
Expression of Apoptosis-Related Biomarkers in Inflamed Nasal Sinus Epithelium of Patients with Chronic Rhinosinusitis with Nasal Polyps (CRSwNP)—Evaluation at mRNA and miRNA Levels

 , , ,
, , ,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. TUNEL-Assay
2.3. Quantitative Real-Time Reverse Transcription Polymerase Chain Reaction (RT-qPCR)
2.4. Statistical Analysis
3. Results
3.1. Apoptosis In Situ
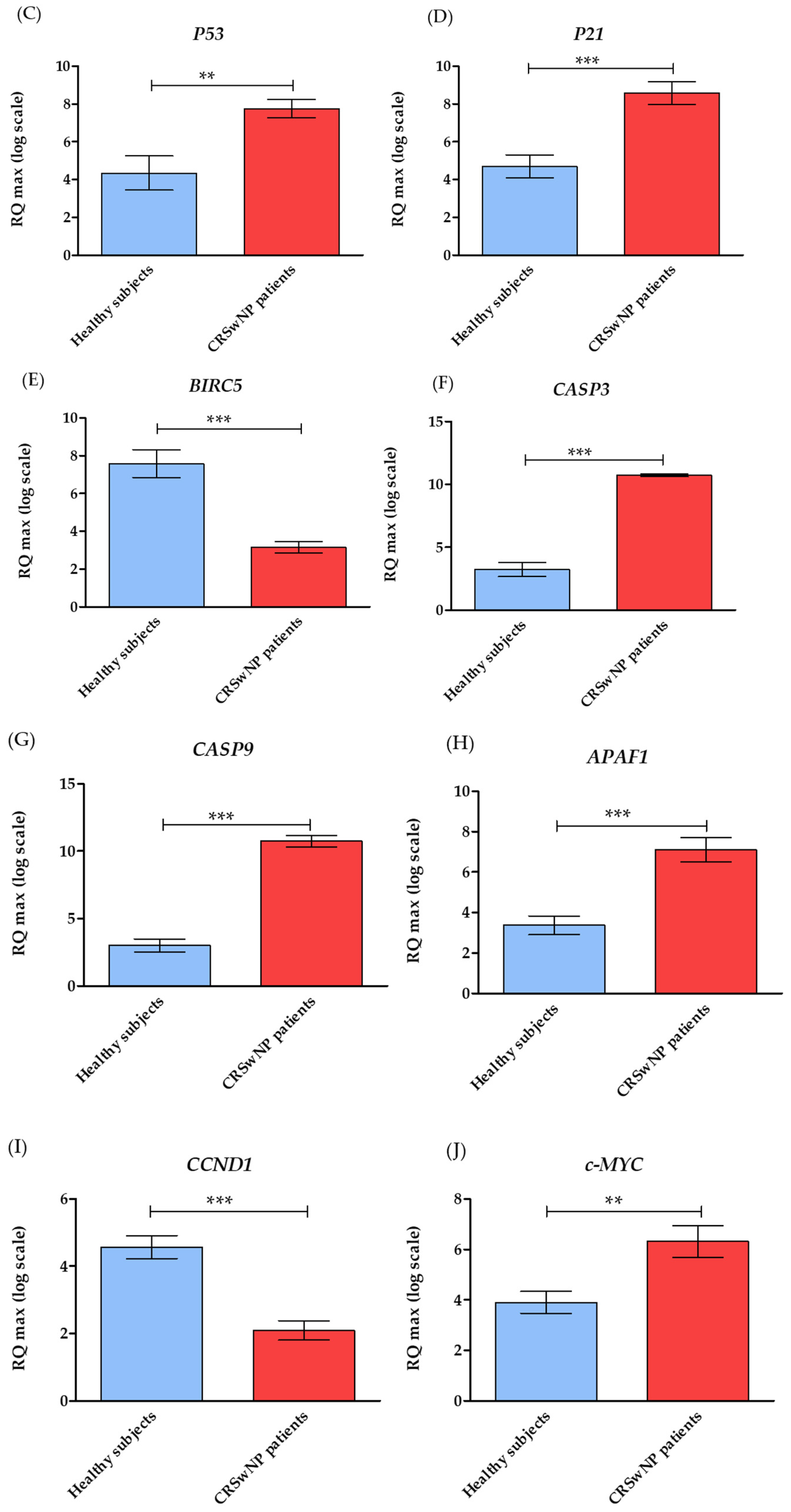
3.2. The Level of Genes Associated with Apoptosis in CRSwNP Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- White, S.R. Apoptosis and the Airway Epithelium. J. Allergy 2011, 2011, 948406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soroosh, A.; Fang, K.; Hoffman, J.M.; Law, I.K.M.; Videlock, E.; Lokhandwala, Z.A.; Zhao, J.J.; Hamidi, S.; Padua, D.M.; Frey, M.R.; et al. Correction: Loss of MiR-24-3p Promotes Epithelial Cell Apoptosis and Impairs the Recovery from Intestinal Inflammation. Cell Death Dis. 2022, 13, 8. [Google Scholar] [CrossRef] [PubMed]
- Lam, K.; Schleimer, R.; Kern, R.C. The Etiology and Pathogenesis of Chronic Rhinosinusitis: A Review of Current Hypotheses. Curr. Allergy Asthma Rep. 2015, 15, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shih, L.C.; Hsieh, H.H.; Tsay, G.J.; Lee, I.T.; Tsou, Y.A.; Lin, C.L.; Shen, T.C.; Bau, D.T.; Tai, C.J.; Der Lin, C.; et al. Chronic Rhinosinusitis and Premorbid Autoimmune Diseases: A Population-Based Case–Control Study. Sci. Rep. 2020, 10, 18635. [Google Scholar] [CrossRef]
- Yoshikawa, T.; Minaga, K.; Hara, A.; Sekai, I.; Otsuka, Y.; Takada, R.; Kamata, K.; Watanabe, T.; Kudo, M. A Unique Profile of Serum Cytokines in Type 1 Autoimmune Pancreatitis and Chronic Rhinosinusitis. Asian Pac. J. Allergy Immunol. Case Rep. 2021. Epub ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Staudacher, A.G.; Peters, A.T.; Kato, A.; Stevens, W.W. Use of Endotypes, Phenotypes, and Inflammatory Markers to Guide Treatment Decisions in Chronic Rhinosinusitis. Ann. Allergy Asthma Immunol. 2020, 124, 318–325. [Google Scholar] [CrossRef] [Green Version]
- Yao, Y.; Xie, S.; Yang, C.; Zhang, J.; Wu, X.; Sun, H. Biomarkers in the Evaluation and Management of Chronic Rhinosinusitis with Nasal Polyposis. Eur. Arch. Oto-Rhino-Laryngol. 2017, 274, 3559–3566. [Google Scholar] [CrossRef]
- Fujieda, S.; Imoto, Y.; Kato, Y.; Ninomiya, T.; Tokunaga, T.; Tsutsumiuchi, T.; Yoshida, K.; Kidoguchi, M.; Takabayashi, T. Eosinophilic Chronic Rhinosinusitis. Allergol. Int. 2019, 68, 403–412. [Google Scholar] [CrossRef]
- Wang, H.; Li, Z.Y.; Jiang, W.X.; Liao, B.; Zhai, G.T.; Wang, N.; Zhen, Z.; Ruan, J.W.; Long, X.B.; Wang, H.; et al. The Activation and Function of IL-36γ in Neutrophilic Inflammation in Chronic Rhinosinusitis. J. Allergy Clin. Immunol. 2018, 141, 1646–1658. [Google Scholar] [CrossRef] [Green Version]
- Cao, P.P.; Wang, Z.C.; Schleimer, R.P.; Liu, Z. Pathophysiologic Mechanisms of Chronic Rhinosinusitis and Their Roles in Emerging Disease Endotypes. Ann. Allergy Asthma Immunol. 2019, 122, 33–40. [Google Scholar] [CrossRef] [Green Version]
- Tomassen, P.; Vandeplas, G.; Van Zele, T.; Cardell, L.O.; Arebro, J.; Olze, H.; Förster-Ruhrmann, U.; Kowalski, M.L.; Olszewska-Ziaber, A.; Holtappels, G.; et al. Inflammatory Endotypes of Chronic Rhinosinusitis Based on Cluster Analysis of Biomarkers. J. Allergy Clin. Immunol. 2016, 137, 1449–1456. [Google Scholar] [CrossRef] [Green Version]
- Newton, J.R.; Ah-See, K.W. A Review of Nasal Polyposis. Ther. Clin. Risk Manag. 2008, 4, 507–512. [Google Scholar] [CrossRef] [Green Version]
- Coste, A.; Rateau, J.G.; Roudot-Thoraval, F.; Chapelin, C.; Gilain, L.; Poron, F.; Peynègre, R.; Bernaudin, J.F.; Escudier, E. Increased Epithelial Cell Proliferation in Nasal Polyps. Arch. Otolaryngol.-Head Neck Surg. 1996, 122, 432–436. [Google Scholar] [CrossRef]
- Hsu, M.C.; Shun, C.T.; Liu, C.M. Increased Epithelial Cell Proliferation in Nasal Polyps. J. Formos. Med. Assoc. 2002, 101, 227–229. [Google Scholar]
- Wu, C.C.; Lee, T.J.; Chang, P.H.; Tsai, C.N.; Lee, Y.S.; Fu, C.H.; Huang, C.C. Similar Cellular Proliferation Activities in Nasal Polyps and Adjacentinferior Turbinate. Am. J. Otolaryngol. Head Neck Med. Surg. 2012, 33, 14–19. [Google Scholar] [CrossRef]
- Chao, S. Role of Delayed Cellular Apoptosis in the Pathogenesis of Nasal Polyps. J. Allergy Clin. Immunol. 2004, 113, S280. [Google Scholar] [CrossRef]
- Küpper, D.S.; Valera, F.C.P.; Malinsky, R.; Milanezi, C.M.; Silva, J.S.; Tamashiro, E.; Anselmo-Lima, W.T. Expression of Apoptosis Mediators P53 and Caspase 3, 7, and 9 in Chronic Rhinosinusitis with Nasal Polyposis. Am. J. Rhinol. Allergy 2014, 28, 187–191. [Google Scholar] [CrossRef]
- Cohen, L.; Xueping, E.; Tarsi, J.; Ramkumar, T.; Horiuchi, T.K.; Cochran, R.; DeMartino, S.; Schechtman, K.B.; Hussain, I.; Holtzman, M.J.; et al. Epithelial Cell Proliferation Contributes to Airway Remodeling in Severe Asthma. Am. J. Respir. Crit. Care Med. 2007, 176, 138–145. [Google Scholar] [CrossRef] [Green Version]
- Chan, S.L.; Yu, V.C. Proteins of the Bcl-2 Family in Apoptosis Signalling: From Mechanistic Insights to Therapeutic Opportunities. Clin. Exp. Pharmacol. Physiol. 2004, 31, 119–128. [Google Scholar] [CrossRef]
- Ladokhin, A.S. Regulation of Apoptosis by the Bcl-2 Family of Proteins: Field on a Brink. Cells 2020, 9, 2121. [Google Scholar] [CrossRef]
- Christgen, S.; Tweedell, R.E.; Kanneganti, T.D. Programming Inflammatory Cell Death for Therapy. Pharmacol. Ther. 2022, 232, 108010. [Google Scholar] [CrossRef]
- Zou, H.; Li, Y.; Liu, X.; Wang, X. An APAf-1 Cytochrome C Multimeric Complex Is a Functional Apoptosome That Activates Procaspase-9. J. Biol. Chem. 1999, 274, 11549–11556. [Google Scholar] [CrossRef] [Green Version]
- Gross, A. BCL-2 Family Proteins as Regulators of Mitochondria Metabolism. Biochim. Biophys. Acta Bioenerg. 2016, 1857, 1243–1246. [Google Scholar] [CrossRef]
- Vucicevic, K.; Jakovljevic, V.; Colovic, N.; Tosic, N.; Kostic, T.; Glumac, I.; Pavlovic, S.; Karan-Djurasevic, T.; Colovic, M. Association of Bax Expression and Bcl2/Bax Ratio with Clinical and Molecular Prognostic Markers in Chronic Lymphocytic Leukemia. J. Med. Biochem. 2016, 35, 150. [Google Scholar] [CrossRef] [Green Version]
- Zhan, Y.; Zhu, M.; Liu, S.; Lu, J.; Ni, Z.; Cai, H.; Zhang, W. MicroRNA-93 Inhibits the Apoptosis and Inflammatory Response of Tubular Epithelial Cells via the PTEN/AKT/MTOR Pathway in Acute Kidney Injury. Mol. Med. Rep. 2021, 24, 1–11. [Google Scholar] [CrossRef]
- Muñoz-Carrillo, J.L.; Vázquez-Alcaraz, S.J.; Vargas-Barbosa, J.M.; Ramos-Gracia, L.G.; Alvarez-Barreto, I.; Medina-Quiroz, A.; Díaz-Huerta, K.K. The Role of microRNAs in Pulp Inflammation. Cells 2021, 10, 2142. [Google Scholar] [CrossRef]
- Jiang, Y.; Xu, X.; Xiao, L.; Wang, L.; Qiang, S. The Role of MicroRNA in the Inflammatory Response of Wound Healing. Front. Immunol. 2022, 13, 1231. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Chen, J.; Zhao, Y.; Yang, J.; Xue, K.; Wang, Z. MicroRNA-761 Suppresses Remodeling of Nasal Mucosa and Epithelial-Mesenchymal Transition in Mice with Chronic Rhinosinusitis through LCN2. Stem Cell Res. Ther. 2020, 11, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.H.; Zhang, Y.N.; Li, H.B.; Hu, C.Y.; Wang, N.; Cao, P.P.; Liao, B.; Lu, X.; Cui, Y.H.; Liu, Z. Overexpression of MiR-125b, a Novel Regulator of Innate Immunity, in Eosinophilic Chronic Rhinosinusitis with Nasal Polyps. Am. J. Respir. Crit. Care Med. 2012, 185, 140–151. [Google Scholar] [CrossRef]
- Fokkens, W.J.; Lund, V.J.; Hopkins, C.; Hellings, P.W.; Kern, R.; Reitsma, S.; Toppila-Salmi, S.; Bernal-Sprekelsen, M.; Mullol, J. Executive Summary of EPOS 2020 Including Integrated Care Pathways. Rhinology 2020, 58, 82–111. [Google Scholar] [CrossRef]
- Śmieszek, A.; Stręk, Z.; Kornicka, K.; Grzesiak, J.; Weiss, C.; Marycz, K. Antioxidant and Anti-Senescence Effect of Metformin on Mouse Olfactory Ensheathing Cells (MOECs) May Be Associated with Increased Brain-Derived Neurotrophic Factor Levels—An Ex Vivo Study. Int. J. Mol. Sci. 2017, 18, 872. [Google Scholar] [CrossRef] [PubMed]
- Smieszek, A.; Marcinkowska, K.; Pielok, A.; Sikora, M.; Valihrach, L.; Marycz, K. The Role of MiR-21 in Osteoblasts-Osteoclasts Coupling In Vitro. Cells 2020, 9, 479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smieszek, A.; Kornicka, K.; Szłapka-Kosarzewska, J.; Androvic, P.; Valihrach, L.; Langerova, L.; Rohlova, E.; Kubista, M.; Marycz, K. Metformin Increases Proliferative Activity and Viability of Multipotent Stromal Stem Cells Isolated from Adipose Tissue Derived from Horses with Equine Metabolic Syndrome. Cells 2019, 8, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chomczynski, P.; Sacchi, N. Single-Step Method of RNA Isolation by Acid Guanidinium Thiocyanate-Phenol-Chloroform Extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Khalmuratova, R.; Lee, M.; Mo, J.H.; Jung, Y.J.; Park, J.W.; Shin, H.W. Wogonin Attenuates Nasal Polyp Formation by Inducing Eosinophil Apoptosis through HIF-1α and Survivin Suppression. Sci. Rep. 2018, 8, 6201. [Google Scholar] [CrossRef]
- Fan, G.K.; Itoh, T.; Imanaka, M.; Fujieda, S.; Takenaka, H. Eosinophilic Apoptosis in Sinus Mucosa: Relationship to Tissue Eosinophilia and Its Resolution in Allergic Sinusitis. J. Allergy Clin. Immunol. 2000, 106, 551–558. [Google Scholar] [CrossRef]
- Ge, Y.; Tsukatani, T.; Nishimura, T.; Furukawa, M.; Miwa, T. Cell Death of Olfactory Receptor Neurons in a Rat with Nasosinusitis Infected Artificially with Staphylococcus. Chem. Senses 2002, 27, 521–527. [Google Scholar] [CrossRef] [Green Version]
- Tesfaigzi, J.; Hotchkiss, J.A.; Harkema, J.R. Expression of the Bcl-2 Protein in Nasal Epithelia of F344/N Rats during Mucous Cell Metaplasia and Remodeling. Am. J. Respir. Cell Mol. Biol. 1998, 18, 794–799. [Google Scholar] [CrossRef] [Green Version]
- Ingle, R.R.; Setzen, G.; Koltai, P.J.; Monte, D.; Pastore, J.; Jennings, T.A. P53 Protein Expression in Benign Lesions of the Upper Respiratory Tract. Arch. Otolaryngol. Head Neck Surg. 1997, 123, 297–300. [Google Scholar] [CrossRef]
- Riley, T.; Sontag, E.; Chen, P.; Levine, A. Transcriptional Control of Human P53-Regulated Genes. Nat. Rev. Mol. Cell Biol. 2008, 9, 402–412. [Google Scholar] [CrossRef]
- Viksne, R.J.; Sumeraga, G.; Pilmane, M. Characterization of Cytokines and Proliferation Marker Ki67 in Chronic Rhinosinusitis with Nasal Polyps: A Pilot Study. Medicina 2021, 57, 607. [Google Scholar] [CrossRef]
- Aylon, Y.; Oren, M. The Paradox of P53: What, How, and Why? Cold Spring Harb. Perspect. Med. 2016, 6, a026328. [Google Scholar] [CrossRef] [Green Version]
- Lavezzi, A.; Mantovani, M.; Cazzullo, A.; Turconi, P.; Matturri, L. P53 Over-Expression and Its Correlation with PCNA Index in Nasal Polyps. Rhinology 1999, 37, 160–163. [Google Scholar]
- Garavello, W.; Viganò, P.; Romagnoli, M.; Sordo, L.; Berti, E.; Tredici, G.; Gaini, R.M. Expression of Cell Cycle Regulatory Proteins and Analysis of Apoptosis in Normal Nasal Mucosa and in Nasal Polyps. Am. J. Rhinol. 2005, 19, 549–553. [Google Scholar] [CrossRef]
- Chalastras, T.; Athanassiadou, P.; Patsouris, E.; Eleftheriadou, A.; Kandiloros, D.; Papaxoinis, K.; Nicolopoulou-Stamati, P. Differential Rates of Proliferation and Apoptosis in Nasal Polyps Correspond to Alterations in DNA Spatial Distribution and Nuclear Polarization as Observed by Confocal Microscopy. Eur. Arch. Oto-Rhino-Laryngol. 2010, 267, 1075–1080. [Google Scholar] [CrossRef]
- Asada, M.; Yamada, T.; Ichijo, H.; Delia, D.; Miyazono, K.; Fukumuro, K.; Mizutani, S. Apoptosis Inhibitory Activity of Cytoplasmic P21(Cip1/WAF1) in Monocytic Differentiation. EMBO J. 1999, 18, 1223–1234. [Google Scholar] [CrossRef] [Green Version]
- Jung, P.; Hermeking, H. The C-MYC-AP4-P21 Cascade. Cell Cycle 2009, 8, 982–989. [Google Scholar] [CrossRef]
- Noori, A.R.; Tashakor, A.; Nikkhah, M.; Eriksson, L.A.; Hosseinkhani, S.; Fearnhead, H.O. Loss of WD2 Subdomain of Apaf-1 Forms an Apoptosome Structure Which Blocks Activation of Caspase-3 and Caspase-9. Biochimie 2021, 180, 23–29. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, N.; Bo, M.; Holtappels, G.; Zheng, M.; Lou, H.; Wang, H.; Zhang, L.; Bachert, C. Diversity of TH Cytokine Profiles in Patients with Chronic Rhinosinusitis: A Multicenter Study in Europe, Asia, and Oceania. J. Allergy Clin. Immunol. 2016, 138, 1344–1353. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.H.; Sang, H.L.; Kim, K.R.; Lee, H.M.; Lee, S.H.; Kim, T.H. Expression and Distributional Patterns of the Inhibitor of Apoptosis Protein Family and Caspase 3 in Nasal Polyps. Arch. Otolaryngol. Head Neck Surg. 2008, 134, 316–321. [Google Scholar] [CrossRef]
- Hou, H.; Xing, W.; Li, W. Brahma-Related Gene 1 Induces Apoptosis in a P53-Dependent Manner in Human Rheumatoid Fibroblast-like Synoviocyte MH7A. Medicine 2016, 95, e5241. [Google Scholar] [CrossRef]
- Frazzi, R. BIRC3 and BIRC5: Multi-faceted Inhibitors in Cancer. Cell Biosci. 2021, 11, 8. [Google Scholar] [CrossRef]
- Xu, L.; Yu, W.; Xiao, H.; Lin, K. BIRC5 Is a Prognostic Biomarker Associated with Tumor Immune Cell Infiltration. Sci. Rep. 2021, 11, 390. [Google Scholar] [CrossRef]
- Cha, S.; Seo, E.H.; Lee, S.H.; Kim, K.S.; Oh, C.S.; Moon, J.S.; Kim, J.K. MicroRNA Expression in Extracellular Vesicles from Nasal Lavage Fluid in Chronic Rhinosinusitis. Biomedicines 2021, 9, 471. [Google Scholar] [CrossRef]
- Faraoni, I.; Antonetti, F.R.; Cardone, J.; Bonmassar, E. MiR-155 Gene: A Typical Multifunctional MicroRNA. Biochim. Biophys. Acta-Mol. Basis Dis. 2009, 1792, 497–505. [Google Scholar] [CrossRef]
- Li, M.; Marin-Muller, C.; Bharadwaj, U.; Chow, K.-H.; Yao, Q.; Chen, C. MicroRNAs: Control and Loss of Control in Human Physiology and Disease. World J. Surg. 2009, 33, 667–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kmiołek, T.; Paradowska-Gorycka, A. MiRNAs as Biomarkers and Possible Therapeutic Strategies in Rheumatoid Arthritis. Cells 2022, 11, 452. [Google Scholar] [CrossRef] [PubMed]
- Mi, S.; Zhang, J.; Zhang, W.; Huang, R.S. Circulating MicroRNAs as Biomarkers for Inflammatory Diseases. Microrna 2013, 2, 64–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhaumik, D.; Scott, G.K.; Schokrpur, S.; Patil, C.K.; Orjalo, A.V.; Rodier, F.; Lithgow, G.J.; Campisi, J. MicroRNAs MiR-146a/b Negatively Modulate the Senescence-Associated Inflammatory Mediators IL-6 and IL-8. Aging 2009, 1, 402. [Google Scholar] [CrossRef]
- Sun, S.; Li, L.; Dong, L.; Cheng, J.; Zhao, C.; Bao, C.; Wang, H. Circulating MRNA and MicroRNA Profiling Analysis in Patients with Ischemic Stroke. Mol. Med. Rep. 2020, 22, 792–802. [Google Scholar] [CrossRef]
- Tang, J.; Yi, S.; Liu, Y. Long Non-Coding RNA PVT1 Can Regulate the Proliferation and Inflammatory Responses of Rheumatoid Arthritis Fibroblast-like Synoviocytes by Targeting MicroRNA-145-5p. Hum. Cell 2020, 33, 1081–1090. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, L.; Zhu, W.; Chen, H.; Ding, Y.; Cui, G. Inhibition of MicroRNA-203 Protects against Traumatic Brain Injury Induced Neural Damages via Suppressing Neuronal Apoptosis and Dementia-Related Molecues. Physiol. Behav. 2021, 228, 113190. [Google Scholar] [CrossRef]
- Cimmino, A.; Calin, G.A.; Fabbri, M.; Iorio, M.V.; Ferracin, M.; Shimizu, M.; Wojcik, S.E.; Aqeilan, R.I.; Zupo, S.; Dono, M.; et al. MiR-15 and MiR-16 Induce Apoptosis by Targeting BCL2. Proc. Natl. Acad. Sci. USA 2005, 102, 13944–13949. [Google Scholar] [CrossRef] [Green Version]
- Borgdorff, V.; Bishop, C.L.; Fessart, D.; Bergin, A.H.; Overhoff, M.G.; Beach, D.H. Multiple MicroRNAs Rescue from Ras-Induced Senescence by Inhibiting P21Waf1/Cip1. Oncogene 2010, 29, 2262–2271. [Google Scholar] [CrossRef] [Green Version]
- Ugalde, A.P.; Ramsay, A.J.; De La Rosa, J.; Varela, I.; Marĩo, G.; Cadĩanos, J.; Lu, J.; Freije, J.M.P.; López-Otín, C. Aging and Chronic DNA Damage Response Activate a Regulatory Pathway Involving MiR-29 and P53. EMBO J. 2011, 30, 2219–2232. [Google Scholar] [CrossRef]
- Liu, S.; Liu, M.; Dong, L. The Clinical Value of LncRNA MALAT1 and Its Targets MiR-125b, MiR-133, MiR-146a, and MiR-203 for Predicting Disease Progression in Chronic Obstructive Pulmonary Disease Patients. J. Clin. Lab. Anal. 2020, 34, e23410. [Google Scholar] [CrossRef]
- Taganov, K.D.; Boldin, M.P.; Chang, K.-J.; Baltimore, D. NF-B-Dependent Induction of MicroRNA MiR-146, an Inhibitor Targeted to Signaling Proteins of Innate Immune Responses. Proc. Natl. Acad. Sci. USA 2006, 103, 12481–12486. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.F.; Lu, Y.J.; Wang, Z.G. MicroRNAs and Apoptosis: Implications in the Molecular Therapy of Human Disease. Clin. Exp. Pharmacol. Physiol. 2009, 36, 951–960. [Google Scholar] [CrossRef]
- Mogilyansky, E.; Rigoutsos, I. The MiR-17/92 Cluster: A Comprehensive Update on Its Genomics, Genetics, Functions and Increasingly Important and Numerous Roles in Health and Disease. Cell Death Differ. 2013, 20, 1603–1614. [Google Scholar] [CrossRef]
- Matsubara, H.; Takeuchi, T.; Nishikawa, E.; Yanagisawa, K.; Hayashita, Y.; Ebi, H.; Yamada, H.; Suzuki, M.; Nagino, M.; Nimura, Y.; et al. Apoptosis Induction by Antisense Oligonucleotides against MiR-17-5p and MiR-20a in Lung Cancers Overexpressing MiR-17-92. Oncogene 2007, 26, 6099–6105. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Tan, J.; Li, J.; Sun, X.; Du, L.; Tao, S. MiRNA-145-5p Induces Apoptosis after Ischemia-Reperfusion by Targeting Dual Specificity Phosphatase 6. J. Cell. Physiol. 2019, 234, 16281–16289. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.Y.; Zhang, Y.Y.; Zhu, B.L.; Feng, F.Z.; Zhang, H.T.; Yan, H.; Zhou, B. MiR-203a-3p Regulates the Biological Behaviors of Ovarian Cancer Cells through Mediating the Akt/GSK-3β/Snail Signaling Pathway by Targeting ATM. J. Ovarian Res. 2019, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Wang, Y.; Wang, Q.; Ghaffar, M.; Wang, Y.; Sheng, W.; Zhang, F. Increased MiR-203-3p and Reduced MiR-21-5p Synergistically Inhibit Proliferation, Migration, and Invasion in Esophageal Cancer Cells. Anti-Cancer Drugs 2019, 30, 38–45. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morawska-Kochman, M.; Śmieszek, A.; Marcinkowska, K.; Marycz, K.M.; Nelke, K.; Zub, K.; Zatoński, T.; Bochnia, M. Expression of Apoptosis-Related Biomarkers in Inflamed Nasal Sinus Epithelium of Patients with Chronic Rhinosinusitis with Nasal Polyps (CRSwNP)—Evaluation at mRNA and miRNA Levels. Biomedicines 2022, 10, 1400. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10061400
Morawska-Kochman M, Śmieszek A, Marcinkowska K, Marycz KM, Nelke K, Zub K, Zatoński T, Bochnia M. Expression of Apoptosis-Related Biomarkers in Inflamed Nasal Sinus Epithelium of Patients with Chronic Rhinosinusitis with Nasal Polyps (CRSwNP)—Evaluation at mRNA and miRNA Levels. Biomedicines. 2022; 10(6):1400. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10061400
Chicago/Turabian StyleMorawska-Kochman, Monika, Agnieszka Śmieszek, Klaudia Marcinkowska, Krzysztof Mariusz Marycz, Kamil Nelke, Krzysztof Zub, Tomasz Zatoński, and Marek Bochnia. 2022. "Expression of Apoptosis-Related Biomarkers in Inflamed Nasal Sinus Epithelium of Patients with Chronic Rhinosinusitis with Nasal Polyps (CRSwNP)—Evaluation at mRNA and miRNA Levels" Biomedicines 10, no. 6: 1400. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10061400