
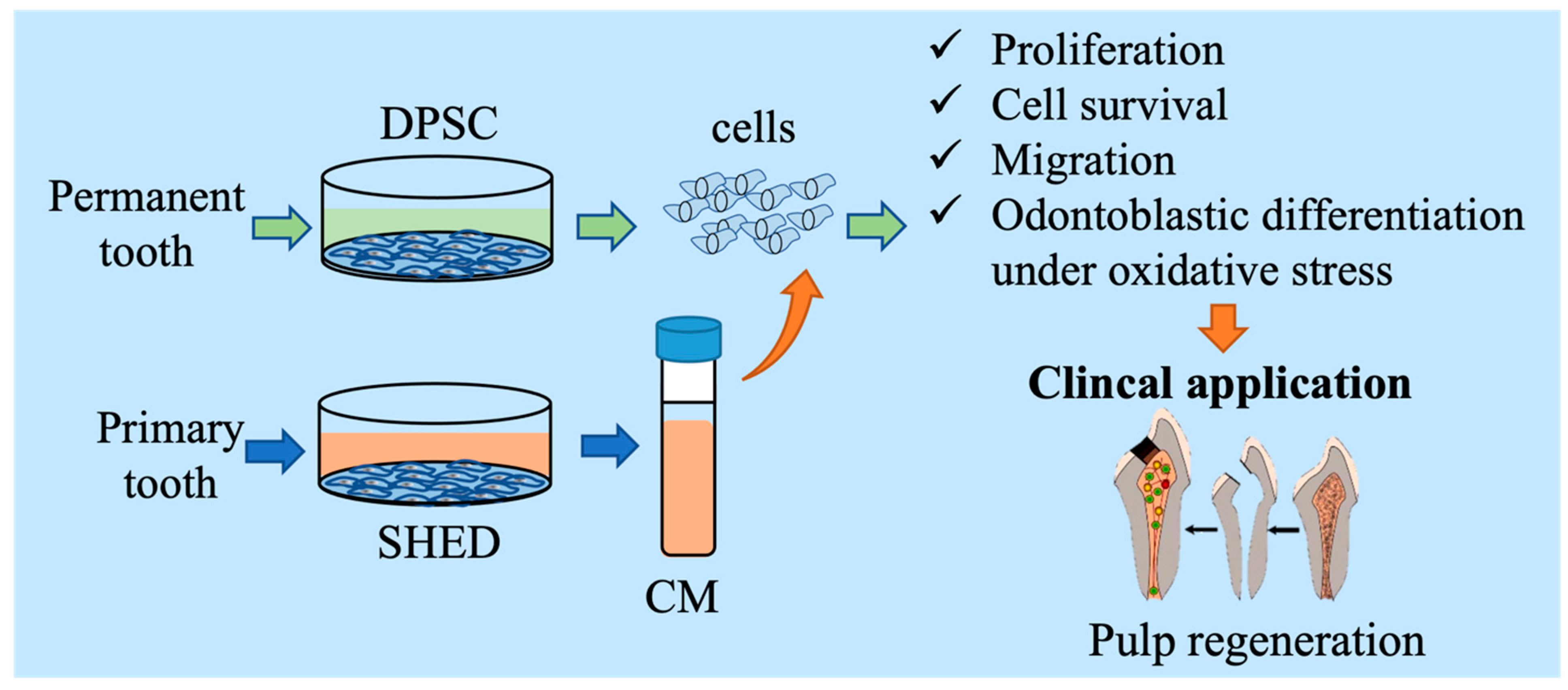
Investigating the Effects of Conditioned Media from Stem Cells of Human Exfoliated Deciduous Teeth on Dental Pulp Stem Cells

 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
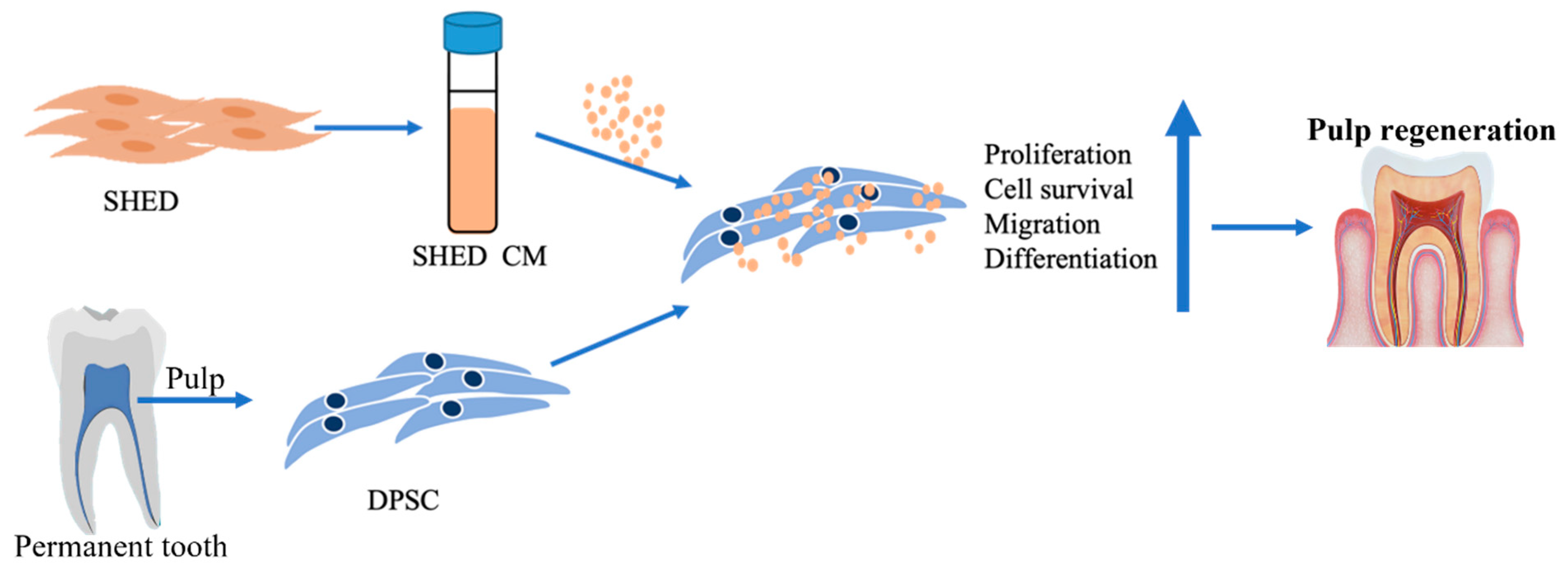
Abstract
:1. Introduction
2. Materials and Methods
2.1. Primary Culture and Expansion of Dental Pulp Stem Cell
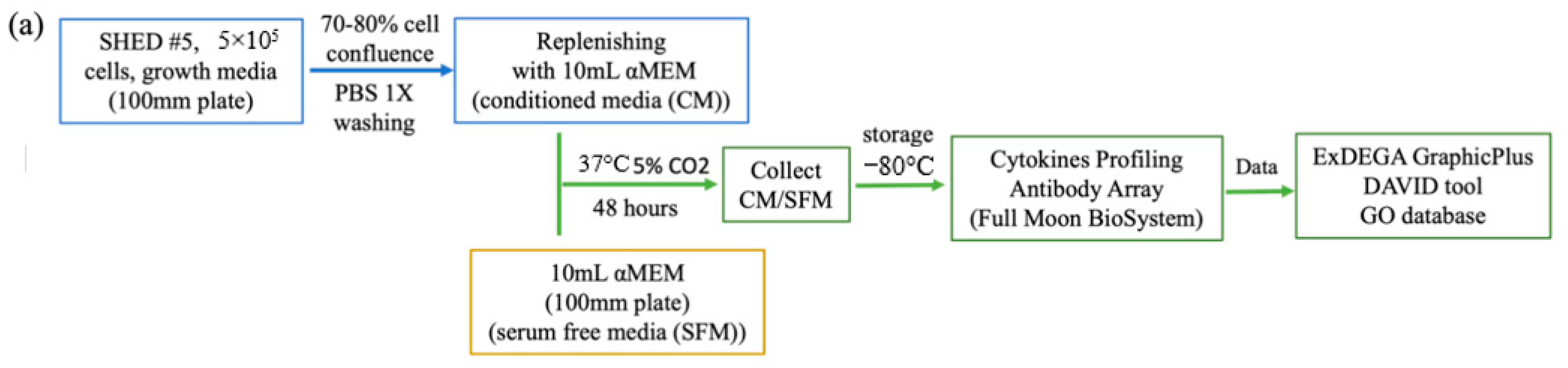
2.2. Preparation of Conditioned Medium
2.3. Proliferation Assays
2.4. Cell migration Assay
2.5. Antioxidative Stress
2.6. Osteogenic Differentiation Assays
2.7. Cytokine Profiling
2.8. Quantitative Real Time Polymerase Chain Reaction (qRT–PCR)
2.9. Statistical Analyses
3. Results
3.1. Characteristics of DPSCs and SHEDs
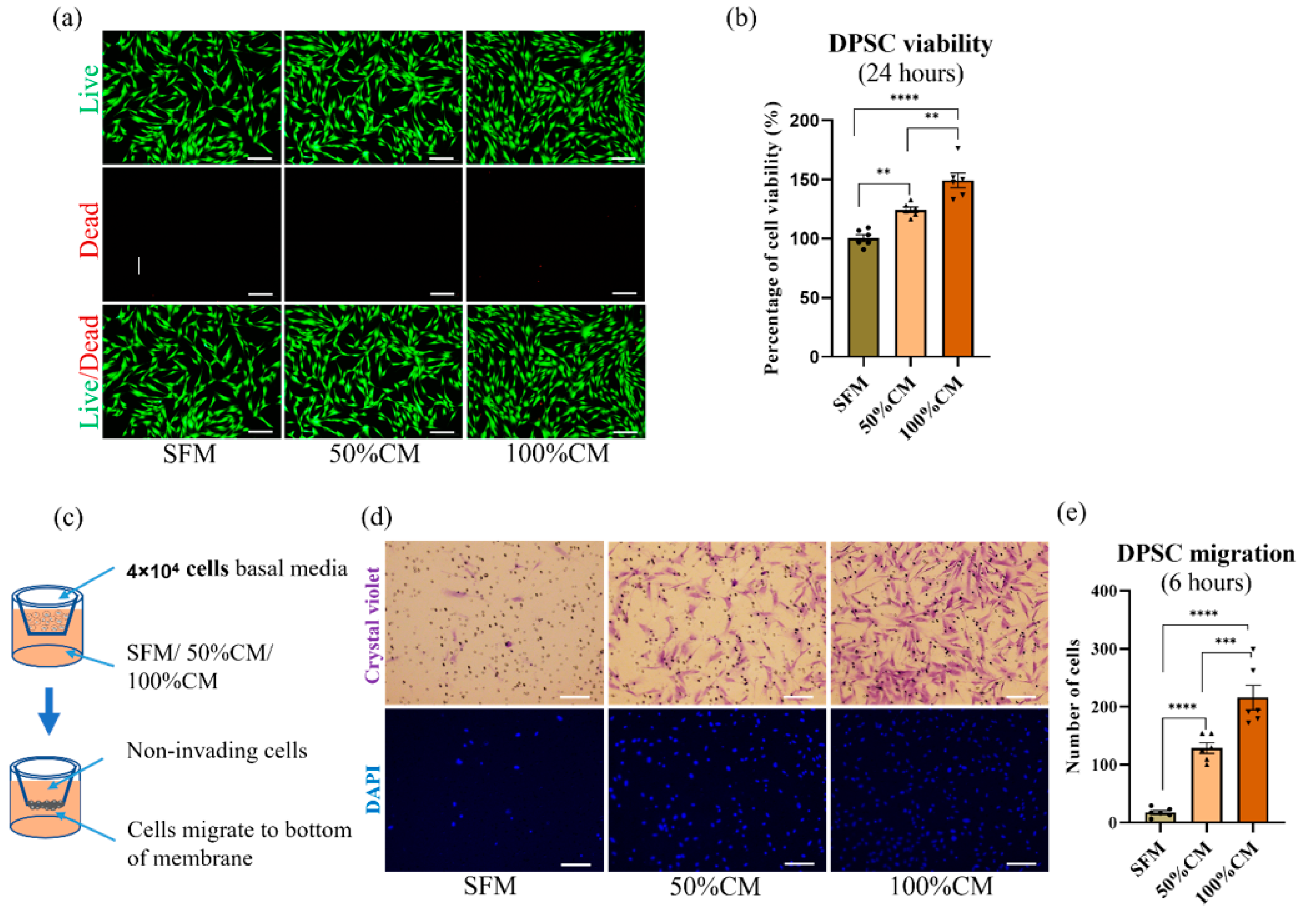
3.2. Effect of CM on Cell Viability and Proliferation
3.3. SHED CM Attracted DPSC Migration
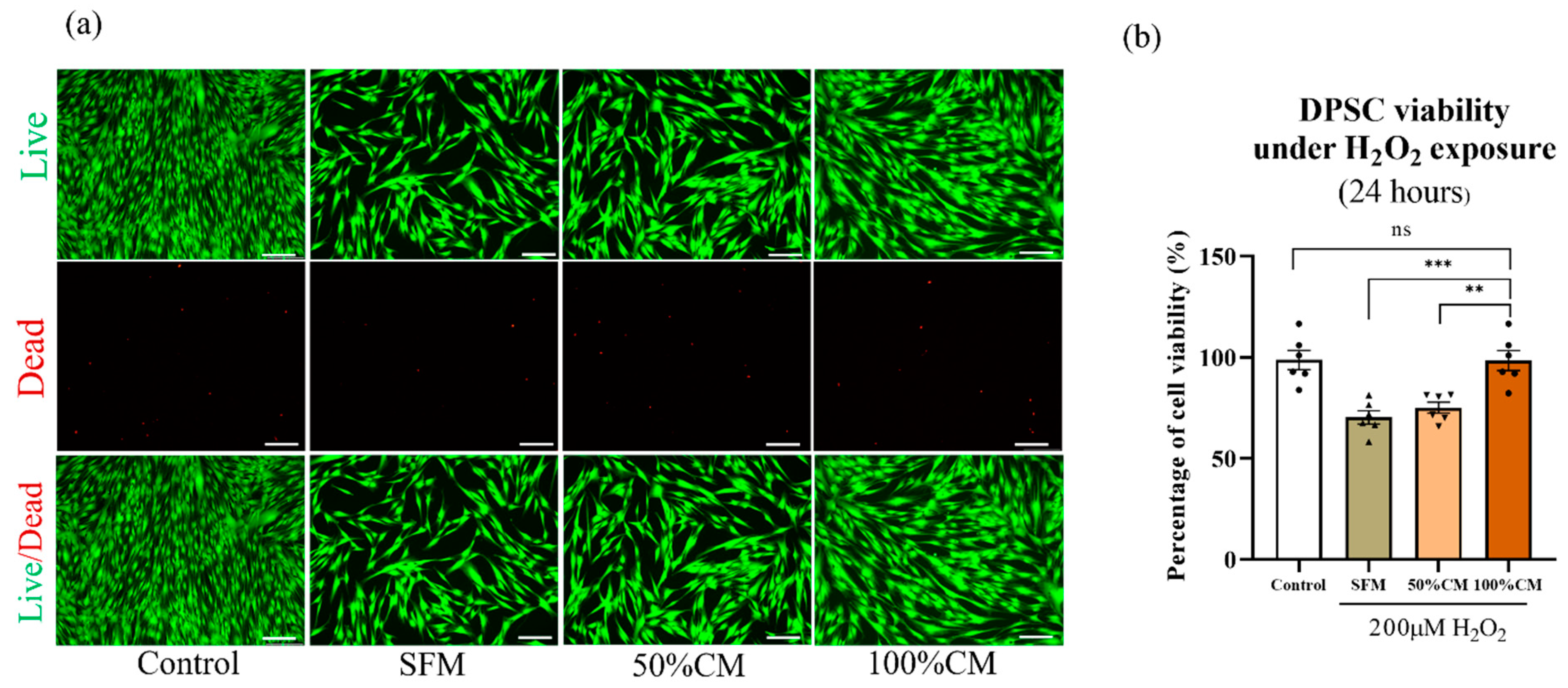
3.4. Protective Effect of SHED CM against H2O2-Induced Cell Death on DPSCs
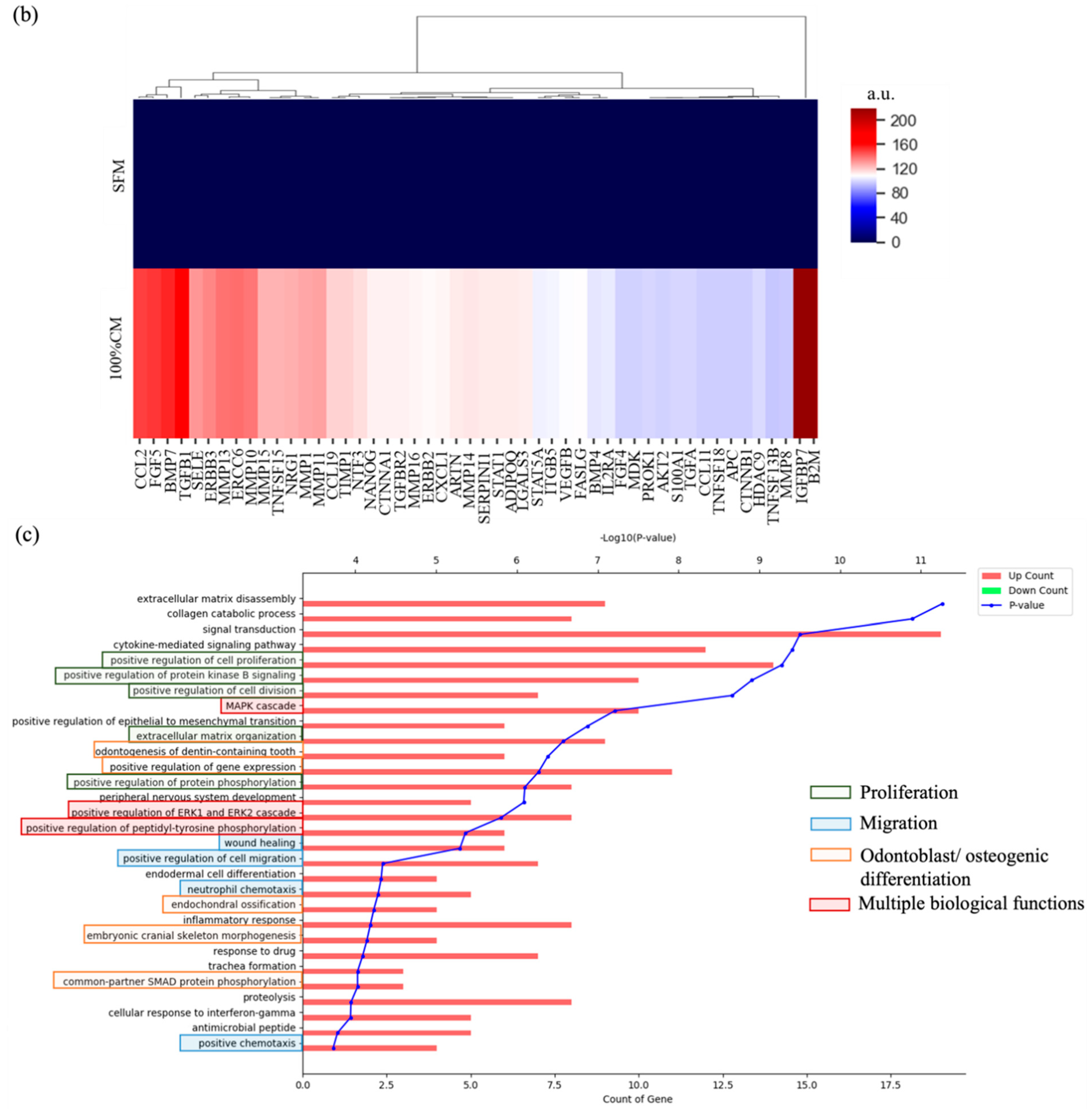
3.5. Cytokine Profiling Results
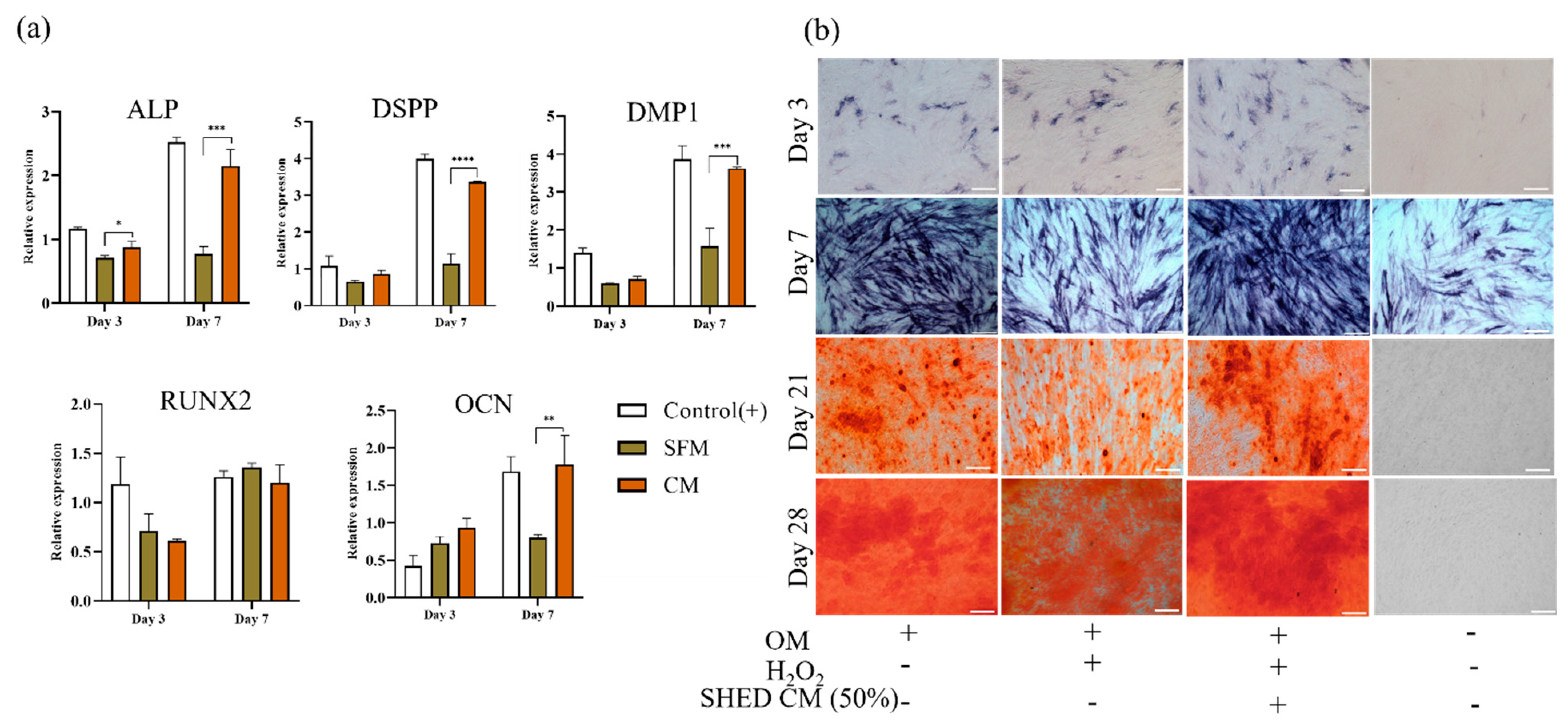
3.6. Odontoblast/Osteoblast Induction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Miura, M.; Gronthos, S.; Zhao, M.; Lu, B.; Fisher, L.W.; Robey, P.G.; Shi, S. SHED: Stem cells from human exfoliated deciduous teeth. Proc. Natl. Acad. Sci. USA 2003, 100, 5807–5812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, Q.; Dong, Z.; Wang, W.; Li, B.; Jin, Y. Dental stem cell and dental tissue regeneration. Front. Med. 2019, 13, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Wu, Y.; Smales, R.J. Identifying and reducing risks for potential fractures in endodontically treated teeth. J. Endod. 2010, 36, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Khasnis, S.A.; Kidiyoor, K.H.; Patil, A.B.; Kenganal, S.B. Vertical root fractures and their management. J. Conserv. Dent. 2014, 17, 103–110. [Google Scholar] [CrossRef]
- Greta, D.C.; Colosi, H.A.; Gasparik, C.; Dudea, D. Color comparison between non-vital and vital teeth. J. Adv. Prosthodont. 2018, 10, 218–226. [Google Scholar] [CrossRef] [Green Version]
- Krastl, G.; Allgayer, N.; Lenherr, P.; Filippi, A.; Taneja, P.; Weiger, R. Tooth discoloration induced by endodontic materials: A literature review. Dent. Traumatol. 2013, 29, 2–7. [Google Scholar] [CrossRef]
- Li, Z.; Liu, L.; Wang, L.; Song, D. The effects and potential applications of concentrated growth factor in dentin–pulp complex regeneration. Stem Cell Res. Ther. 2021, 12, 357. [Google Scholar] [CrossRef]
- Ling, L.; Zhao, Y.M.; Wang, X.T.; Wen, Q.; Ge, L.H. Regeneration of Dental Pulp Tissue by Autologous Grafting Stem Cells Derived from Inflammatory Dental Pulp Tissue in Immature Premolars in a Beagle Dog. Chin. J. Dent. Res. 2020, 23, 143–150. [Google Scholar] [CrossRef]
- Jung, C.; Kim, S.; Sun, T.; Cho, Y.B.; Song, M. Pulp-dentin regeneration: Current approaches and challenges. J. Tissue Eng. 2019, 10, 2041731418819263. [Google Scholar] [CrossRef] [Green Version]
- Nakashima, M.; Iohara, K.; Murakami, M.; Nakamura, H.; Sato, Y.; Ariji, Y.; Matsushita, K. Pulp regeneration by transplantation of dental pulp stem cells in pulpitis: A pilot clinical study. Stem Cell Res. Ther. 2017, 8, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xuan, K.; Li, B.; Guo, H.; Sun, W.; Kou, X.; He, X.; Zhang, Y.; Sun, J.; Liu, A.; Liao, L.; et al. Deciduous autologous tooth stem cells regenerate dental pulp after implantation into injured teeth. Sci. Transl. Med. 2018, 10, eaaf3227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cehreli, Z.C.; Unverdi, G.E.; Ballikaya, E. Deciduous Tooth Pulp Autotransplantation for the Regenerative Endodontic Treatment of Permanent Teeth with Pulp Necrosis: A Case Series. J. Endod. 2022. S0099-2399(22)00068-1. [Google Scholar] [CrossRef] [PubMed]
- Meza, G.; Urrejola, D.; Saint Jean, N.; Inostroza, C.; López, V.; Khoury, M.; Brizuela, C. Personalized Cell Therapy for Pulpitis Using Autologous Dental Pulp Stem Cells and Leukocyte Platelet-rich Fibrin: A Case Report. J. Endod. 2019, 45, 144–149. [Google Scholar] [CrossRef]
- Mattei, V.; Martellucci, S.; Pulcini, F.; Santilli, F.; Sorice, M.; Delle Monache, S. Regenerative Potential of DPSCs and Revascularization: Direct, Paracrine or Autocrine Effect? Stem Cell Rev. Rep. 2021, 17, 1635–1646. [Google Scholar] [CrossRef]
- Iezzi, I.; Pagella, P.; Mattioli-Belmonte, M.; Mitsiadis, T.A. The effects of ageing on dental pulp stem cells, the tooth longevity elixir. Eur. Cell Mater. 2019, 37, 175–185. [Google Scholar] [CrossRef]
- Wu, W.; Zhou, J.; Xu, C.T.; Zhang, J.; Jin, Y.J.; Sun, G.L. Derivation and growth characteristics of dental pulp stem cells from patients of different ages. Mol. Med. Rep. 2015, 12, 5127–5134. [Google Scholar] [CrossRef] [Green Version]
- Iohara, K.; Murakami, M.; Nakata, K.; Nakashima, M. Age-dependent decline in dental pulp regeneration after pulpectomy in dogs. Exp. Gerontol. 2014, 52, 39–45. [Google Scholar] [CrossRef]
- Yi, Q.; Liu, O.; Yan, F.; Lin, X.; Diao, S.; Wang, L.; Jin, L.; Wang, S.; Lu, Y.; Fan, Z. Analysis of Senescence-Related Differentiation Potentials and Gene Expression Profiles in Human Dental Pulp Stem Cells. Cells Tissues Organs 2017, 203, 1–11. [Google Scholar] [CrossRef]
- Delle Monache, S.; Martellucci, S.; Clementi, L.; Pulcini, F.; Santilli, F.; Mei, C.; Piccoli, L.; Angelucci, A.; Mattei, V. In Vitro Conditioning Determines the Capacity of Dental Pulp Stem Cells to Function as Pericyte-Like Cells. Stem Cells Dev. 2019, 28, 695–706. [Google Scholar] [CrossRef]
- Bloom, A.B.; Zaman, M.H. Influence of the microenvironment on cell fate determination and migration. Physiol. Genom. 2014, 46, 309–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.G. Biological Molecules for the Regeneration of the Pulp-Dentin Complex. Dent. Clin. N. Am. 2017, 61, 127–141. [Google Scholar] [CrossRef] [PubMed]
- Bar, J.K.; Lis-Nawara, A.; Grelewski, P.G. Dental Pulp Stem Cell-Derived Secretome and Its Regenerative Potential. Int. J. Mol. Sci. 2021, 22, 12018. [Google Scholar] [CrossRef] [PubMed]
- Jin, Q.; Li, P.; Yuan, K.; Zhao, F.; Zhu, X.; Zhang, P.; Huang, Z. Extracellular vesicles derived from human dental pulp stem cells promote osteogenesis of adipose-derived stem cells via the MAPK pathway. J. Tissue Eng. 2020, 11, 2041731420975569. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Yoon, J.Y.; Lee, J.H.; Lee, H.H.; Knowles, J.C.; Kim, H.W. Emerging biogenesis technologies of extracellular vesicles for tissue regenerative therapeutics. J. Tissue Eng. 2021, 12, 20417314211019015. [Google Scholar] [CrossRef]
- Al-Sharabi, N.; Xue, Y.; Fujio, M.; Ueda, M.; Gjerde, C.; Mustafa, K.; Fristad, I. Bone marrow stromal cell paracrine factors direct osteo/odontogenic differentiation of dental pulp cells. Tissue Eng. Part A 2014, 20, 3063–3072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, S.; Zhao, Y.; Fang, T.J.; Ge, L. Effect of the Soluble Factors Released by Dental Apical Papilla-Derived Stem Cells on the Osteo/Odontogenic, Angiogenic, and Neurogenic Differentiation of Dental Pulp Cells. Stem Cells Dev. 2020, 29, 795–805. [Google Scholar] [CrossRef]
- Bhandi, S.; Al Khatani, A.; Abdulaziz Sumayli, H.; Yahya Sabyei, M.; Mohammed Al Zailai, A.; Ali Sumayli, M.; Ibrahim Hakami, H.; Abdurabu Jafer, M.; Vyas, N.; Ali Baeshen, H.; et al. Comparative analysis of cytokines and growth factors in the conditioned media of stem cells from the pulp of deciduous, young, and old permanent tooth. Saudi J. Biol. Sci. 2021, 28, 3559–3565. [Google Scholar] [CrossRef]
- Kang, C.-M.; Shin, M.K.; Jeon, M.; Lee, Y.-H.; Song, J.S.; Lee, J.-H. Distinctive cytokine profiles of stem cells from human exfoliated deciduous teeth and dental pulp stem cells. J. Dent. Sci. 2022, 17, 276–283. [Google Scholar] [CrossRef]
- Muhammad, S.A.; Nordin, N.; Hussin, P.; Mehat, M.Z.; Abu Kasim, N.H.; Fakurazi, S. Protective effects of stem cells from human exfoliated deciduous teeth derived conditioned medium on osteoarthritic chondrocytes. PLoS ONE 2020, 15, e0238449. [Google Scholar] [CrossRef]
- Hiraki, T.; Kunimatsu, R.; Nakajima, K.; Abe, T.; Yamada, S.; Rikitake, K.; Tanimoto, K. Stem cell-derived conditioned media from human exfoliated deciduous teeth promote bone regeneration. Oral Dis. 2020, 26, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, Y.; Ishigami, M.; Matsubara, K.; Kondo, M.; Wakayama, H.; Goto, H.; Ueda, M.; Yamamoto, A. Multifaceted therapeutic benefits of factors derived from stem cells from human exfoliated deciduous teeth for acute liver failure in rats. J. Tissue Eng. Regen. Med. 2017, 11, 1888–1896. [Google Scholar] [CrossRef] [PubMed]
- de Cara, S.P.H.M.; Origassa, C.S.T.; de Sá Silva, F.; Moreira, M.S.N.A.; de Almeida, D.C.; Pedroni, A.C.F.; Carvalho, G.L.; Cury, D.P.; Câmara, N.O.S.; Marques, M.M. Angiogenic properties of dental pulp stem cells conditioned medium on endothelial cells in vitro and in rodent orthotopic dental pulp regeneration. Heliyon 2019, 5, e01560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kichenbrand, C.; Velot, E.; Menu, P.; Moby, V. Dental Pulp Stem Cell-Derived Conditioned Medium: An Attractive Alternative for Regenerative Therapy. Tissue Eng. Part B Rev. 2019, 25, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Stanko, P.; Altanerova, U.; Jakubechova, J.; Repiska, V.; Altaner, C. Dental Mesenchymal Stem/Stromal Cells and Their Exosomes. Stem Cells Int. 2018, 2018, 8973613. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.T.; Gronthos, S.; Shi, S. Mesenchymal stem cells derived from dental tissues vs. those from other sources: Their biology and role in regenerative medicine. J. Dent. Res. 2009, 88, 792–806. [Google Scholar] [CrossRef]
- Kato, M.; Tsunekawa, S.; Nakamura, N.; Miura-Yura, E.; Yamada, Y.; Hayashi, Y.; Nakai-Shimoda, H.; Asano, S.; Hayami, T.; Motegi, M.; et al. Secreted Factors from Stem Cells of Human Exfoliated Deciduous Teeth Directly Activate Endothelial Cells to Promote All Processes of Angiogenesis. Cells 2020, 9, 2385. [Google Scholar] [CrossRef]
- Laureys, W.G.; Cuvelier, C.A.; Dermaut, L.R.; De Pauw, G.A. The critical apical diameter to obtain regeneration of the pulp tissue after tooth transplantation, replantation, or regenerative endodontic treatment. J. Endod. 2013, 39, 759–763. [Google Scholar] [CrossRef]
- Estefan, B.S.; El Batouty, K.M.; Nagy, M.M.; Diogenes, A. Influence of Age and Apical Diameter on the Success of Endodontic Regeneration Procedures. J. Endod. 2016, 42, 1620–1625. [Google Scholar] [CrossRef]
- Sugimura-Wakayama, Y.; Katagiri, W.; Osugi, M.; Kawai, T.; Ogata, K.; Sakaguchi, K.; Hibi, H. Peripheral Nerve Regeneration by Secretomes of Stem Cells from Human Exfoliated Deciduous Teeth. Stem Cells Dev. 2015, 24, 2687–2699. [Google Scholar] [CrossRef]
- Chen, F.; Liu, Y.; Wong, N.K.; Xiao, J.; So, K.F. Oxidative Stress in Stem Cell Aging. Cell Transpl. 2017, 26, 1483–1495. [Google Scholar] [CrossRef] [PubMed]
- Pervaiz, S.; Taneja, R.; Ghaffari, S. Oxidative stress regulation of stem and progenitor cells. Antioxid. Redox. Signal 2009, 11, 2777–2789. [Google Scholar] [CrossRef] [PubMed]
- Petrenko, Y.; Vackova, I.; Kekulova, K.; Chudickova, M.; Koci, Z.; Turnovcova, K.; Kupcova Skalnikova, H.; Vodicka, P.; Kubinova, S. A Comparative Analysis of Multipotent Mesenchymal Stromal Cells derived from Different Sources, with a Focus on Neuroregenerative Potential. Sci. Rep. 2020, 10, 4290. [Google Scholar] [CrossRef] [PubMed]
- Saleem, R.; Mohamed-Ahmed, S.; Elnour, R.; Berggreen, E.; Mustafa, K.; Al-Sharabi, N. Conditioned Medium from Bone Marrow Mesenchymal Stem Cells Restored Oxidative Stress-Related Impaired Osteogenic Differentiation. Int. J. Mol. Sci. 2021, 22, 13458. [Google Scholar] [CrossRef]
- Kadekar, D.; Rangole, S.; Kale, V.; Limaye, L. Conditioned Medium from Placental Mesenchymal Stem Cells Reduces Oxidative Stress during the Cryopreservation of Ex Vivo Expanded Umbilical Cord Blood Cells. PLoS ONE 2016, 11, e0165466. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Li, D.; Li, X.; Shi, Q.; Ju, X. Mesenchymal stem cell conditioned medium alleviates oxidative stress injury induced by hydrogen peroxide via regulating miR143 and its target protein in hepatocytes. BMC Immunol. 2017, 18, 51. [Google Scholar] [CrossRef]
- Jiang, F.; Xu, X.R.; Li, W.M.; Xia, K.; Wang, L.F.; Yang, X.C. Monotropein alleviates H2O2-induced inflammation, oxidative stress and apoptosis via NF-κB/AP-1 signaling. Mol. Med. Rep. 2020, 22, 4828–4836. [Google Scholar] [CrossRef]
- Ogata, K.; Katagiri, W.; Osugi, M.; Kawai, T.; Sugimura, Y.; Hibi, H.; Nakamura, S.; Ueda, M. Evaluation of the therapeutic effects of conditioned media from mesenchymal stem cells in a rat bisphosphonate-related osteonecrosis of the jaw-like model. Bone 2015, 74, 95–105. [Google Scholar] [CrossRef]
- Bai, X.C.; Lu, D.; Bai, J.; Zheng, H.; Ke, Z.Y.; Li, X.M.; Luo, S.Q. Oxidative stress inhibits osteoblastic differentiation of bone cells by ERK and NF-kappaB. Biochem. Biophys. Res. Commun. 2004, 314, 197–207. [Google Scholar] [CrossRef]
- Liang, M.; Liu, W.; Peng, Z.; Lv, S.; Guan, Y.; An, G.; Zhang, Y.; Huang, T.; Wang, Y. The therapeutic effect of secretome from human umbilical cord-derived mesenchymal stem cells in age-related osteoporosis. Artif. Cells Nanomed. Biotechnol. 2019, 47, 1357–1366. [Google Scholar] [CrossRef] [Green Version]
- Aziz, N.S.; Yusop, N.; Ahmad, A. Importance of Stem Cell Migration and Angiogenesis Study for Regenerative Cell-based Therapy: A Review. Curr. Stem Cell Res. Ther. 2020, 15, 284–299. [Google Scholar] [CrossRef] [PubMed]
- Ghannam, M.G.; Alameddine, H.; Bordoni, B. Anatomy, Head and Neck, Pulp (Tooth). In StatPearls; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2022. [Google Scholar]
- Mjör, I.A.; Sveen, O.B.; Heyeraas, K.J. Pulp-dentin biology in restorative dentistry. Part 1: Normal structure and physiology. Quintessence Int. 2001, 32, 427–446. [Google Scholar]
- Xie, Z.; Shen, Z.; Zhan, P.; Yang, J.; Huang, Q.; Huang, S.; Chen, L.; Lin, Z. Functional Dental Pulp Regeneration: Basic Research and Clinical Translation. Int. J. Mol. Sci. 2021, 22, 8991. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.T. Dental pulp and dentin tissue engineering and regeneration: Advancement and challenge. Front. Biosci. 2011, 3, 788–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, H.; Zhao, W.; Liu, A.; Wu, M.; Shuai, Y.; Li, B.; Huang, X.; Liu, X.; Yang, X.; Guo, X.; et al. SHED promote angiogenesis in stem cell-mediated dental pulp regeneration. Biochem. Biophys. Res. Commun. 2020, 529, 1158–1164. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, J.; Takahashi, N.; Matsumoto, T.; Yoshioka, Y.; Yamamoto, N.; Nishikawa, M.; Hibi, H.; Ishigro, N.; Ueda, M.; Furukawa, K.; et al. Factors secreted from dental pulp stem cells show multifaceted benefits for treating experimental rheumatoid arthritis. Bone 2016, 83, 210–219. [Google Scholar] [CrossRef]
- Liang, C.; Liao, L.; Tian, W. Stem Cell-based Dental Pulp Regeneration: Insights From Signaling Pathways. Stem Cell Rev. Rep. 2021, 17, 1251–1263. [Google Scholar] [CrossRef]
- Bai, Y.; Li, P.; Yin, G.; Huang, Z.; Liao, X.; Chen, X.; Yao, Y. BMP-2, VEGF and bFGF synergistically promote the osteogenic differentiation of rat bone marrow-derived mesenchymal stem cells. Biotechnol. Lett. 2013, 35, 301–308. [Google Scholar] [CrossRef]
- Khanna-Jain, R.; Mannerström, B.; Vuorinen, A.; Sándor, G.K.; Suuronen, R.; Miettinen, S. Osteogenic differentiation of human dental pulp stem cells on β-tricalcium phosphate/poly (l-lactic acid/caprolactone) three-dimensional scaffolds. J. Tissue Eng. 2012, 3, 2041731412467998. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Chen, E.; Chen, M.; Ye, C.; Qi, Y.; Ding, Q.; Li, H.; Xue, D.; Gao, X.; Pan, Z. IGFBP7 regulates the osteogenic differentiation of bone marrow-derived mesenchymal stem cells via Wnt/β-catenin signaling pathway. Faseb. J. 2018, 32, 2280–2291. [Google Scholar] [CrossRef] [Green Version]
- Omori, M.; Tsuchiya, S.; Hara, K.; Kuroda, K.; Hibi, H.; Okido, M.; Ueda, M. A new application of cell-free bone regeneration: Immobilizing stem cells from human exfoliated deciduous teeth-conditioned medium onto titanium implants using atmospheric pressure plasma treatment. Stem Cell Res. Ther. 2015, 6, 124. [Google Scholar] [CrossRef] [Green Version]
- Hilkens, P.; Driesen, R.B.; Wolfs, E.; Gervois, P.; Vangansewinkel, T.; Ratajczak, J.; Dillen, Y.; Bronckaers, A.; Lambrichts, I. Cryopreservation and Banking of Dental Stem Cells. Adv. Exp. Med. Biol. 2016, 951, 199–235. [Google Scholar] [CrossRef]
- Anitua, E.; Troya, M.; Zalduendo, M. Progress in the use of dental pulp stem cells in regenerative medicine. Cytotherapy 2018, 20, 479–498. [Google Scholar] [CrossRef]
- Gunawardena, T.N.A.; Masoudian, Z.; Rahman, M.T.; Ramasamy, T.S.; Ramanathan, A.; Abu Kasim, N.H. Dental derived stem cell conditioned media for hair growth stimulation. PLoS ONE 2019, 14, e0216003. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-S.; Lee, J.-H.; Mandakhbayar, N.; Jin, G.-Z.; Kim, S.-J.; Yoon, J.-Y.; Jo, S.B.; Park, J.-H.; Singh, R.K.; Jang, J.-H.; et al. Therapeutic tissue regenerative nanohybrids self-assembled from bioactive inorganic core/chitosan shell nanounits. Biomaterials 2021, 274, 120857. [Google Scholar] [CrossRef]
- Won, J.-E.; Lee, Y.S.; Park, J.-H.; Lee, J.-H.; Shin, Y.S.; Kim, C.-H.; Knowles, J.C.; Kim, H.-W. Hierarchical microchanneled scaffolds modulate multiple tissue-regenerative processes of immune-responses, angiogenesis, and stem cell homing. Biomaterials 2020, 227, 119548. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vu, H.T.; Han, M.-R.; Lee, J.-H.; Kim, J.-S.; Shin, J.-S.; Yoon, J.-Y.; Park, J.-H.; Dashnyam, K.; Knowles, J.C.; Lee, H.-H.; et al. Investigating the Effects of Conditioned Media from Stem Cells of Human Exfoliated Deciduous Teeth on Dental Pulp Stem Cells. Biomedicines 2022, 10, 906. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10040906
Vu HT, Han M-R, Lee J-H, Kim J-S, Shin J-S, Yoon J-Y, Park J-H, Dashnyam K, Knowles JC, Lee H-H, et al. Investigating the Effects of Conditioned Media from Stem Cells of Human Exfoliated Deciduous Teeth on Dental Pulp Stem Cells. Biomedicines. 2022; 10(4):906. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10040906
Chicago/Turabian StyleVu, Huong Thu, Mi-Ran Han, Jun-Haeng Lee, Jong-Soo Kim, Ji-Sun Shin, Ji-Young Yoon, Jeong-Hui Park, Khandmaa Dashnyam, Jonathan Campbell Knowles, Hae-Hyoung Lee, and et al. 2022. "Investigating the Effects of Conditioned Media from Stem Cells of Human Exfoliated Deciduous Teeth on Dental Pulp Stem Cells" Biomedicines 10, no. 4: 906. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10040906