
Sleep Disturbance Alters Cocaine-Induced Locomotor Activity: Involvement of Striatal Neuroimmune and Dopamine Signaling

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal and Reagents
2.2. SF Procedure and Cocaine Administration
2.3. Behavioral Experiments
2.4. Adult Microglia Isolation
2.5. RNA Extraction, Reverse Transcription, and Quantitative Polymerase Chain Reaction (qPCR)
2.6. Western Blots
2.7. Double Immunofluorescence Staining
2.8. Statistical Analysis
3. Results
3.1. Effects of SF on Cocaine-Mediated Behavioral Changes
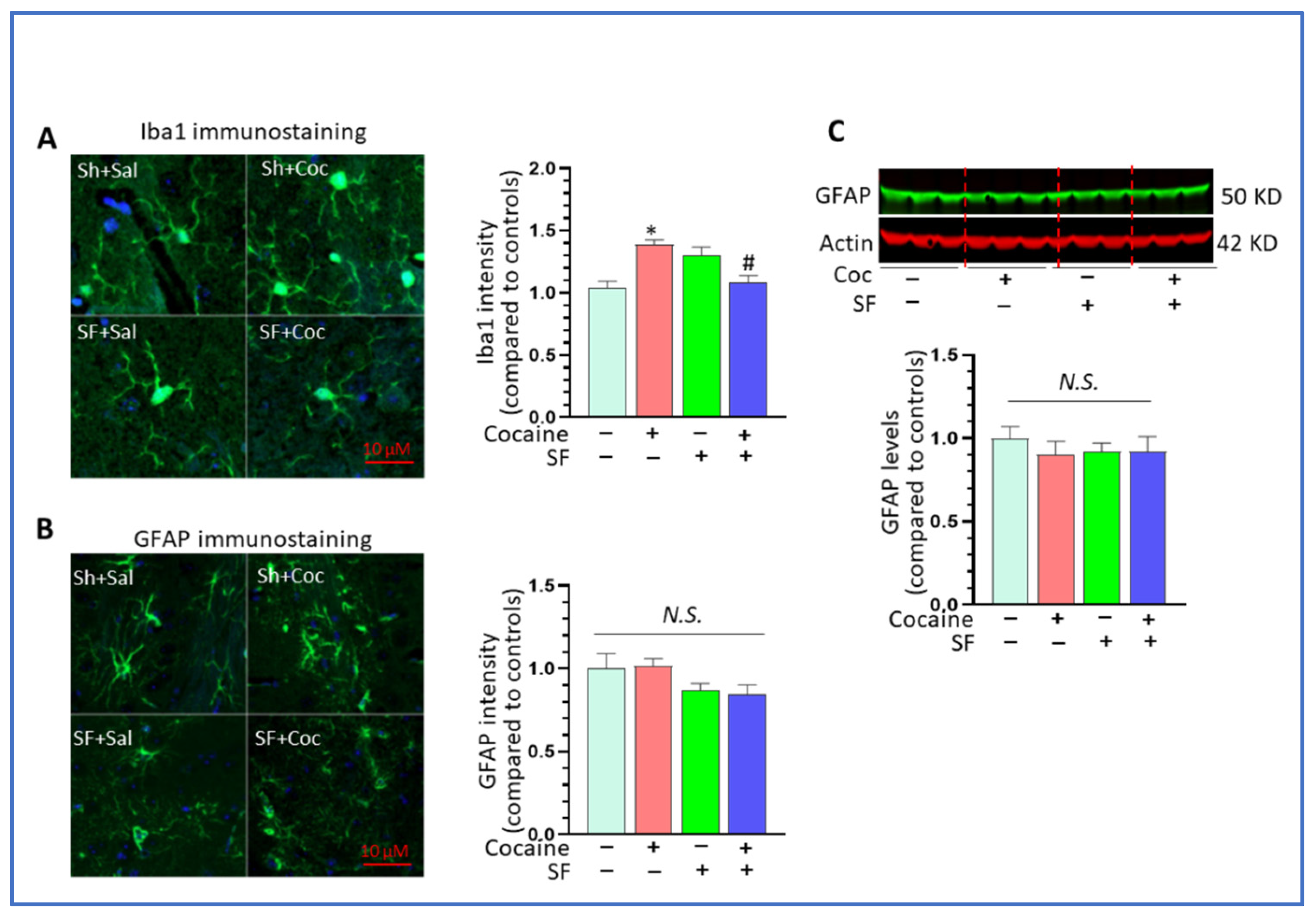
3.2. SF Blunted Cocaine-Mediated Neuroinflammation in the Striatum
3.3. The Combined Effects of SF and Cocaine on Microglial Immune Responses
3.4. Combined Effects of SF and Cocaine on Astrocytes
3.5. Effects of Cocaine and SF on NLRP3 Inflammasome and Dopamine Signaling In Vivo
3.6. Effects of SF on Cocaine-Mediated Locomotion Sensitivity and Neuroimmune Signaling in Withdrawn Mice
3.7. Effects of SF and Cocaine on Microglial Activation and Dopamine System in Withdrawn Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buysse, D.J. Sleep Health: Can We Define It? Does It Matter? Sleep 2014, 37, 9–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, C.C.; Vincent, G.E.; Coates, A.M.; Khalesi, S.; Irwin, C.; Dorrian, J.; Ferguson, S.A. A Time to Rest, a Time to Dine: Sleep, Time-Restricted Eating, and Cardiometabolic Health. Nutrients 2022, 14, 420. [Google Scholar] [CrossRef] [PubMed]
- Grandner, M.A.; Fernandez, F.-X. The translational neuroscience of sleep: A contextual framework. Science 2021, 374, 568–573. [Google Scholar] [CrossRef] [PubMed]
- Ohayon, M.M. Epidemiology of insomnia: What we know and what we still need to learn. Sleep Med. Rev. 2002, 6, 97–111. [Google Scholar] [CrossRef] [PubMed]
- Bjorness, T.E.; Greene, R.W. Interaction between cocaine use and sleep behavior: A comprehensive review of cocaine’s disrupting influence on sleep behavior and sleep disruptions influence on reward seeking. Pharmacol. Biochem. Behav. 2021, 206, 173194. [Google Scholar] [CrossRef] [PubMed]
- Koob, G.F.; Colrain, I.M. Alcohol use disorder and sleep disturbances: A feed-forward allostatic framework. Neuro Psychopharmacol. 2020, 45, 141–165. [Google Scholar] [CrossRef] [PubMed]
- Angarita, G.A.; Emadi, N.; Hodges, S.; Morgan, P.T. Sleep abnormalities associated with alcohol, cannabis, cocaine, and opiate use: A comprehensive review. Addict. Sci. Clin. Pract. 2016, 11, 9. [Google Scholar] [CrossRef] [Green Version]
- Ahrens, A.M.; Ahmed, O.J. Neural circuits linking sleep and addiction: Animal models to understand why select individuals are more vulnerable to substance use disorders after sleep deprivation. Neurosci. Biobehav. Rev. 2020, 108, 435–444. [Google Scholar] [CrossRef]
- Logan, R.W.; Hasler, B.P.; Forbes, E.E.; Franzen, P.L.; Torregrossa, M.M.; Huang, Y.H.; Buysse, D.J.; Clark, D.B.; McClung, C.A. Impact of Sleep and Circadian Rhythms on Addiction Vulnerability in Adolescents. Biol. Psychiatry 2018, 83, 987–996. [Google Scholar] [CrossRef]
- Namba, M.D.; Leyrer-Jackson, J.M.; Nagy, E.K.; Olive, M.F.; Neisewander, J.L. Neuroimmune Mechanisms as Novel Treatment Targets for Substance Use Disorders and Associated Comorbidities. Front. Neurosci. 2021, 15, 650785. [Google Scholar] [CrossRef]
- Berríos-Cárcamo, P.; Quezada, M.; Quintanilla, M.E.; Morales, P.; Ezquer, M.; Herrera-Marschitz, M.; Israel, Y.; Ezquer, F. Oxidative Stress and Neuroinflammation as a Pivot in Drug Abuse. A Focus on the Therapeutic Potential of Antioxidant and Anti-inflammatory Agents and Biomolecules. Antioxidants 2020, 9, 830. [Google Scholar] [CrossRef] [PubMed]
- Liao, K.; Guo, M.; Niu, F.; Yang, L.; Callen, S.E.; Buch, S. Cocaine-mediated induction of microglial activation involves the ER stress-TLR2 axis. J. Neuroinflamm. 2016, 13, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, M.-L.; Liao, K.; Periyasamy, P.; Yang, L.; Cai, Y.; Callen, S.E.; Buch, S. Cocaine-mediated microglial activation involves the ER stress-autophagy axis. Autophagy 2015, 11, 995–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, K.H.; Wiley, C.A.; Bradberry, C.W. Psychostimulant Abuse and Neuroinflammation: Emerging Evidence of Their Interconnection. Neurotox. Res. 2013, 23, 174–188. [Google Scholar] [CrossRef]
- Cui, C.; Shurtleff, D.; Harris, R.A. Neuroimmune Mechanisms of Alcohol and Drug Addiction. Int. Rev. Neurobiol. 2014, 118, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.H.; Lutjens, R.; van der Stap, L.D.; Lekic, D.; Romano-Spica, V.; Morales, M.; Koob, G.F.; Repunte-Canonigo, V.; Sanna, P.P. Gene expression evidence for remodeling of lateral hypothalamic circuitry in cocaine addiction. Proc. Natl. Acad. Sci. USA 2005, 102, 11533–11538. [Google Scholar] [CrossRef] [Green Version]
- Piechota, M.; Korostynski, M.; Solecki, W.; Gieryk, A.; Slezak, M.; Bilecki, W.; Ziolkowska, B.; Kostrzewa, E.; Cymerman, I.; Swiech, L.; et al. The dissection of transcriptional modules regulated by various drugs of abuse in the mouse striatum. Genome Biol. 2010, 11, R48. [Google Scholar] [CrossRef] [Green Version]
- Vallender, E.J.; Goswami, D.B.; Shinday, N.M.; Westmoreland, S.V.; Yao, W.-D.; Rowlett, J.K. Transcriptomic profiling of the ventral tegmental area and nucleus accumbens in rhesus macaques following long-term cocaine self-administration. Drug Alcohol Depend. 2017, 175, 9–23. [Google Scholar] [CrossRef]
- Northcutt, A.; Hutchinson, M.; Wang, X.; Baratta, M.; Hiranita, T.; Cochran, T.; Pomrenze, M.B.; Galer, E.; Kopajtic, T.A.; Li, C.; et al. DAT isn’t all that: Cocaine reward and reinforcement require Toll-like receptor 4 signaling. Mol. Psychiatry 2015, 20, 1525–1537. [Google Scholar] [CrossRef] [Green Version]
- Little, K.Y.; Ramssen, E.; Welchko, R.; Volberg, V.; Roland, C.J.; Cassin, B. Decreased brain dopamine cell numbers in human cocaine users. Psychiatry Res. 2009, 168, 173–180. [Google Scholar] [CrossRef]
- Kohno, M.; Link, J.; Dennis, L.E.; McCready, H.; Huckans, M.; Hoffman, W.F.; Loftis, J.M. Neuroinflammation in addiction: A review of neuroimaging studies and potential immunotherapies. Pharmacol. Biochem. Behav. 2019, 179, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Kashima, D.T.; Grueter, B.A. Toll-like receptor 4 deficiency alters nucleus accumbens synaptic physiology and drug reward behavior. Proc. Natl. Acad. Sci. USA 2017, 114, 8865–8870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, R.; Bu, Q.; Fu, D.; Shao, X.; Jiang, L.; Guo, W.; Chen, B.; Liu, B.; Hu, Z.; Tian, J.; et al. Toll-like receptor 3 modulates the behavioral effects of cocaine in mice. J. Neuro Inflamm. 2018, 15, 93. [Google Scholar] [CrossRef] [PubMed]
- Catale, C.; Bussone, S.; Iacono, L.L.; Carola, V. Microglial alterations induced by psychoactive drugs: A possible mechanism in substance use disorder? Semin. Cell Dev. Biol. 2019, 94, 164–175. [Google Scholar] [CrossRef]
- Bachtell, R.K.; Jones, J.D.; Heinzerling, K.G.; Beardsley, P.M.; Comer, S.D. Glial and neuroinflammatory targets for treating substance use disorders. Drug Alcohol Depend. 2017, 180, 156–170. [Google Scholar] [CrossRef]
- Ngarka, L.; Fodjo, J.N.S.; Aly, E.; Masocha, W.; Njamnshi, A.K. The Interplay between Neuroinfections, the Immune System and Neurological Disorders: A Focus on Africa. Front. Immunol. 2021, 12, 803475. [Google Scholar] [CrossRef]
- Dauvilliers, Y. Hypocretin/Orexin, Sleep and Alzheimer’s Disease. Front. Neurol. Neurosci. 2021, 45, 139–149. [Google Scholar]
- Wadhwa, M.; Kumari, P.; Chauhan, G.; Roy, K.; Alam, S.; Kishore, K.; Ray, K.; Panjwani, U. Sleep deprivation induces spatial memory impairment by altered hippocampus neuroinflammatory responses and glial cells activation in rats. J. Neuro Immunol. 2017, 312, 38–48. [Google Scholar] [CrossRef]
- Wadhwa, M.; Prabhakar, A.; Ray, K.; Roy, K.; Kumari, P.; Jha, P.K.; Kishore, K.; Kumar, S.; Panjwani, U. Inhibiting the microglia activation improves the spatial memory and adult neurogenesis in rat hippocampus during 48 h of sleep deprivation. J. Neuro Inflamm. 2017, 14, 222. [Google Scholar] [CrossRef] [Green Version]
- Hurtado-Alvarado, G.; Becerril-Villanueva, E.; de Oca, A.C.-M.; Domínguez-Salazar, E.; Salinas-Jazmín, N.; Pérez-Tapia, S.; Pavon, L.; Velázquez-Moctezuma, J.; Gómez-González, B. The yin/yang of inflammatory status: Blood-brain barrier regulation during sleep. Brain Behav. Immun. 2018, 69, 154–166. [Google Scholar] [CrossRef]
- Kincheski, G.C.; Valentim, I.S.; Clarke, J.R.; Cozachenco, D.; Castelo-Branco, M.T.; Ramos-Lobo, A.M.; Rumjanek, V.M.; Donato, J., Jr.; De Felice, F.G.; Ferreira, S.T. Chronic sleep restriction promotes brain inflammation and synapse loss, and potentiates memory impairment induced by amyloid-beta oligomers in mice. Brain Behav. Immun. 2017, 64, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Manchanda, S.; Singh, H.; Kaur, T.; Kaur, G. Low-grade neuroinflammation due to chronic sleep deprivation results in anxiety and learning and memory impairments. Mol. Cell. Biochem. 2018, 449, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; Kunitachi, S.; Iyo, M.; Hashimoto, K. The antibiotic minocycline prevents methamphetamine-induced rewarding effects in mice. Pharmacol. Biochem. Behav. 2012, 101, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Manev, H. Effects of minocycline on cocaine sensitization and phosphorylation of GluR1 receptors in 5-lipoxygenase deficient mice. Neuro Pharmacol. 2011, 60, 1058–1063. [Google Scholar] [CrossRef] [Green Version]
- Besedovsky, L.; Schmidt, E.-M.; Linz, B.; Diekelmann, S.; Lange, T.; Born, J. Signs of enhanced sleep and sleep-associated memory processing following the anti-inflammatory antibiotic minocycline in men. J. Psycho Pharmacol. 2017, 31, 204–210. [Google Scholar] [CrossRef]
- Bhagavan, S.M.; Sahota, P.K. Sleep Fragmentation and Atherosclerosis: Is There a Relationship? Mo. Med. 2021, 118, 272–276. [Google Scholar]
- Hakim, F.; Wang, Y.; Zhang, S.X.; Zheng, J.; Yolcu, E.S.; Carreras, A.; Khalyfa, A.; Shirwan, H.; Almendros, I.; Gozal, D. Fragmented Sleep Accelerates Tumor Growth and Progression through Recruitment of Tumor-Associated Macrophages and TLR4 Signaling. Cancer Res. 2014, 74, 1329–1337. [Google Scholar] [CrossRef] [Green Version]
- Nair, D.; Zhang, S.X.L.; Ramesh, V.; Hakim, F.; Kaushal, N.; Wang, Y.; Gozal, D. Sleep Fragmentation Induces Cognitive Deficits Via Nicotinamide Adenine Dinucleotide Phosphate Oxidase–Dependent Pathways in Mouse. Am. J. Respir. Crit. Care Med. 2011, 184, 1305–1312. [Google Scholar] [CrossRef]
- Ramesh, V.; Nair, D.; Zhang, S.X.L.; Hakim, F.; Kaushal, N.; Kayali, F.; Wang, Y.; Li, R.C.; Carreras, A.; Gozal, D. Disrupted sleep without sleep curtailment induces sleepiness and cognitive dysfunction via the tumor necrosis factor-α pathway. J. Neuro Inflamm. 2012, 9, 91. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.N.; Penrod, R.D.; Taniguchi, M.; Cowan, C.W. Assessment of Cocaine-induced Behavioral Sensitization and Conditioned Place Preference in Mice. J. Vis. Exp. 2016, 108, e53107. [Google Scholar] [CrossRef] [Green Version]
- Zombeck, J.A.; Swearingen, S.P.; Rhodes, J.S. Acute locomotor responses to cocaine in adolescents vs. adults from four divergent inbred mouse strains. Genes Brain Behav. 2010, 9, 892–898. [Google Scholar] [CrossRef] [PubMed]
- Berro, L.; Santos, R.; Hollais, A.; Wuo-Silva, R.; Fukushiro, D.; Mári-Kawamoto, E.; Costa, J.; Trombin, T.; Patti, C.; Grapiglia, S.; et al. Acute total sleep deprivation potentiates cocaine-induced hyperlocomotion in mice. Neurosci. Lett. 2014, 579, 130–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosário, B.; Nazaré, M.D.F.S.D.; Estadella, D.; Ribeiro, D.A.; Viana, M.D.B. Behavioral and neurobiological alterations induced by chronic use of crack cocaine. Rev. Neurosci. 2019, 31, 59–75. [Google Scholar] [CrossRef] [PubMed]
- Naranjo, C.; Del Reguero, L.; Moratalla, G.; Hercberg, M.; Valenzuela, M.; Failde, I. Anxiety, depression and sleep disorders in patients with diabetic neuropathic pain: A systematic review. Expert Rev. Neurother. 2019, 19, 1201–1209. [Google Scholar] [CrossRef]
- Zilkha, N.; Barnea-Ygael, N.; Keidar, L.; Zangen, A. Increased relapse to cocaine-seeking in a genetic model for depression. Addict. Biol. 2020, 25, e12756. [Google Scholar] [CrossRef]
- Strekalova, T.; Liu, Y.; Kiselev, D.; Khairuddin, S.; Chiu, J.L.Y.; Lam, J.; Chan, Y.-S.; Pavlov, D.; Proshin, A.; Lesch, K.-P.; et al. Chronic mild stress paradigm as a rat model of depression: Facts, artifacts, and future perspectives. Psychopharmacology 2022, 239, 663–693. [Google Scholar] [CrossRef]
- Pretre, V.; Papadopoulos, D.; Regard, J.; Pelletier, M.; Woo, J. Interleukin-1 (IL-1) and the inflammasome in cancer. Cytokine 2022, 153, 155850. [Google Scholar] [CrossRef]
- Schroder, K.; Tschopp, J. The inflammasomes. Cell 2010, 140, 821–832. [Google Scholar] [CrossRef] [Green Version]
- Kohno, M.; Dennis, L.E.; McCready, H.; Hoffman, W.F. Dopamine dysfunction in stimulant use disorders: Mechanistic comparisons and implications for treatment. Mol. Psychiatry 2021, 27, 220–229. [Google Scholar] [CrossRef]
- Dauvilliers, Y.; Tafti, M.; Landolt, H.P. Catechol-O-methyltransferase, dopamine, and sleep-wake regulation. Sleep Med. Rev. 2015, 22, 47–53. [Google Scholar] [CrossRef]
- Nestler, E.J. Historical review: Molecular and cellular mechanisms of opiate and cocaine addiction. Trends Pharmacol. Sci. 2004, 25, 210–218. [Google Scholar] [CrossRef]
- Coley, A.A.; Gao, W.J. PSD95: A synaptic protein implicated in schizophrenia or autism? Prog. Neuro Psychopharmacol. Biol. Psychiatry 2018, 82, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Peart, D.R.; Andrade, A.K.; Logan, C.N.; Knackstedt, L.A.; Murray, J.E. Regulation of cocaine-related behaviours by estrogen and progesterone. Neurosci. Biobehav. Rev. 2022, 135, 104584. [Google Scholar] [CrossRef] [PubMed]
- Bjorness, T.E.; Greene, R.W. Sleep deprivation alters the time course but not magnitude of locomotor sensitization to cocaine. Sci. Rep. 2018, 8, 17672. [Google Scholar] [CrossRef] [PubMed]
- Bjorness, T.E.; Greene, R.W. Sleep Deprivation Enhances Cocaine Conditioned Place Preference in an Orexin Receptor-Modulated Manner. eNeuro 2020, 7. [Google Scholar] [CrossRef]
- Berro, L.; Hollais, A.; Patti, C.; Fukushiro, D.; Mári-Kawamoto, E.; Talhati, F.; Costa, J.; Zanin, K.; Lopes-Silva, L.; Ceccon, L.; et al. Sleep deprivation impairs the extinction of cocaine-induced environmental conditioning in mice. Pharmacol. Biochem. Behav. 2014, 124, 13–18. [Google Scholar] [CrossRef]
- Chen, B.; Wang, Y.; Liu, X.; Liu, Z.; Dong, Y.; Huang, Y.H. Sleep Regulates Incubation of Cocaine Craving. J. Neurosci. 2015, 35, 13300–13310. [Google Scholar] [CrossRef] [Green Version]
- Saito, L.P.; Fukushiro, D.F.; Hollais, A.; Mári-Kawamoto, E.; Costa, J.M.; Berro, L.F.; Aramini, T.C.; Wuo-Silva, R.; Andersen, M.L.; Tufik, S.; et al. Acute total sleep deprivation potentiates amphetamine-induced locomotor-stimulant effects and behavioral sensitization in mice. Pharmacol. Biochem. Behav. 2014, 117, 7–16. [Google Scholar] [CrossRef]
- Kameda, S.R.; Fukushiro, D.F.; Trombin, T.F.; Sanday, L.; Wuo-Silva, R.; Saito, L.P.; Tufik, S.; D’Almeida, V.; Frussa-Filho, R. The effects of paradoxical sleep deprivation on amphetamine-induced behavioral sensitization in adult and adolescent mice. Psychiatry Res. 2014, 218, 335–340. [Google Scholar] [CrossRef]
- Shahveisi, K.; Abdoli, N.; Farnia, V.; Khazaie, H.; Hosseini, M.; Ghazvini, H.; Khodamoradi, M. REM sleep deprivation before extinction or reinstatement alters methamphetamine reward memory via D1-like dopamine receptors. Pharmacol. Biochem. Behav. 2022, 213, 173319. [Google Scholar] [CrossRef]
- Ahmadi-Soleimani, S.M.; Azizi, H.; Abbasi-Mazar, A. Intermittent REM sleep deprivation attenuates the development of morphine tolerance and dependence in male rats. Neurosci. Lett. 2021, 748, 135735. [Google Scholar] [CrossRef] [PubMed]
- Eacret, D.; Lemchi, C.; Caulfield, J.I.; Cavigelli, S.A.; Veasey, S.C.; Blendy, J.A. Chronic Sleep Deprivation Blocks Voluntary Morphine Consumption but Not Conditioned Place Preference in Mice. Front. Neurosci. 2022, 16, 836693. [Google Scholar] [CrossRef] [PubMed]
- García-García, F.; Priego-Fernández, S.; López-Muciño, L.A.; Acosta-Hernández, M.E.; Peña-Escudero, C. Increased alcohol consumption in sleep-restricted rats is mediated by delta FosB induction. Alcohol 2021, 93, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Araujo, N.P.; Andersen, M.L.; Abilio, V.; Gomes, D.C.; Carvalho, R.C.; Silva, R.; Ribeiro, R.D.A.; Tufik, S.; Frussa-Filho, R. Sleep deprivation abolishes the locomotor stimulant effect of ethanol in mice. Brain Res. Bull. 2006, 69, 332–337. [Google Scholar] [CrossRef]
- Reeves-Darby, J.A.; Berro, L.F.; Rowlett, J.K.; Platt, D.M. Enhancement of cue-induced reinstatement of alcohol seeking by acute total sleep restriction in male Wistar rats. Pharmacol. Biochem. Behav. 2021, 205, 173188. [Google Scholar] [CrossRef]
- Almendros, I.; Basoglu, Ö.K.; Conde, S.V.; Liguori, C.; Saaresranta, T. Metabolic dysfunction in OSA: Is there something new under the sun? J. Sleep Res. 2022, 31, e13418. [Google Scholar] [CrossRef]
- Yeghiazarians, Y.; Jneid, H.; Tietjens, J.R.; Redline, S.; Brown, D.L.; El-Sherif, N.; Mehra, R.; Bozkurt, B.; Ndumele, C.E.; Somers, V.K. Obstructive Sleep Apnea and Cardiovascular Disease: A Scientific Statement from the American Heart Association. Circulation 2021, 144, e56–e67. [Google Scholar] [CrossRef]
- Cheng, Y.; Kim, W.-K.; Wellman, L.L.; Sanford, L.D.; Guo, M.-L. Short-Term Sleep Fragmentation Dysregulates Autophagy in a Brain Region-Specific Manner. Life 2021, 11, 1098. [Google Scholar] [CrossRef]
- Zernig, G.; Ahmed, S.; Cardinal, R.; Morgan, D.; Acquas, E.; Foltin, R.W.; Vezina, P.; Negus, S.S.; Crespo, J.A.; Stöckl, P.; et al. Explaining the Escalation of Drug Use in Substance Dependence: Models and Appropriate Animal Laboratory Tests. Pharmacology 2007, 80, 65–119. [Google Scholar] [CrossRef]
- Volkow, N.D.; Tomasi, D.; Wang, G.-J.; Telang, F.; Fowler, J.S.; Logan, J.; Benveniste, H.; Kim, R.; Thanos, P.K.; Ferre, S. Evidence That Sleep Deprivation Downregulates Dopamine D2R in Ventral Striatum in the Human Brain. J. Neurosci. 2012, 32, 6711–6717. [Google Scholar] [CrossRef] [Green Version]
- Wiers, C.E.; Shumay, E.; Cabrera, E.; Shokri-Kojori, E.; Gladwin, T.E.; Skarda, E.; Cunningham, S.I.; Kim, S.W.; Wong, T.C.; Tomasi, D.; et al. Reduced sleep duration mediates decreases in striatal D2/D3 receptor availability in cocaine abusers. Transl. Psychiatry 2016, 6, e752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasler, B.P.; Soehner, A.M.; Clark, D.B. Sleep and circadian contributions to adolescent alcohol use disorder. Alcohol 2015, 49, 377–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tavernier, R.; Munroe, M.; Willoughby, T. Perceived morningness-eveningness predicts academic adjustment and substance use across university, but social jetlag is not to blame. Chronobiol. Int. 2015, 32, 1233–1245. [Google Scholar] [CrossRef] [PubMed]
- Depoy, L.M.; McClung, C.A.; Logan, R.W. Neural Mechanisms of Circadian Regulation of Natural and Drug Reward. Neural Plast. 2017, 2017, 5720842. [Google Scholar] [CrossRef] [Green Version]
- Periyasamy, P.; Guo, M.-L.; Buch, S. Cocaine induces astrocytosis through ER stress-mediated activation of autophagy. Autophagy 2016, 12, 1310–1329. [Google Scholar] [CrossRef] [Green Version]
- Bachtell, R.; Hutchinson, M.; Wang, X.; Rice, K.C.; Maier, S.F.; Watkins, L.R. Targeting the Toll of Drug Abuse: The Translational Potential of Toll-Like Receptor 4. CNS Neurol. Disord. Drug Targets 2015, 14, 692–699. [Google Scholar] [CrossRef]
- McCarthy, G.M.; Farris, S.P.; Blednov, Y.A.; Harris, R.A.; Mayfield, R.D. Microglial-specific transcriptome changes following chronic alcohol consumption. Neuro Pharmacol. 2018, 128, 416–424. [Google Scholar] [CrossRef]
- Belin, D.; Deroche-Gamonet, V.; Jaber, M. Cocaine-induced sensitization is associated with altered dynamics of transcriptional responses of the dopamine transporter, tyrosine hydroxylase, and dopamine D2 receptors in C57Bl/6J mice. Psycho Pharmacol. 2007, 193, 567–578. [Google Scholar] [CrossRef]
- Qiu, C.; Wang, M.; Yu, W.; Rong, Z.; Zheng, H.S.; Sun, T.; Liu, S.B.; Zhao, M.G.; Wu, Y.M. Activation of the Hippocampal LXRbeta Improves Sleep-Deprived Cognitive Impairment by Inhibiting Neuroinflammation. Mol. Neurobiol. 2021, 58, 5272–5288. [Google Scholar] [CrossRef]
- Bertrand, S.J.; Zhang, Z.; Patel, R.; O’Ferrell, C.; Punjabi, N.M.; Kudchadkar, S.R.; Kannan, S. Transient neonatal sleep fragmentation results in long-term neuroinflammation and cognitive impairment in a rabbit model. Exp. Neurol. 2020, 327, 113212. [Google Scholar] [CrossRef]
- Tsai, S.-J. Role of interleukin 8 in depression and other psychiatric disorders. Prog. Neuro-Psycho Pharmacol. Biol. Psychiatry 2021, 106, 110173. [Google Scholar] [CrossRef] [PubMed]
- Kimura, L.F.; Novaes, L.S.; Picolo, G.; Munhoz, C.D.; Cheung, C.W.; Camarini, R. How environmental enrichment balances out neuroinflammation in chronic pain and comorbid depression and anxiety disorders. Br. J. Pharmacol. 2022, 179, 1640–1660. [Google Scholar] [CrossRef]
- Arenas, D.J.; Thomas, A.; Wang, J.; DeLisser, H.M. A Systematic Review and Meta-analysis of Depression, Anxiety, and Sleep Disorders in US Adults with Food Insecurity. J. Gen. Intern. Med. 2019, 34, 2874–2882. [Google Scholar] [CrossRef] [PubMed]
- Motomura, Y.; Katsunuma, R.; Ayabe, N.; Oba, K.; Terasawa, Y.; Kitamura, S.; Moriguchi, Y.; Hida, A.; Kamei, Y.; Mishima, K. Decreased activity in the reward network of chronic insomnia patients. Sci. Rep. 2021, 11, 3600. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roodsari, S.K.; Cheng, Y.; Reed, K.M.; Wellman, L.L.; Sanford, L.D.; Kim, W.-K.; Guo, M.-L. Sleep Disturbance Alters Cocaine-Induced Locomotor Activity: Involvement of Striatal Neuroimmune and Dopamine Signaling. Biomedicines 2022, 10, 1161. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10051161
Roodsari SK, Cheng Y, Reed KM, Wellman LL, Sanford LD, Kim W-K, Guo M-L. Sleep Disturbance Alters Cocaine-Induced Locomotor Activity: Involvement of Striatal Neuroimmune and Dopamine Signaling. Biomedicines. 2022; 10(5):1161. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10051161
Chicago/Turabian StyleRoodsari, Soheil Kazemi, Yan Cheng, Kirstin M. Reed, Laurie L. Wellman, Larry D. Sanford, Woong-Ki Kim, and Ming-Lei Guo. 2022. "Sleep Disturbance Alters Cocaine-Induced Locomotor Activity: Involvement of Striatal Neuroimmune and Dopamine Signaling" Biomedicines 10, no. 5: 1161. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10051161