maxillofac. Plast. Reconstr. Surg. 2015, 37, 18. [Google Scholar] [CrossRef] [Green Version]
- Ferneini, E.M. Medication-Related Osteonecrosis of the Jaw (MRONJ). J. Oral Maxillofac. Surg. 2021, 79, 1801–1802. [Google Scholar] [CrossRef] [PubMed]
- Bukowski, J.F.; Dascher, C.C.; Das, H. Alternative bisphosphonate targets and mechanisms of action. Biochem. Biophys. Res. Commun. 2005, 328, 746–750. [Google Scholar] [CrossRef] [PubMed]
- Omi, M.; Mishina, Y. Role of osteoclasts in oral homeostasis and jawbone diseases. Oral Sci. Int. 2020, 18, 14–27. [Google Scholar] [CrossRef]
- Osaka, R.; Kato, H.; Hamada, Y.; Fujimoto, Y.; Mizusawa, N.; Watanabe, D.; Kaneko, A. Clinicostatistical analyses of medication-related osteonecrosis of the jaws (MRONJ): Evaluation of the treatment method and prognosis. Oral Sci. Int. 2021, 18, 184–192. [Google Scholar] [CrossRef]
- Kamaguchi, M.; Ohga, N.; Hata, H.; Yoshikawa, K.; Sakata, K.I.; Asaka, T.; Sato, J.; Kitagawa, Y. Treatment of medication-related osteonecrosis of the jaw using segmental mandibulectomy and a reconstruction plate: A case report and 5-year follow-up. Oral Sci. Int. 2022, 19, 52–58. [Google Scholar] [CrossRef]
- Abe, F.; Takahashi, H.; Tanaka, A. Investigation on the Action and Effect of Culture Supernatant of Human Dental Pulp Stem Cells Using Rats with Medication-Related Osteonecrosis of the Jaw. J. Hard Tissue Biol. 2019, 28, 349–357. [Google Scholar] [CrossRef] [Green Version]
- Kushiro, H.; Takahashi, H.; Tanaka, A. Effects of the prevention of medication-related osteonecrosis of the jaw by local administration of a dental pulp stem cell-conditioned medium to the rat tooth extraction socket. Odontol. Soc. Nippon Dent. Univ. 2021, 109, 836–844. [Google Scholar] [CrossRef]
- Bronckaers, A.; Hilkens, P.; Fanton, Y.; Struys, T.; Gervois, P.; Politis, C.; Martens, W.; Lambrichts, I. Angiogenic properties of human dental pulp stem cells. PLoS ONE 2013, 8, e71104. [Google Scholar] [CrossRef] [Green Version]
- Ogata, K.; Matsumura, M.; Moriyama, M.; Katagiri, W.; Hibi, H.; Nakamura, S. Cytokine Mixtures Mimicking Secretomes From Mesenchymal Stem Cells Improve Medication-Related Osteonecrosis of the Jaw in a Rat Model. JBMR Plus 2018, 2, 69–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumi, K.; Abe, T.; Kunimatsu, R.; Oki, N.; Tsuka, Y.; Awada, T.; Nakajima, K.; Ando, K.; Tanimoto, K. The effect of mesenchymal stem cells on chemotaxis of osteoclast precursor cells. J. Oral Sci. 2018, 60, 221–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubio-Almanza, M.; Camara-Gomez, R.; Merino-Torres, J.F. Obesity and type 2 diabetes: Also linked in therapeutic options. Endocrinol. Diabetes Nutr. 2019, 66, 140–149. [Google Scholar] [CrossRef]
- Sims, E.K.; Carr, A.L.J.; Oram, R.A.; DiMeglio, L.A.; Evans-Molina, C. 100 years of insulin: Celebrating the past, present and future of diabetes therapy. Nat. Med. 2021, 27, 1154–1164. [Google Scholar] [CrossRef]
- Tsoutsouki, J.; Wunna, W.; Chowdhury, A.; Chowdhury, T.A. Advances in the management of diabetes: Therapies for type 2 diabetes. Postgrad. Med. J. 2020, 96, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, D.; Papanas, N.; Schnell, O.; Nguyen, B.D.T.; Nguyen, K.T.; Kulkantrakorn, K.; Deerochanawong, C. Current concepts in the management of diabetic polyneuropathy. J. Diabetes Investig. 2021, 12, 464–475. [Google Scholar] [CrossRef]
- Patel, K.; Horak, H.; Tiryaki, E. Diabetic neuropathies. Muscle Nerve 2021, 63, 22–30. [Google Scholar] [CrossRef]
- Diabetes, C.; Complications Trial Research, G.; Nathan, D.M.; Genuth, S.; Lachin, J.; Cleary, P.; Crofford, O.; Davis, M.; Rand, L.; Siebert, C. The effect of intensive treatment of diabetes on the development and progression of long-term complications in insulin-dependent diabetes mellitus. N. Engl. J. Med. 1993, 329, 977–986. [Google Scholar] [CrossRef]
- Gaede, P.; Vedel, P.; Larsen, N.; Jensen, G.V.; Parving, H.H.; Pedersen, O. Multifactorial intervention and cardiovascular disease in patients with type 2 diabetes. N. Engl. J. Med. 2003, 348, 383–393. [Google Scholar] [CrossRef] [Green Version]
- Izumoto-Akita, T.; Tsunekawa, S.; Yamamoto, A.; Uenishi, E.; Ishikawa, K.; Ogata, H.; Iida, A.; Ikeniwa, M.; Hosokawa, K.; Niwa, Y.; et al. Secreted factors from dental pulp stem cells improve glucose intolerance in streptozotocin-induced diabetic mice by increasing pancreatic beta-cell function. BMJ Open Diabetes Res. Care 2015, 3, e000128. [Google Scholar] [CrossRef] [Green Version]
- Brissova, M.; Aamodt, K.; Brahmachary, P.; Prasad, N.; Hong, J.Y.; Dai, C.; Mellati, M.; Shostak, A.; Poffenberger, G.; Aramandla, R.; et al. Islet microenvironment, modulated by vascular endothelial growth factor-A signaling, promotes beta cell regeneration. Cell Metab. 2014, 19, 498–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elghazi, L.; Rachdi, L.; Weiss, A.J.; Cras-Meneur, C.; Bernal-Mizrachi, E. Regulation of beta-cell mass and function by the Akt/protein kinase B signalling pathway. Diabetes Obes. Metab. 2007, 9, 147–157. [Google Scholar] [CrossRef]
- Matei, I.V.; Ii, H.; Yaegaki, K. Hydrogen sulfide enhances pancreatic beta-cell differentiation from human tooth under normal and glucotoxic conditions. Regen. Med. 2017, 12, 125–141. [Google Scholar] [CrossRef]
- Makinen, K.; Manninen, H.; Hedman, M.; Matsi, P.; Mussalo, H.; Alhava, E.; Yla-Herttuala, S. Increased vascularity detected by digital subtraction angiography after VEGF gene transfer to human lower limb artery: A randomized, placebo-controlled, double-blinded phase II study. Mol. Ther. 2002, 6, 127–133. [Google Scholar] [CrossRef]
- Ropper, A.H.; Gorson, K.C.; Gooch, C.L.; Weinberg, D.H.; Pieczek, A.; Ware, J.H.; Kershen, J.; Rogers, A.; Simovic, D.; Schratzberger, P.; et al. Vascular endothelial growth factor gene transfer for diabetic polyneuropathy: A randomized, double-blinded trial. Ann. Neurol. 2009, 65, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Makino, E.; Nakamura, N.; Miyabe, M.; Ito, M.; Kanada, S.; Hata, M.; Saiki, T.; Sango, K.; Kamiya, H.; Nakamura, J.; et al. Conditioned media from dental pulp stem cells improved diabetic polyneuropathy through anti-inflammatory, neuroprotective and angiogenic actions: Cell-free regenerative medicine for diabetic polyneuropathy. J. Diabetes Investig. 2019, 10, 1199–1208. [Google Scholar] [CrossRef]
- Kanada, S.; Makino, E.; Nakamura, N.; Miyabe, M.; Ito, M.; Hata, M.; Yamauchi, T.; Sawada, N.; Kondo, S.; Saiki, T.; et al. Direct Comparison of Therapeutic Effects on Diabetic Polyneuropathy between Transplantation of Dental Pulp Stem Cells and Administration of Dental Pulp Stem Cell-Secreted Factors. Int. J. Mol. Sci. 2020, 21, 64. [Google Scholar] [CrossRef] [PubMed]
- Hata, M.; Omi, M.; Kobayashi, Y.; Nakamura, N.; Miyabe, M.; Ito, M.; Ohno, T.; Imanishi, Y.; Himeno, T.; Kamiya, H.; et al. Sustainable Effects of Human Dental Pulp Stem Cell Transplantation on Diabetic Polyneuropathy in Streptozotocine-Induced Type 1 Diabetes Model Mice. Cells 2021, 10, 2473. [Google Scholar] [CrossRef]
- Olsburgh, S.; Jacoby, T.; Krejci, I. Crown fractures in the permanent dentition: Pulpal and restorative considerations. Dent. Traumatol. 2002, 18, 103–115. [Google Scholar] [CrossRef] [Green Version]
- Jones, L.C. Dental Trauma. Oral Maxillofac. Surg. Clin. N. Am. 2020, 32, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Laschke, M.W.; Harder, Y.; Amon, M.; Martin, I.; Farhadi, J.; Ring, A.; Torio-Padron, N.; Schramm, R.; Rucker, M.; Junker, D.; et al. Angiogenesis in tissue engineering: Breathing life into constructed tissue substitutes. Tissue Eng. 2006, 12, 2093–2104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunawardena, T.N.A.; Rahman, M.T.; Abdullah, B.J.J.; Abu Kasim, N.H. Conditioned media derived from mesenchymal stem cell cultures: The next generation for regenerative medicine. J. Tissue Eng. Regen. Med. 2019, 13, 569–586. [Google Scholar] [CrossRef] [PubMed]
- Pawitan, J.A. Prospect of stem cell conditioned medium in regenerative medicine. Biomed. Res. Int. 2014, 2014, 965849. [Google Scholar] [CrossRef] [Green Version]
- de Cara, S.; Origassa, C.S.T.; de Sa Silva, F.; Moreira, M.; de Almeida, D.C.; Pedroni, A.C.F.; Carvalho, G.L.; Cury, D.P.; Camara, N.O.S.; Marques, M.M. Angiogenic properties of dental pulp stem cells conditioned medium on endothelial cells in vitro and in rodent orthotopic dental pulp regeneration. Heliyon 2019, 5, e01560. [Google Scholar] [CrossRef] [Green Version]
- Sarra, G.; Machado, M.E.L.; Caballero-Flores, H.V.; Moreira, M.S.; Pedroni, A.C.F.; Marques, M.M. Effect of human dental pulp stem cell conditioned medium in the dentin-pulp complex regeneration: A pilot in vivo study. Tissue Cell 2021, 72, 101536. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367. [Google Scholar] [CrossRef]
- Cai, Y.; Li, J.; Jia, C.; He, Y.; Deng, C. Therapeutic applications of adipose cell-free derivatives: A review. Stem Cell Res. Ther. 2020, 11, 312. [Google Scholar] [CrossRef]
- Chen, X.; Chen, A.; Woo, T.L.; Choo, A.B.; Reuveny, S.; Oh, S.K. Investigations into the metabolism of two-dimensional colony and suspended microcarrier cultures of human embryonic stem cells in serum-free media. Stem Cells Dev. 2010, 19, 1781–1792. [Google Scholar] [CrossRef]


{kind=link}
| Anti-Inflammatory Factors (Intensity) | ||
| BMMSC-CM | DPSC-CM | |
| Follistatin | 2328 | 36241 |
| TGF-β1 | 2186 | 11623 |
| IL-10 | 234 | 7989 |
| IL-13 | 80 | 5098 |
| VEGF | 1242 | 3549 |
| IGF-1 | 3324 | 3521 |
| HGF | 0 | 1857 |
| TECK | 1513 | 1609 |
| MCP-1 | 483 | 965 |
| IL-29 | 1019 | 943 |
| Adiponectin | 0 | 502 |
| ED-Siglec-9 | 0 | 396 |
| GM-CSF | 188 | 159 |
| Secreted Factors Only in DPSC-CM (Intensity) | ||
| BMMSC-CM | DPSC-CM | |
| HGF | 0 | 1857 |
| NCAM-1 | 0 | 573 |
| Adiponectin | 0 | 502 |
| ED-Siglec-9 | 0 | 396 |
| MMP-13 | 0 | 377 |
| NT-3 | 0 | 249 |
| BDNF | 0 | 179 |
| MMP-9 | 0 | 136 |
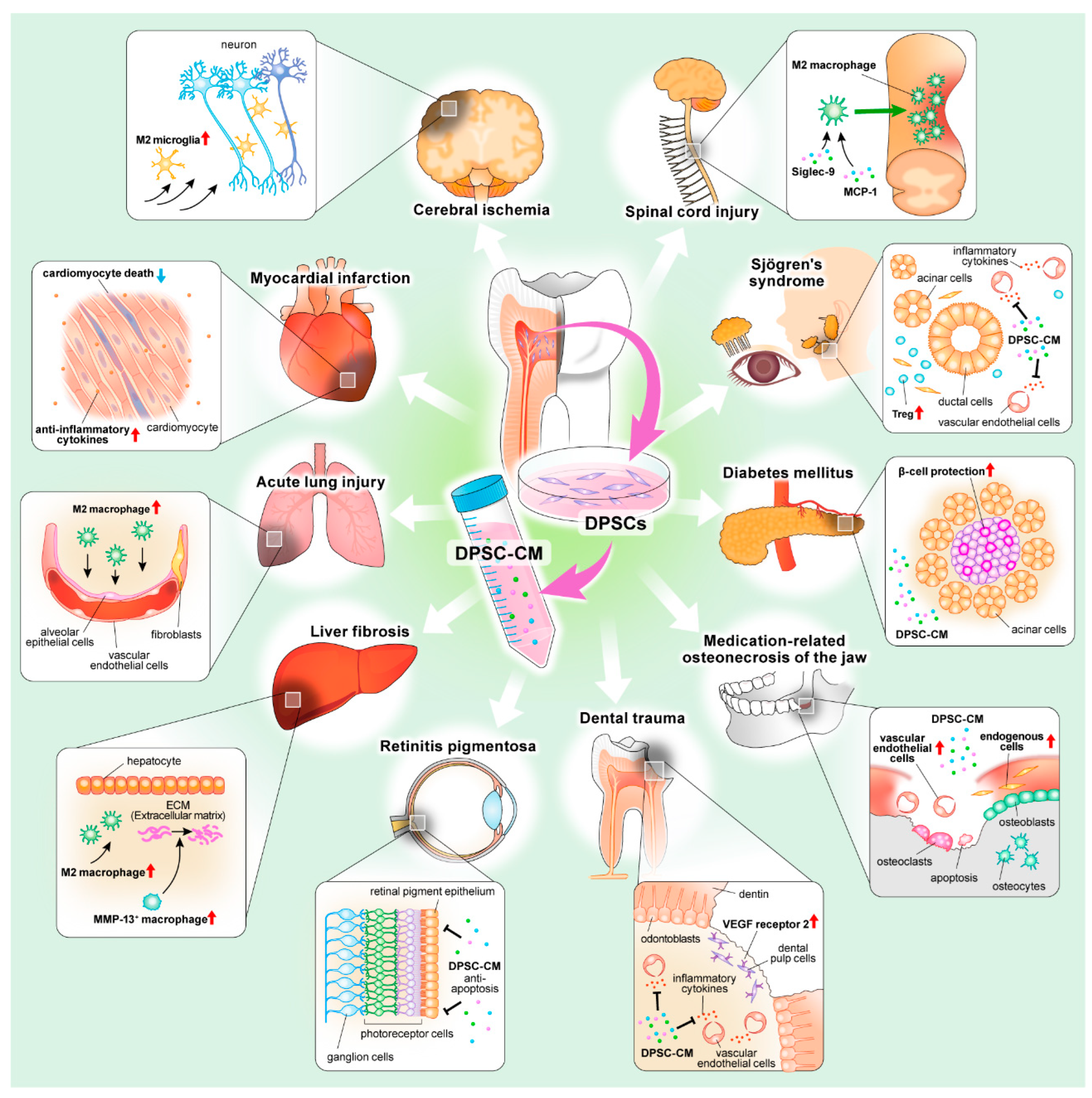
| Type | Disease | Animal Model | Administration Method | Mechanism | Reference |
|---|---|---|---|---|---|
| Cerebrovascular disease | Forcal cerebral ischemia | Cerebral ischemia model (Sprague-Dawley rats) | Intranasal injection | Induction of activated M2-type microglia | Inoue T et al. [32] |
| Aneurysmal subarachnoid hemorrhage | Experimental aSAH model (Wistar rats) | Intrathecal injection | Inhibitor for MMP-3 and 9 by TIMP-2 | Chen TF et al. [35] | |
| Heart disease | Myocardial infarction | Myocardial ischemia-reperfusion injury model (C57BL/6J mice) | Intravenous injection | Reduction of cardiomyocyte death and suppression of inflammatory responses | Yamaguchi S et al. [46] |
| Lung disease | Acute lung injury | Bleomycin-induced acute lung injury model (C57BL/6J mice) | Intravenous injection | Induction of M2 macrophage differentiation | Wakayama H et al. [53] |
| Liver disease | Liver fibrosis | Carbon tetrachloride (CCl4)-induced liver fbrosis model (C57BL/6J mice) | Intravenous injection | Induction of the MMP-13+ restorative hepatic macrophages | Hirata M et al. [61] |
| Acute liver failure | D-galactosamine-induced acute liver failure model (Sprague-Dawley rats) | Intravenous injection | Induction of an anti-inflammatory M2 environment and activation of adult LPCs | Matsushita Y et al. [62] | |
| Eye disease | Retinitis pigmentosa | C57BL/6 J mice with RPGR knockout | Subretinal injection | Anti-apoptotic activity | Li XX et al. [74] |
| Neurological disorder | Spinal cord injury | Contusion model of spinal cord injury (Sprague-Dawley rats) | Injection into the injury epicenter | Induction of an M2-dominant neurorepairing microenvironment by ED-Siglec-9 and MCP-1 | Asadi-Golshan R et al. [82] Matsubara K et al. [83] |
| Facial nerve injury | Facial nerves transection model (Sprague-Dawley rats) | Local implantation with an atelocollagen sponge | Induction of the polarization of M2 macrophages by MCP-1/Siglec-9 | Kano F et al. [84] | |
| Parkinson’s disease | Rotenone-induced Parkinson’s disease model (Lewis rats) | Intravenous injection | Upregulation of tyrosine hydroxylase expression | Chen YR et al. [87] | |
| Alzheimer’s disease | Aβ1-40 peptide-induced Alzheimer’s disease model (imprinting control region mice) | Intranasal injection | Activated M2-type microglia | Mita T et al. [92] | |
| Amyotrophic lateral sclerosis | Transgenic mice (B6SJL-Tg (SOD1G93A)1 Gur/J) expressing the humansuperoxide dismutase 1 (mSOD1G93A) mutation | Intraperitoneal injection | Directly protection of motor neurons | Wang J et al. [94] | |
| Multiple sclerosis | Experimental autoimmune encephalomyelitis (EAE) model (C57BL/6J mice) | Intravenous injection | Induction of M2 macrophage polarization through ED-Siglec-9 by interacting with the sialic acid-bound CCR2 | Shimojima C et al. [96] | |
| Autoimmune disease | Sjögren’s syndrome | Nonobese diabetic (NOD) mice | Intravenous injection | Induction of Tregs throug the TGF-β/Smad pathway | Matsumura-Kawashima M et al. [30] Ogata K et al. [31] |
| Bone and Cartilage disease | Osteoarthritis | Mouse mechanical stress-induced osteoarthritis model (Institute of Cancer Research mice) | Local injection | Stimulation of endogenous cells to proliferate and replace the lost tissue | Lo Monaco M et al. [115] Ogasawara N et al. [121] |
| Medication-related osteonecrosis of the jaw (MRONJ) | Zoledronate-induced MRONJ model (Wistar rats) | Local injection | Induced epithelial tissue formation by increasing vascular endothelial cells | Abe F et al. [129] Kushiro H et al. [130] | |
| Metabolic disease | Diabetes mellitus | Streptozotocin-induced diabetic mice (C57BL/6J mice) | Intravenous injection | Protection and encouragment of the propagation for β-cells | Izumoto-Akita T et al. [141] |
| Diabetic polyneuropathy | Streptozotocin-induced diabetic rats (Sprague-Dawley rats) Streptozotocin-induced diabetic mice (C57BL/6J mice) | Local injection into the hindlimb skeletal muscles | Angiogenesis and improvement of blood flow, anti-inflammatory action, and neuroprotective action | Makino E et al. [147] Kanada S et al. [148] Hata M et al. [149] | |
| Dental disease | Dental trauma | Orthotopic transplantation model (Wistar rats) | Local injection with a collagen | Immunopositivity reaction for VEGF receptor 2, and mainly controlling pulp tissue inflammation | de Cara S et al. [155] Sarra G et al. [156] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogata, K.; Moriyama, M.; Matsumura-Kawashima, M.; Kawado, T.; Yano, A.; Nakamura, S. The Therapeutic Potential of Secreted Factors from Dental Pulp Stem Cells for Various Diseases. Biomedicines 2022, 10, 1049. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10051049
Ogata K, Moriyama M, Matsumura-Kawashima M, Kawado T, Yano A, Nakamura S. The Therapeutic Potential of Secreted Factors from Dental Pulp Stem Cells for Various Diseases. Biomedicines. 2022; 10(5):1049. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10051049
Chicago/Turabian StyleOgata, Kenichi, Masafumi Moriyama, Mayu Matsumura-Kawashima, Tatsuya Kawado, Aiko Yano, and Seiji Nakamura. 2022. "The Therapeutic Potential of Secreted Factors from Dental Pulp Stem Cells for Various Diseases" Biomedicines 10, no. 5: 1049. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10051049