Antigen Load and T Cell Function: A Challenging Interaction in HBV Infection

 and
and
Abstract
:1. Introduction
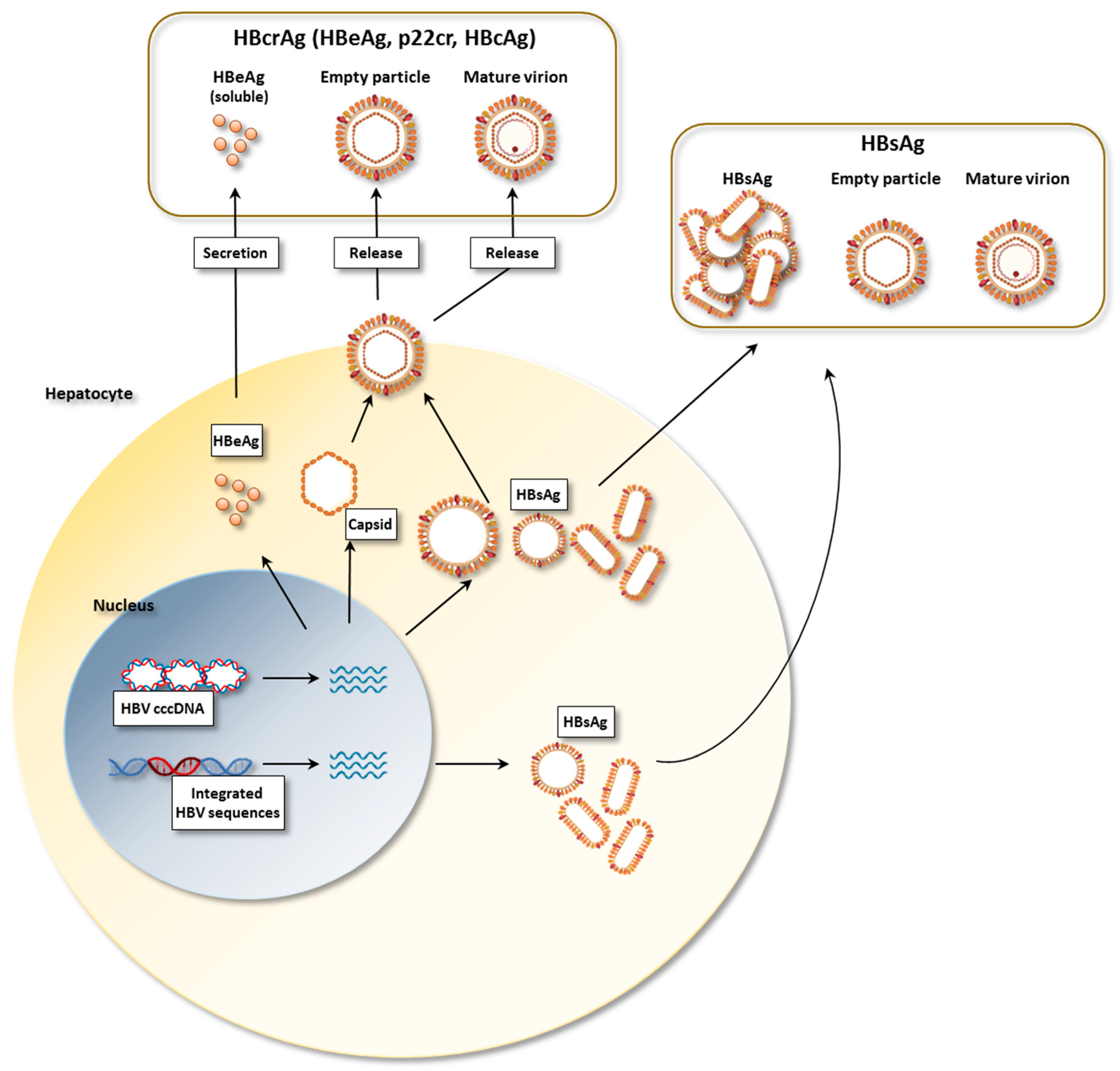
2. HBV Proteins
3. The Role of Antigen Expression in Animal Models
4. Human Studies
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- McLane, L.M.; Abdel-Hakeem, M.S.; Wherry, E.J. CD8 T cell exhaustion during chronic viral infection and cancer. Annu. Rev. Immunol. 2019, 37, 457–495. [Google Scholar] [CrossRef] [Green Version]
- Fisicaro, P.; Barili, V.; Rossi, M.; Montali, I.; Vecchi, A.; Acerbi, G.; Laccabue, D.; Zecca, A.; Penna, A.; Missale, G.; et al. Pathogenetic mechanisms of T cell dysfunction in chronic HBV infection and related therapeutic approaches. Front. Immunol. 2020, 11, 849. [Google Scholar] [CrossRef]
- Rydell, G.E.; Prakash, K.; Norder, H.; Lindh, M. Hepatitis B surface antigen on subviral particles reduces the neutralizing effect of anti-HBs antibodies on hepatitis B viral particles in vitro. Virology 2017, 509, 67–70. [Google Scholar] [CrossRef]
- Iannacone, M.; Guidotti, L.G. Immunobiology and pathogenesis of hepatitis B virus infection. Nat. Rev. Immunol. 2022, 22, 19–32. [Google Scholar] [CrossRef]
- Fanning, G.C.; Zoulim, F.; Hou, J.; Bertoletti, A. Author correction: Therapeutic STRATEGIES for hepatitis B virus infection: Towards a cure. Nat. Rev. Drug Discov. 2020, 19, 291. [Google Scholar] [CrossRef] [Green Version]
- Slagle, B.L.; Bouchard, M.J. Hepatitis B virus X and regulation of viral gene expression. Cold Spring Harb. Perspect. Med. 2016, 6, a021402. [Google Scholar] [CrossRef] [Green Version]
- Bill, C.A.; Summers, J. Genomic DNA double-strand breaks are targets for hepadnaviral DNA integration. Proc. Natl. Acad. Sci. USA 2004, 101, 11135–11140. [Google Scholar] [CrossRef] [Green Version]
- Wooddell, C.I.; Yuen, M.-F.; Chan, H.L.-Y.; Gish, R.G.; Locarnini, S.A.; Chavez, D.; Ferrari, C.; Given, B.D.; Hamilton, J.; Kanner, S.B.; et al. RNAi-based treatment of chronically infected patients and chimpanzees reveals that integrated hepatitis B virus DNA is a source of HBsAg. Sci. Transl. Med. 2017, 9, eaan0241. [Google Scholar] [CrossRef] [Green Version]
- Meier, M.-A.; Calabrese, D.; Suslov, A.; Terracciano, L.M.; Heim, M.H.; Wieland, S. Ubiquitous expression of HBsAg from integrated HBV DNA in patients with low viral load. J. Hepatol. 2021, 75, 840–847. [Google Scholar] [CrossRef]
- Rydell, G.E.; Larsson, S.B.; Prakash, K.; Andersson, M.; Norder, H.; Hellstrand, K.; Norkrans, G.; Lindh, M. Abundance of noncircular intrahepatic hepatitis B virus DNA may reflect frequent integration into human DNA in chronically infected patients. J. Infect. Dis. 2020, jiaa572. [Google Scholar] [CrossRef]
- Mason, W.S.; Gill, U.S.; Litwin, S.; Zhou, Y.; Peri, S.; Pop, O.; Hong, M.L.W.; Naik, S.; Quaglia, A.; Bertoletti, A.; et al. HBV DNA integration and clonal hepatocyte expansion in chronic hepatitis B patients considered immune tolerant. Gastroenterology 2016, 151, 986–998.e4. [Google Scholar] [CrossRef] [Green Version]
- Cornberg, M.; Wong, V.W.-S.; Locarnini, S.; Brunetto, M.; Janssen, H.L.A.; Chan, H.L.-Y. The role of quantitative hepatitis B surface antigen revisited. J. Hepatol. 2017, 66, 398–411. [Google Scholar] [CrossRef] [Green Version]
- Tsai, K.-N.; Ou, J.-H.J. Hepatitis B virus e antigen and viral persistence. Curr. Opin. Virol. 2021, 51, 158–163. [Google Scholar] [CrossRef]
- Kimura, T.; Ohno, N.; Terada, N.; Rokuhara, A.; Matsumoto, A.; Yagi, S.; Tanaka, E.; Kiyosawa, K.; Ohno, S.; Maki, N. Hepatitis B virus DNA-negative dane particles lack core protein but contain a 22-kDa precore protein without C-terminal arginine-rich domain. J. Biol. Chem. 2005, 280, 21713–21719. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.T.; Billaud, J.-N.; Sällberg, M.; Guidotti, L.G.; Chisari, F.V.; Jones, J.; Hughes, J.; Milich, D.R. A function of the hepatitis B virus precore protein is to regulate the immune response to the core antigen. Proc. Natl. Acad. Sci. USA 2004, 101, 14913–14918. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Sällberg, M.; Hughes, J.; Jones, J.; Guidotti, L.G.; Chisari, F.V.; Billaud, J.-N.; Milich, D.R. Immune tolerance split between hepatitis B virus precore and core proteins. J. Virol. 2005, 79, 3016–3027. [Google Scholar] [CrossRef] [Green Version]
- Mak, L.-Y.; Wong, D.K.-H.; Cheung, K.-S.; Seto, W.-K.; Lai, C.-L.; Yuen, M.-F. Review article: Hepatitis B core-related antigen (HBcrAg): An emerging marker for chronic hepatitis B virus infection. Aliment. Pharmacol. Ther. 2018, 47, 43–54. [Google Scholar] [CrossRef]
- Wong, D.K.-H.; Seto, W.-K.; Cheung, K.-S.; Chong, C.-K.; Huang, F.-Y.; Fung, J.; Lai, C.-L.; Yuen, M.-F. Hepatitis B virus core-related antigen as a surrogate marker for covalently closed circular DNA. Liver Int. 2017, 37, 995–1001. [Google Scholar] [CrossRef]
- Testoni, B.; Lebossé, F.; Scholtes, C.; Berby, F.; Miaglia, C.; Subic, M.; Loglio, A.; Facchetti, F.; Lampertico, P.; Levrero, M.; et al. Serum hepatitis B core-related antigen (HBcrAg) correlates with covalently closed circular DNA transcriptional activity in chronic hepatitis B patients. J. Hepatol. 2019, 70, 615–625. [Google Scholar] [CrossRef]
- Chen, S.; Jia, J.; Gao, Y.; Li, H.; Fang, M.; Feng, H.; Guan, W.; Ji, J.; Gao, Z.; Gao, C. Clinical evaluation of hepatitis B core-related antigen in chronic hepatitis B and hepatocellular carcinoma patients. Clin. Chim. Acta 2018, 486, 237–244. [Google Scholar] [CrossRef]
- Erken, R.; Zaaijer, H.L.; Willemse, S.B.; Bakker, E.; Takkenberg, B.B.; Reesink, H.W.; Kootstra, N.A. Hepatitis B core related antigen in relation to intrahepatic and circulating viral markers, before and after combination therapy. Ann. Hepatol. 2021, 26, 100540. [Google Scholar] [CrossRef]
- Watanabe, T.; Inoue, T.; Tanaka, Y. Hepatitis B core-related antigen and new therapies for hepatitis, B. Microorganisms 2021, 9, 2083. [Google Scholar] [CrossRef]
- Inoue, T.; Kusumoto, S.; Iio, E.; Ogawa, S.; Suzuki, T.; Yagi, S.; Kaneko, A.; Matsuura, K.; Aoyagi, K.; Tanaka, Y. Clinical efficacy of a novel, high-sensitivity HBcrAg assay in the management of chronic hepatitis B and HBV reactivation. J. Hepatol. 2021, 75, 302–310. [Google Scholar] [CrossRef]
- Mueller, S.N.; Ahmed, R. High antigen levels are the cause of T cell exhaustion during chronic viral infection. Proc. Natl. Acad. Sci. USA 2009, 106, 8623–8628. [Google Scholar] [CrossRef] [Green Version]
- Wherry, E.J.; Blattman, J.N.; Murali-Krishna, K.; van der Most, R.; Ahmed, R. Viral persistence alters CD8 T-cell immunodominance and tissue distribution and results in distinct stages of functional impairment. J. Virol. 2003, 77, 4911–4927. [Google Scholar] [CrossRef] [Green Version]
- Fuller, M.J.; Khanolkar, A.; Tebo, A.E.; Zajac, A.J. Maintenance, loss, and resurgence of T cell responses during acute, protracted, and chronic viral infections. J. Immunol. 2004, 172, 4204–4214. [Google Scholar] [CrossRef]
- Bucks, C.M.; Norton, J.A.; Boesteanu, A.C.; Mueller, Y.M.; Katsikis, P.D. Chronic antigen stimulation alone is sufficient to drive CD8+ T cell exhaustion. J. Immunol. 2009, 182, 6697–6708. [Google Scholar] [CrossRef] [Green Version]
- Tay, S.S.; Wong, Y.C.; McDonald, D.M.; Wood, N.A.W.; Roediger, B.; Sierro, F.; Mcguffog, C.; Alexander, I.E.; Bishop, G.A.; Gamble, J.R.; et al. Antigen expression level threshold tunes the fate of CD8 T cells during primary hepatic immune responses. Proc. Natl. Acad. Sci. USA 2014, 111, E2540–E2549. [Google Scholar] [CrossRef] [Green Version]
- Isogawa, M.; Chung, J.; Murata, Y.; Kakimi, K.; Chisari, F.V. CD40 activation rescues antiviral CD8+ T cells from PD-1-mediated exhaustion. PLoS Pathog. 2013, 9, e1003490. [Google Scholar] [CrossRef] [Green Version]
- Bénéchet, A.P.; De Simone, G.; Di Lucia, P.; Cilenti, F.; Barbiera, G.; Le Bert, N.; Fumagalli, V.; Lusito, E.; Moalli, F.; Bianchessi, V.; et al. Dynamics and genomic landscape of CD8+ T cells undergoing hepatic priming. Nature 2019, 574, 200–205. [Google Scholar] [CrossRef]
- Fumagalli, V.; Di Lucia, P.; Venzin, V.; Bono, E.B.; Jordan, R.; Frey, C.R.; Delaney, W.; Chisari, F.V.; Guidotti, L.G.; Iannacone, M. Serum HBsAg clearance has minimal impact on CD8+ T cell responses in mouse models of HBV infection. J. Exp. Med. 2020, 217, e20200298. [Google Scholar] [CrossRef]
- Michler, T.; Kosinska, A.D.; Festag, J.; Bunse, T.; Su, J.; Ringelhan, M.; Imhof, H.; Grimm, D.; Steiger, K.; Mogler, C.; et al. Knockdown of virus antigen expression increases therapeutic vaccine efficacy in high-titer hepatitis B virus carrier mice. Gastroenterology 2020, 158, 1762–1775.e9. [Google Scholar] [CrossRef]
- Zhu, D.; Liu, L.; Yang, D.; Fu, S.; Bian, Y.; Sun, Z.; He, J.; Su, L.; Zhang, L.; Peng, H.; et al. Clearing persistent extracellular antigen of hepatitis B virus: An immunomodulatory strategy to reverse tolerance for an effective therapeutic vaccination. J. Immunol. 2016, 196, 3079–3087. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, Y.; Gautam, R.; Chun, T.-W.; Sadjadpour, R.; Foulds, K.E.; Shingai, M.; Klein, F.; Gazumyan, A.; Golijanin, J.; Donaldson, M.; et al. Early antibody therapy can induce long-lasting immunity to SHIV. Nature 2017, 543, 559–563. [Google Scholar] [CrossRef]
- Boni, C.; Penna, A.; Bertoletti, A.; Lamonaca, V.; Rapti, I.; Missale, G.; Pilli, M.; Urbani, S.; Cavalli, A.; Cerioni, S.; et al. Transient restoration of anti-viral T cell responses induced by lamivudine therapy in chronic hepatitis B. J. Hepatol. 2003, 39, 595–605. [Google Scholar] [CrossRef]
- Boni, C.; Laccabue, D.; Lampertico, P.; Giuberti, T.; Viganò, M.; Schivazappa, S.; Alfieri, A.; Pesci, M.; Gaeta, G.B.; Brancaccio, G.; et al. Restored function of HBV-specific T cells after long-term effective therapy with nucleos(t)ide analogues. Gastroenterology 2012, 143, 963–973.e9. [Google Scholar] [CrossRef]
- Stoop, J.N.; van der Molen, R.G.; Kuipers, E.J.; Kusters, J.G.; Janssen, H.L.A. Inhibition of viral replication reduces regulatory T cells and enhances the antiviral immune response in chronic hepatitis B. Virology 2007, 361, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Loggi, E.; Bihl, F.K.; Cursaro, C.; Granieri, C.; Galli, S.; Brodosi, L.; Furlini, G.; Bernardi, M.; Brander, C.; Andreone, P. Virus-specific immune response in HBeAg-negative chronic hepatitis B: Relationship with clinical profile and HBsAg serum levels. PLoS ONE 2013, 8, e65327. [Google Scholar] [CrossRef] [Green Version]
- Hoogeveen, R.C.; Robidoux, M.P.; Schwarz, T.; Heydmann, L.; Cheney, J.A.; Kvistad, D.; Aneja, J.; Melgaço, J.G.; Fernandes, C.A.; Chung, R.T.; et al. Phenotype and function of HBV-specific T cells is determined by the targeted epitope in addition to the stage of infection. Gut 2019, 68, 893–904. [Google Scholar] [CrossRef]
- Schuch, A.; Salimi Alizei, E.; Heim, K.; Wieland, D.; Kiraithe, M.M.; Kemming, J.; Llewellyn-Lacey, S.; Sogukpinar, Ö.; Ni, Y.; Urban, S.; et al. Phenotypic and functional differences of HBV core-specific versus HBV polymerase-specific CD8+ T cells in chronically HBV-infected patients with low viral load. Gut 2018, 68, 905–915. [Google Scholar] [CrossRef] [Green Version]
- Le Bert, N.; Gill, U.S.; Hong, M.; Kunasegaran, K.; Tan, D.Z.M.; Ahmad, R.; Cheng, Y.; Dutertre, C.-A.; Heinecke, A.; Rivino, L.; et al. Effects of hepatitis b surface antigen on virus-specific and global T cells in patients with chronic hepatitis B virus infection. Gastroenterology 2020, 159, 652–664. [Google Scholar] [CrossRef]
- Kim, J.H.; Ghosh, A.; Ayithan, N.; Romani, S.; Khanam, A.; Park, J.-J.; Rijnbrand, R.; Tang, L.; Sofia, M.J.; Kottilil, S.; et al. Circulating serum HBsAg level is a biomarker for HBV-specific T and B cell responses in chronic hepatitis B patients. Sci. Rep. 2020, 10, 1835. [Google Scholar] [CrossRef]
- Aliabadi, E.; Urbanek-Quaing, M.; Maasoumy, B.; Bremer, B.; Grasshoff, M.; Li, Y.; Niehaus, C.E.; Wedemeyer, H.; Kraft, A.R.M.; Cornberg, M. Impact of HBsAg and HBcrAg levels on phenotype and function of HBV-specific T cells in patients with chronic hepatitis B virus infection. Gut 2021. [Google Scholar] [CrossRef]
- Peña-Asensio, J.; Calvo, H.; Miquel, J.; Sanz-de-Villalobos, E.; González-Praetorius, A.; Torralba, M.; Larrubia, J.-R. Model to predict on-treatment restoration of functional HBV-specific CD8+ cell response foresees off-treatment HBV control in eAg-negative chronic hepatitis B. Aliment. Pharmacol. Ther. 2022. [Google Scholar] [CrossRef]
- Kondo, Y.; Ninomiya, M.; Kakazu, E.; Kimura, O.; Shimosegawa, T. Hepatitis B surface antigen could contribute to the immunopathogenesis of hepatitis B virus infection. ISRN Gastroenterol. 2013, 2013, 935295. [Google Scholar] [CrossRef]
- Tsai, K.-N.; Kuo, C.-F.; Ou, J.-H.J. Mechanisms of hepatitis B virus persistence. Trends Microbiol. 2018, 26, 33–42. [Google Scholar] [CrossRef]
- Burton, A.R.; Pallett, L.J.; McCoy, L.E.; Suveizdyte, K.; Amin, O.E.; Swadling, L.; Alberts, E.; Davidson, B.R.; Kennedy, P.T.; Gill, U.S.; et al. Circulating and intrahepatic antiviral B cells are defective in hepatitis B. J. Clin. Investig. 2018, 128, 4588–4603. [Google Scholar] [CrossRef] [Green Version]
- Salimzadeh, L.; Le Bert, N.; Dutertre, C.-A.; Gill, U.S.; Newell, E.W.; Frey, C.; Hung, M.; Novikov, N.; Fletcher, S.; Kennedy, P.T.; et al. PD-1 blockade partially recovers dysfunctional virus–specific B cells in chronic hepatitis B infection. J. Clin. Investig. 2018, 128, 4573–4587. [Google Scholar] [CrossRef]
- Le Bert, N.; Salimzadeh, L.; Gill, U.S.; Dutertre, C.-A.; Facchetti, F.; Tan, A.; Hung, M.; Novikov, N.; Lampertico, P.; Fletcher, S.P.; et al. Comparative characterization of B cells specific for HBV nucleocapsid and envelope proteins in patients with chronic hepatitis B. J. Hepatol. 2020, 72, 34–44. [Google Scholar] [CrossRef]
- Huang, D.; Wu, D.; Wang, P.; Wang, Y.; Yuan, W.; Hu, D.; Hu, J.; Wang, Y.; Tao, R.; Xiao, F.; et al. End-of-treatment HBcrAg and HBsAb levels identify durable functional cure after Peg-IFN-based therapy in patients with CHB. J. Hepatol. 2022. [Google Scholar] [CrossRef]
- Sonneveld, M.J.; Chiu, S.-M.; Park, J.Y.; Brakenhoff, S.M.; Kaewdech, A.; Seto, W.-K.; Tanaka, Y.; Carey, I.; Papatheodoridi, M.; van Bömmel, F.; et al. Probability of HBsAg loss after nucleo(s)tide analogue withdrawal depends on HBV genotype and viral antigen levels. J. Hepatol. 2022, 76, 1042–1050. [Google Scholar] [CrossRef] [PubMed]
- García-López, M.; Lens, S.; Pallett, L.J.; Testoni, B.; Rodríguez-Tajes, S.; Mariño, Z.; Bartres, C.; García-Pras, E.; Leonel, T.; Perpiñán, E.; et al. Viral and immune factors associated with successful treatment withdrawal in HBeAg-negative chronic hepatitis B patients. J. Hepatol. 2021, 74, 1064–1074. [Google Scholar] [CrossRef] [PubMed]
- Hirode, G.; Choi, H.S.J.; Chen, C.-H.; Su, T.-H.; Seto, W.-K.; Van Hees, S.; Papatheodoridi, M.; Lens, S.; Wong, G.; Brakenhoff, S.M.; et al. Off-therapy response after nucleos(t)ide analogue withdrawal in patients with chronic hepatitis B: An international, multicenter, multiethnic cohort (RETRACT-B study). Gastroenterology 2022, 162, 757–771.e4. [Google Scholar] [CrossRef] [PubMed]
- Hui, R.W.-H.; Mak, L.-Y.; Seto, W.-K.; Yuen, M.-F. RNA interference as a novel treatment strategy for chronic hepatitis B infection. Clin. Mol. Hepatol. 2022, 13, 581. [Google Scholar] [CrossRef]
- Vaillant, A. Nucleic acid polymers: Broad spectrum antiviral activity, antiviral mechanisms and optimization for the treatment of hepatitis B and hepatitis D infection. Antivir. Res. 2016, 133, 32–40. [Google Scholar] [CrossRef] [Green Version]
- Khan, I.W.; Dad Ullah, M.U.; Choudhry, M.; Ali, M.J.; Ali, M.A.; Lam, S.L.K.; Shah, P.A.; Kaur, S.P.; Lau, D.T.Y. Novel therapies of hepatitis B and D. Microorganisms 2021, 9, 2607. [Google Scholar] [CrossRef]
- Blanchet, M.; Sinnathamby, V.; Vaillant, A.; Labonté, P. Inhibition of HBsAg secretion by nucleic acid polymers in HepG2.2.15 cells. Antivir. Res. 2019, 164, 97–105. [Google Scholar] [CrossRef]
- Corti, D.; Benigni, F.; Shouval, D. Viral envelope-specific antibodies in chronic hepatitis B virus infection. Curr. Opin. Virol. 2018, 30, 48–57. [Google Scholar] [CrossRef]
- Yuen, M.F.; Schiefke, I.; Yoon, J.H. RNA Interference therapy with ARC-520 results in prolonged hepatitis B surface antigen response in patients with chronic hepatitis B infection. J. Hepatol. 2020, 72, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Yuen, M.F.; Wong, D.K.; Schluep, T. Long-term serological, virological and histological responses to RNA inhibition by ARC-520 in Chinese chronic hepatitis B patients on entecavir treatment. Gut 2022, 71, 789–797. [Google Scholar] [CrossRef]
- Gane, E.; Locarnini, S.; Lim, T.H. Short term treatment with RNA interference therapy, JNJ-3989, results in sustained hepatitis B surface antigen supression in patients with chronic hepatitis B receiving nucleos(t)ide analogue treatment. J. Hepatol. 2020, 73, S20. [Google Scholar] [CrossRef]
- Yuen, M.F.; Berliba, E.; Kim, Y.J. Safety and pharmacodynamics of the GalNAc-siRNA AB-729 in subjects with chronic hepatitis B infection. J. Hepatol. 2020, 72, 62A–63A. [Google Scholar]
- Gane, E. Preliminary safety and antiviral activity of VIR-2218, an X-targeting RNAi therapeutic, in chronic hepatitis B patients. In Proceedings of the International Liver Congress™ EASL—European Association for the Study of the Liver, London, UK, 15–19 April 2020. [Google Scholar]
- Yuen, M.F. HBV RNAi inhibitor RG6346 in Phase 1b-2a trial was safe, well-tolerated, and resulted in substantial and durable reductions in serum HBsAg levels. In Proceedings of the AASLD Digital 351, Virtual, 13–16 November 2020. [Google Scholar]
- Streinu-Cercel, A.; Gane, E.; Cheng, W. A phase 2a study evaluating the multi-dose activity of ARB-1467 in HBeAg positive and negative virally suppressed subjects with hepatitis B. J. Hepatol. 2017, 66, S688–S689. [Google Scholar] [CrossRef]
- Thi, E.P.; Dhillon, A.P.; Ardzinski, A. ARB-1740, a RNA interference therapeutic for chronic hepatitis B infection. ACS Infect. Dis. 2019, 5, 725–737. [Google Scholar] [CrossRef]
- Yuen, M.F.; Heo, J.; Jang, J.W. Safety, tolerability and antiviral activity of the antisense oligonucleotide bepirovirsen in patients with chronic hepatitis B: A phase 2 randomized controlled trial. Nat. Med. 2021, 27, 1725–1734. [Google Scholar] [CrossRef]
- Han, K.; Cremer, J.; Elston, R. A Randomized, double-blind, placebo-controlled, first-time-in-human study to assess the safety, tolerability, and pharmacokinetics of single and multiple ascending doses of GSK3389404 in healthy subjects. Clin. Pharmacol. Drug Dev. 2019, 8, 790–801. [Google Scholar] [CrossRef] [Green Version]
- Yuen, M.F.; Gane, E.; Kim, D.J. RO7062931 antisense oligonucleotide phase 1 study demonstrates target engagement in patients with chronic hepatitis B on established nucleos(t)ide therapy. J. Hepatol. 2020, 73, S51. [Google Scholar] [CrossRef]
- Bazinet, M.; Pântea, V.; Placinta, G. Safety and efficacy of 48 weeks REP 2139 or REP 2165, tenofovir disoproxil, and pegylated interferon alfa-2a in patients with chronic HBV infection naïve to nucleos(t)ide therapy. Gastroenterology 2020, 158, 2180–2194. [Google Scholar] [CrossRef]
- Bazinet, M.; Pântea, V.; Cebotarescu, V. Persistent control of hepatitis B virus and hepatitis delta virus infection following REP 2139-Ca and pegylated interferon therapy in chronic hepatitis B virus/hepatitis delta virus coinfection. Hepatol. Commun. 2020, 5, 189–202. [Google Scholar] [CrossRef]
- Bazinet, M.; Pântea, V.; Cebotarescu, V. Safety and efficacy of REP 2139 and pegylated interferon alfa-2a for treatment-naïve patients with chronic hepatitis B virus and hepatitis D virus co-infection (REP 301 and REP 301-LTF): A non-randomised, open-label, phase 2 trial. Lancet Gastroenterol. Hepatol. 2017, 2, 877–889. [Google Scholar] [CrossRef]
- Al-Mahtab, M.; Bazinet, M.; Vaillant, A. Safety and efficacy of nucleic acid polymers in monotherapy and combined with immunotherapy in treatment-naïve Bangladeshi patients with HBeAg+ chronic hepatitis B infection. PLoS ONE 2016, 11, e0156667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gohil, V.; Misner, D.; Chanda, S. The S-antigen transport-inhibiting oligonucleotide polymer (STOPS™) ALG-010133 demonstrates a favorable preclinical profile for the treatment of chronic hepatitis, B. Hepatology 2020, 72, 508A. [Google Scholar]
- Lee, H.W.; Park, J.Y.; Hong, T. A prospective, open-label, dose-escalation, single-center, phase 1 study for GC1102, a recombinant human immunoglobulin for chronic hepatitis B patients. Hepatology 2018, 68, 268A–269A. [Google Scholar]
- Gupta, S.V.; Arizpe, A.; Fanget, M.C. Preliminary pharmacokinetics and safety in healthy volunteers of VIR-3434, a monoclonal antibody for the treatment of chronichepatitis B infection. J. Hepatol. 2021, 75, S733. [Google Scholar]
- Agarwal, K.; Yuen, M.F.; Wedemeyer, H. A phase 1 study evaluating the neutralizing, vaccinal monoclonal antibody VIR-3434 in participants with chronic hepatitisB virus infection. J. Hepatol. 2021, 75, S288. [Google Scholar]


{kind=link}
| Class of Agents | Drug | References |
|---|---|---|
| Small interfering RNAs (siRNA) | ARC520 | [8,59,60] |
| JNJ-3989 | [61] | |
| AB-729 | [62] | |
| VIR-2218 | [63] | |
| RG6346 | [64] | |
| ARB-1467 | [65] | |
| ARB-1740 | [66] | |
| Antisense Oligonucleotides (ASO) | BEPIROVERSEN | [67] |
| GSK3389404 | [68] | |
| RO7062931 | [69] | |
| Nucleic Acid Polymers (NAP) | REP 2139 | [70,71,72,73] |
| REP 2165 | [70] | |
| REP 2055 | [73] | |
| ALG 010133 | [74] | |
| Monoclonal Antibodies (Mab) | GC1102 | [75] |
| VIR-3434 | [76,77] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montali, I.; Vecchi, A.; Rossi, M.; Tiezzi, C.; Penna, A.; Reverberi, V.; Laccabue, D.; Missale, G.; Boni, C.; Fisicaro, P. Antigen Load and T Cell Function: A Challenging Interaction in HBV Infection. Biomedicines 2022, 10, 1224. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10061224
Montali I, Vecchi A, Rossi M, Tiezzi C, Penna A, Reverberi V, Laccabue D, Missale G, Boni C, Fisicaro P. Antigen Load and T Cell Function: A Challenging Interaction in HBV Infection. Biomedicines. 2022; 10(6):1224. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10061224
Chicago/Turabian StyleMontali, Ilaria, Andrea Vecchi, Marzia Rossi, Camilla Tiezzi, Amalia Penna, Valentina Reverberi, Diletta Laccabue, Gabriele Missale, Carolina Boni, and Paola Fisicaro. 2022. "Antigen Load and T Cell Function: A Challenging Interaction in HBV Infection" Biomedicines 10, no. 6: 1224. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10061224