Development of Three-Dimensional Human Intestinal Organoids as a Physiologically Relevant Model for Characterizing the Viral Replication Kinetics and Antiviral Susceptibility of Enteroviruses

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture, Human 3D Small Intestinal Organoids and Drug Compound
2.2. Virus Strains and Titration
2.3. Virus Infection of Human Small Intestinal Organoids and Cell Lines
2.4. Viral Replication Kinetics
2.5. Cell Viability and CPE Inhibition Assays
2.6. Viral Load Reduction and Plaque Reduction Assays
2.7. Quantitative RT-PCR (qRT-PCR)
2.8. Immunofluorescence Microscopy
2.9. Statistical Analysis
3. Results
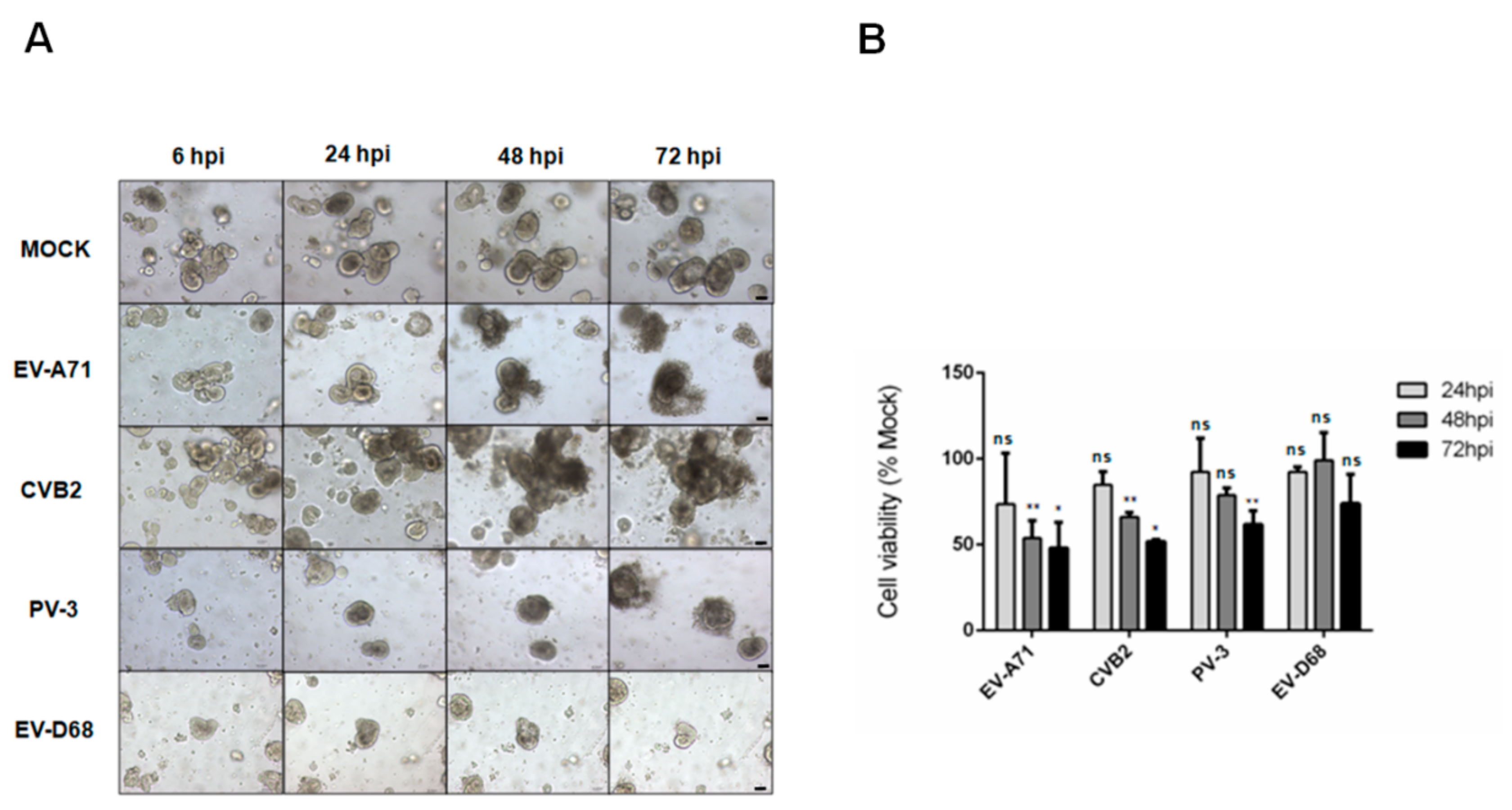
3.1. Assessment of the Susceptibility of Human Small Intestinal Organoids to Different Species of Enteroviruses by CPE and Cell Viability Assays
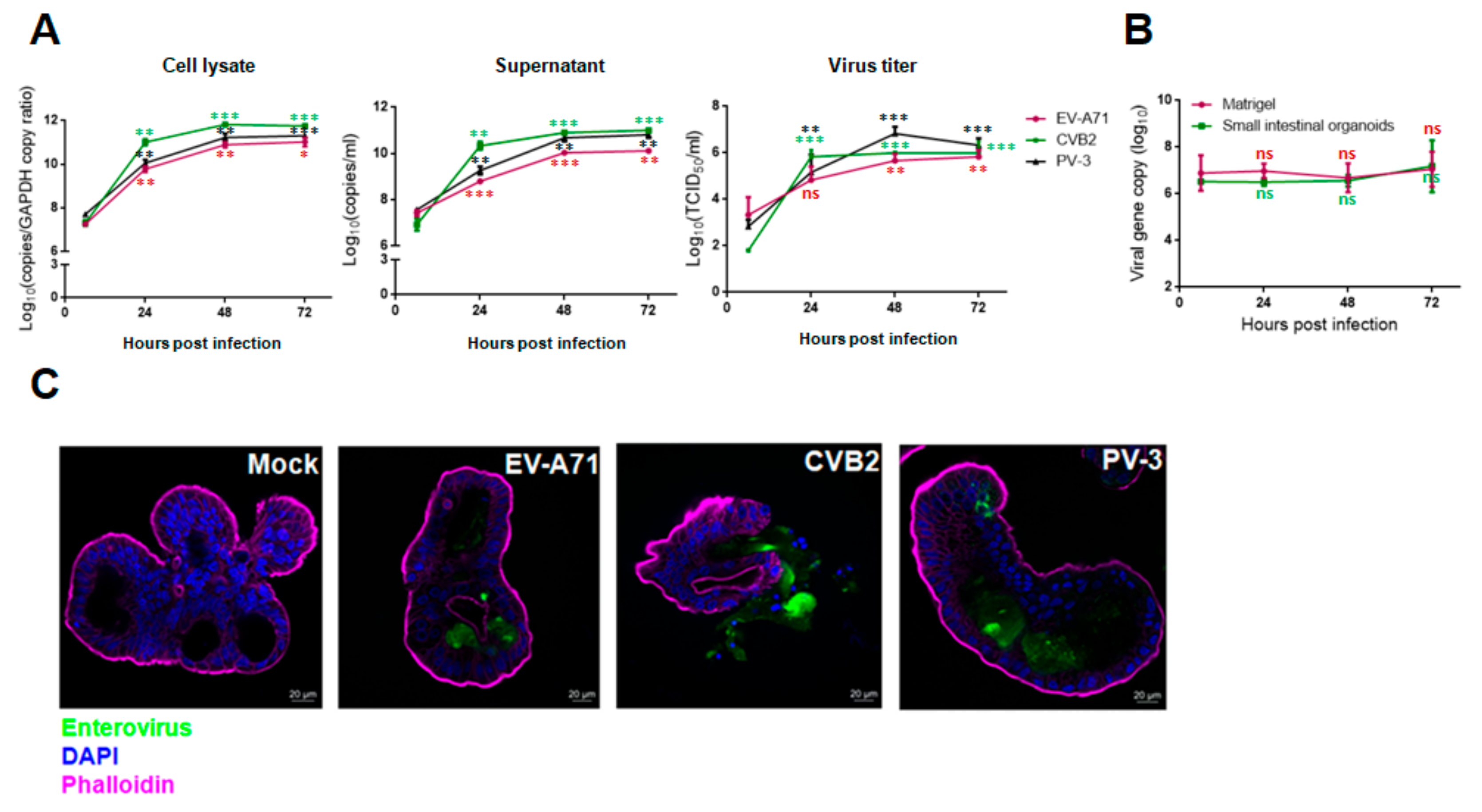
3.2. Viral Replication Kinetics, Infectious Virus Titers, and Viral Antigen Expression of EV-A71, CVB2, PV-3 and EV-D68 Infections in Human Small Intestinal Organoids
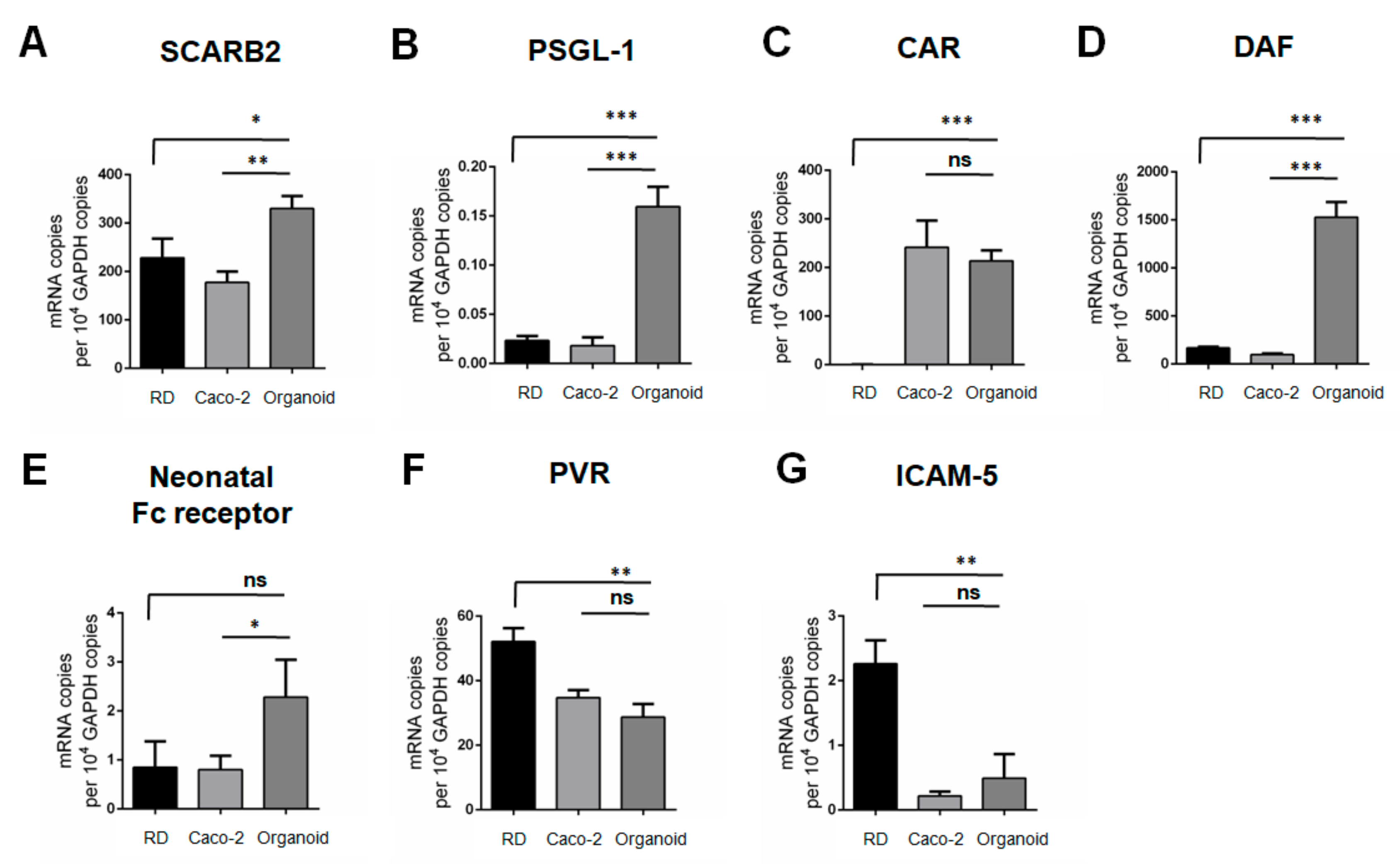
3.3. mRNA Transcript Expression Profiles of Entry Receptors of EV-A71, CVB2, PV-3 and EV-D68 in RD Cells, Caco-2 Cells and Human Small Intestinal Organoids
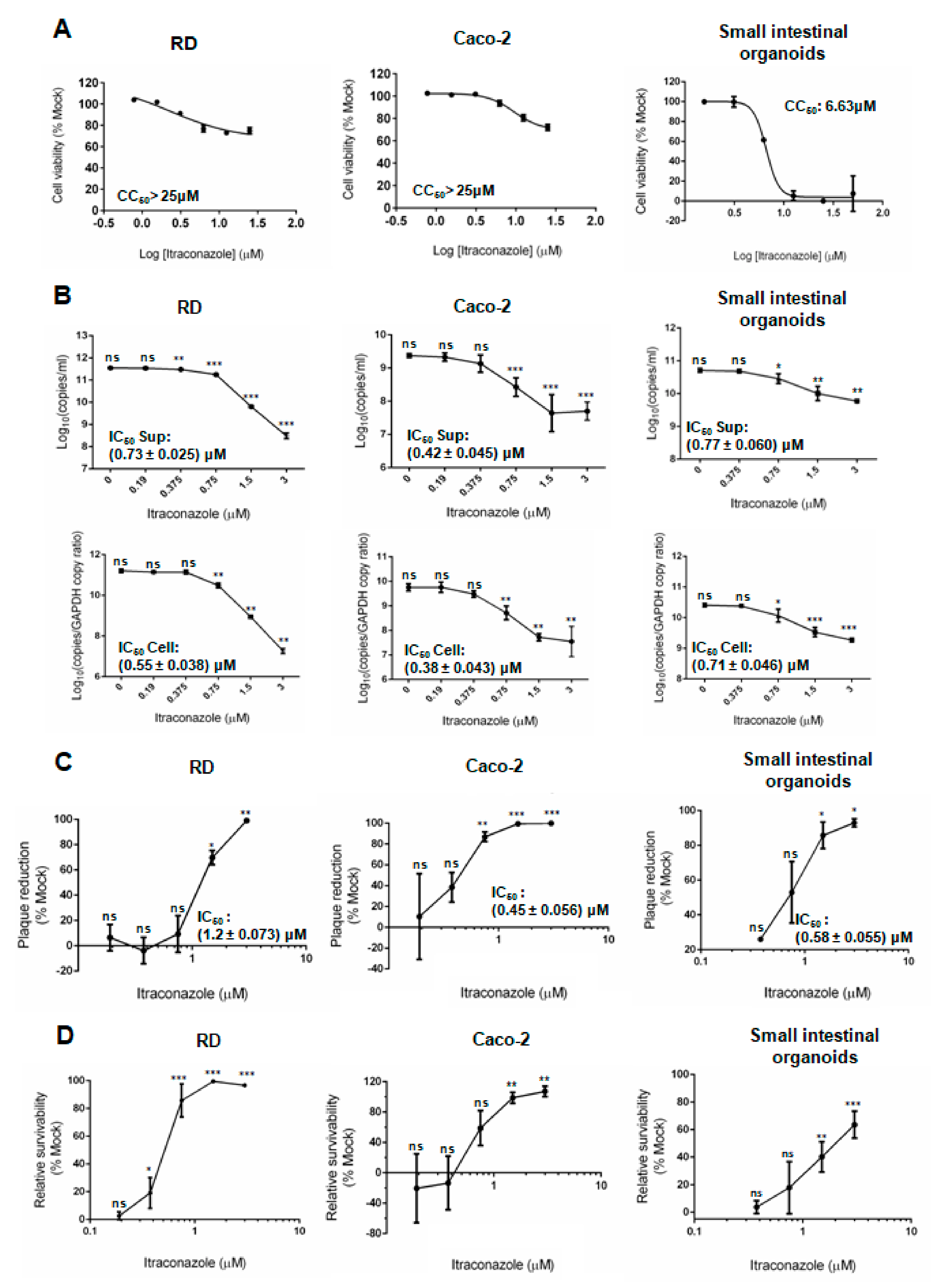
3.4. Comparative Perofrmances of Antiviral Assays in Human Small Intestinal Organoids and Cell Lines
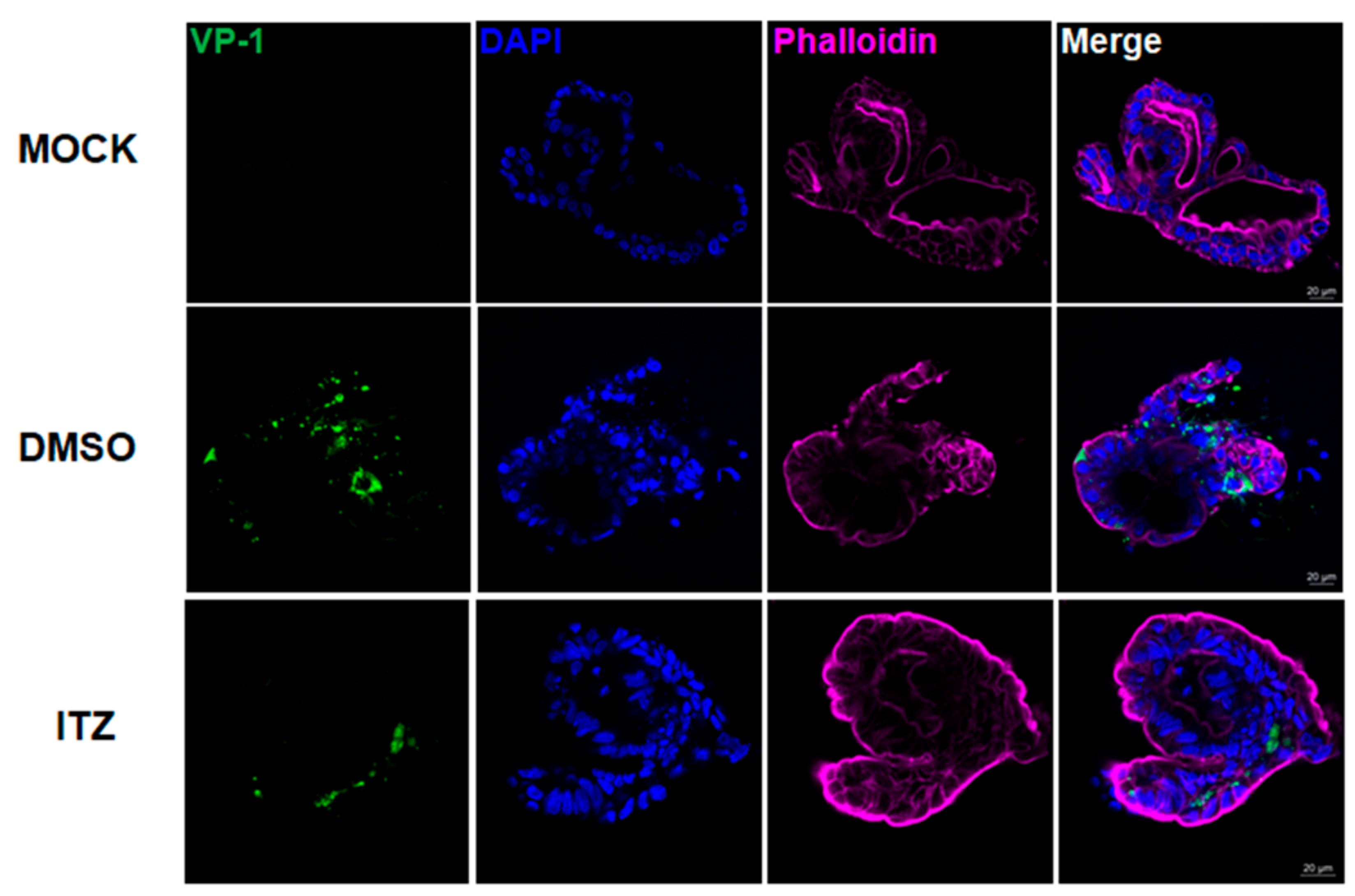
3.5. Immunofluorescence Analysis of ITZ Antiviral and Cell Protection Effects in EV-A71-Infected Human Small Intestinal Organoids
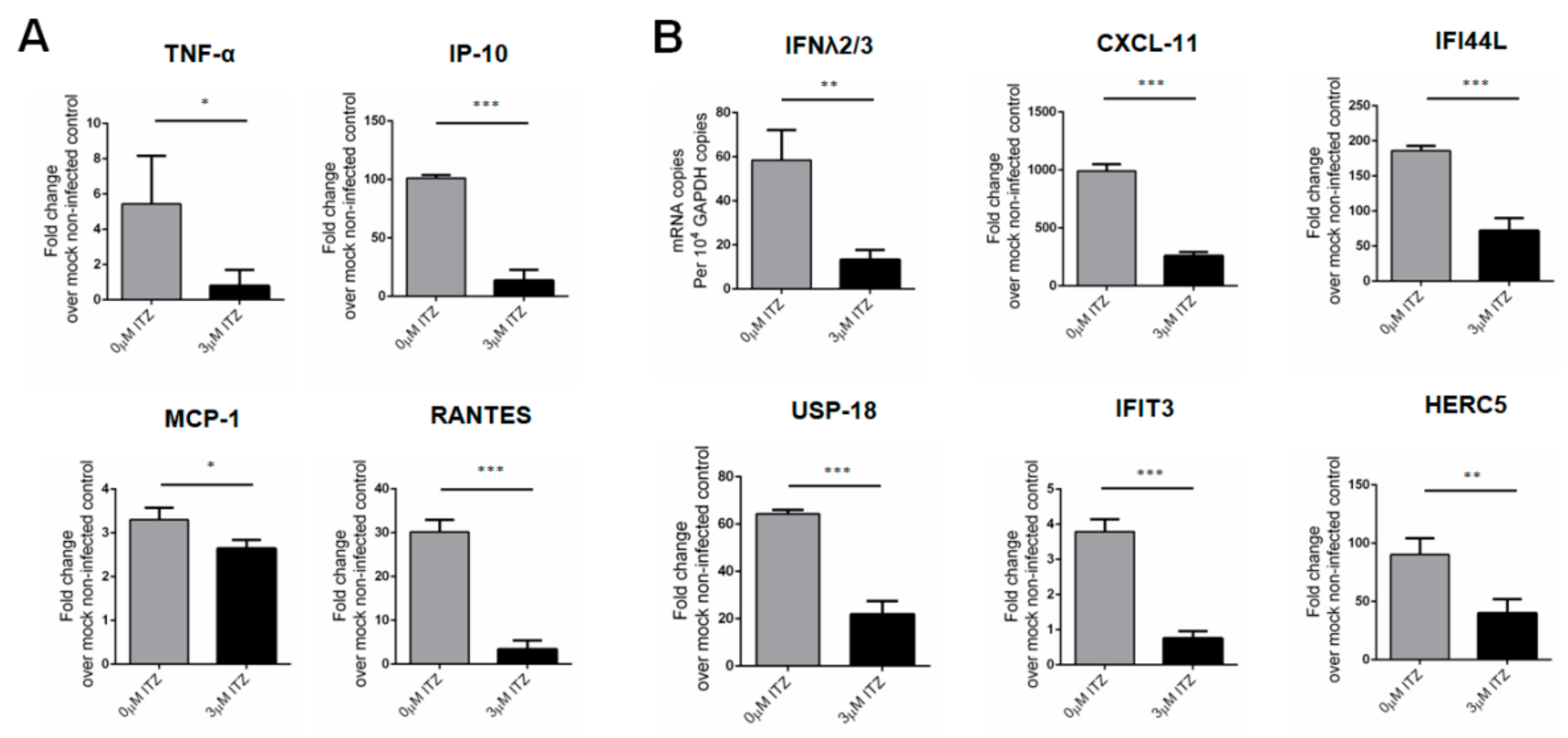
3.6. Characterization of Host Response Changes after ITZ Treatment in EV-A71-Infected Human Small Intestinal Organoids
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagents | Suppliers | Working Concentration |
|---|---|---|
| Advanced DMEM–F-12 medium | Invitrogen | N/A |
| Penicillin-Streptomycin | Invitrogen | 1% |
| HEPES buffer | Invitrogen | 10 mM |
| GlutaMAX | Invitrogen | 1X |
| Epidermal growth factor (EGF) | Invitrogen | 50 ng/mL |
| Noggin-conditioned medium # | N/A | 10% |
| R-spondin-1-conditioned medium # | N/A | 20% |
| Wnt3A-conditioned medium # | N/A | 50% |
| Nicotinamide | Sigma-Aldrich | 10 mM |
| Gastrin I | Sigma-Aldrich | 10 nM |
| A-83-01 | Tocris Bioscience | 500 nM |
| SB202190 | Sigma-Aldrich | 10 μM |
| B27 supplement | Invitrogen | 1X |
| N2 supplement | Invitrogen | 1X |
| N-acetylcysteine | Sigma-Aldrich | 1 mM |
| Species Group | Geographic Origin | Illness | GenBank Accession Number |
|---|---|---|---|
| EV-A71 | Malaysia | Hand, foot and mouth disease | DQ341368.1 |
| CVB2 | Ohio | Flu-like symptoms | AF085363 |
| PV-3 | California | Fatal paralytic poliomyelitis | K01392 |
| EV-D68 | California | Lower respiratory illness | AY426531 |
| Genes | Primer Sequence | |
|---|---|---|
| PanEV | F | 5′- GCCCCTGAATGCGGCTAAT -3′ |
| R | 5′- ATTGTCACCATAAGCAGCCA -3′ | |
| Probe | 5′- 6FAM-CGGACACCCAAAGTAGTCGGTTCCG –lABkFQ -3′ | |
| hGAPDH | F | 5′- ATTCCACCCATGGCAAATTC -3′ |
| R | 5′- CGCTCCTGGAAGATGGTGAT -3′ | |
| SCARB2 | F | 5′- GCTGGGTGTGTTCTTTGGTTTG -3′ |
| R | 5′- TTTCATCCGCTGTTCCCTCATC -3′ | |
| PSGL-1 | F | 5′- TGCTGCTCCTCTGACTT -3′ |
| R | 5′- CCACACAGTCCCAAAGAA -3′ | |
| CAR | F | 5′- TCATTCATCCCTGGGGTCCA -3′ |
| R | 5′- TGAGGAGTGCGTTCAAAGTCT -3′ | |
| DAF | F | 5′- TAAGGCTGTTTGAGGGATGC -3′ |
| R | 5′- AATTCCTGGCGAGAAGGACT -3′ | |
| Neonatal Fc receptor | F | 5′- AAGAAGAACCACAAGCGTTTTGA -3′ |
| R | 5′- GACCCCAGTTCCTCTCTCACA -3′ | |
| PVR | F | 5′- TGGACGGCAAGAATGTGACC -3′ |
| R | 5′- ATCATAGCCAGAGATGGATACC -3′ | |
| ICAM-5 | F | 5′- CAAGCTCTGGTCACACTGGA -3′ |
| R | 5′- GGGAGCGTATAGGACACGAA -3′ | |
| IP-10 | F | 5′- GAAATTATTCCTGCAAGCCAATTT -3′ |
| R | 5′- TCACCCTTCTTTTTCATTGTAGCA -3′ | |
| MCP-1 | F | 5′- AGATCTGTGCTGACCCCAAG -3′ |
| R | 5′- GGAGTTTGGGTTTGCTTGTCC -3′ | |
| RANTES | F | 5′- ACCACACCCTGCTGCTTTGC -3′ |
| R | 5′- CCGAACCCATTTCTTCTCTGG -3′ | |
| TNF-α | F | 5′- GGCTCCAGGCGGTGCTTGTTC -3′ |
| R | 5′- AGACGGCGATGCGGCTGATG -3′ | |
| IFNλ 2/3 | F | 5′- CCTGACGCTGAAGGTTCTGG -3′ |
| R | 5′- ATATGGTGCAGGGTGTGAAGG -3′ | |
| CXCL11 | F | 5′- ATGAGTGTGAAGGGCATGGC -3′ |
| R | 5′- TCACTGCTTTTACCCCAGGG -3′ | |
| IFI44L | F | 5′- AACCTAGACGACATAAAGAGG -3′ |
| R | 5′- CTGAAACCAAGTCTGCATAG -3′ | |
| USP18 | F | 5′- GACTCCTTGATTTGCGTTG -3′ |
| R | 5′- TTGCTTGATAACTCCCTGG -3′ | |
| IFIT3 | F | 5′- TTCACCTGGAACTTATTCAAG -3′ |
| R | 5′- TTTTATGTAGGCCAACAAGTT -3′ | |
| HERC5 | F | 5′- CAGAAAGTTGAATTTGTCGC -3′ |
| R | 5′- CTGAGTCACTCTATACCCAAC -3′ | |
References
- International Committee on Taxonomy of Viruses. Genus: Enterovirus [Internet]. 2019. Available online: https://talk.ictvonline.org/ictv-reports/ictv_online_report/positive-sense-rna-viruses/w/picornaviridae/681/genus-enterovirus (accessed on 2 December 2020).
- Huang, H.-I.; Lin, J.-Y.; Chiang, H.-C.; Huang, P.-N.; Lin, Q.-D.; Shih, S.-R. Exosomes Facilitate Transmission of Enterovirus A71 From Human Intestinal Epithelial Cells. J. Infect Dis. 2020, 222, 456–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, S.S.Y.; Yip, C.C.Y.; Lau, S.K.P.; Yuen, K.Y. Human enterovirus 71 and hand, foot and mouth disease. Epidemiol. Infect. 2010, 138, 1071–1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.Y. Enterovirus 71 infection and neurological complications. Korean J. Pediatr. 2016, 59, 395–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxena, K.; Blutt, S.E.; Ettayebi, K.; Zeng, X.-L.; Broughman, J.R.; Crawford, S.E.; Karandikar, U.C.; Sastri, N.P.; Conner, M.E.; Opekun, A.R.; et al. Human Intestinal Enteroids: A New Model to Study Human Rotavirus Infection, Host Restriction, and Pathophysiology. J. Virol. 2016, 90, 43–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finkbeiner, S.R.; Zeng, X.L.; Utama, B.; Atmar, R.L.; Shroyer, N.F.; Estesa, M.K. Stem cell-derived human intestinal organoids as an infection model for rotaviruses. MBio 2012, 3, e00159-12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ettayebi, K.; Crawford, S.E.; Murakami, K.; Broughman, J.R.; Karandikar, U.; Tenge, V.R.; Neill, F.H.; Blutt, S.E.; Zeng, X.L.; Qu, L.; et al. Replication of human noroviruses in stem cell-derived human enteroids. Science 2016, 353, 1387–1393. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Li, C.; Zhao, G.; Chu, H.; Wang, D.; Yan, H.H.-N.; Poon, V.K.-M.; Wen, L.; Wong, B.H.-Y.; Zhao, X.; et al. Human intestinal tract serves as an alternative infection route for Middle East respiratory syndrome coronavirus. Sci. Adv. 2017, 3, eaao4966. [Google Scholar] [CrossRef] [Green Version]
- Tamhankar, M.; Patterson, J.L. Directional entry and release of Zika virus from polarized epithelial cells. Virol. J. 2019, 16, 99. [Google Scholar] [CrossRef] [Green Version]
- Mee, C.J.; Grove, J.; Harris, H.J.; Hu, K.; Balfe, P.; McKeating, J.A. Effect of Cell Polarization on Hepatitis C Virus Entry. J. Virol. 2008, 82, 461–470. [Google Scholar] [CrossRef] [Green Version]
- Drummond, C.G.; Bolock, A.M.; Ma, C.; Luke, C.J.; Good, M.; Coyne, C.B. Enteroviruses infect human enteroids and induce antiviral signaling in a cell lineage-specific manner. Proc. Natl. Acad. Sci. USA 2017, 114, 1672–1677. [Google Scholar] [CrossRef] [Green Version]
- Good, C.; Wells, A.I.; Coyne, C.B. Type III interferon signaling restricts enterovirus 71 infection of goblet cells. Sci. Adv. 2019, 5, eaau4255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Q.; Yuan, S.; Zhang, C.; Wang, Y.; Wang, Y.; He, G.; Zhang, S.; Altmeyer, R.; Zou, G. Discovery of itraconazole with broad-spectrum in vitro antienterovirus activity that targets nonstructural protein 3A. Antimicrob Agents Chemother. 2015, 59, 2654–2665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strating, J.R.P.M.; van der Linden, L.; Albulescu, L.; Bigay, J.; Arita, M.; Delang, L.; Leyssen, P.; van der Schaar, H.M.; Lanke, K.H.W.; jan Thibaut, H.; et al. Itraconazole inhibits enterovirus replication by targeting the oxysterol-binding protein. Cell Rep. 2015, 10, 600–615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Li, G.; Yuan, S.; Gao, Q.; Lan, K.; Altmeyer, R.; Zou, G. In vitro assessment of combinations of enterovirus inhibitors against enterovirus 71. Antimicrob Agents Chemother. 2016, 60, 5357–5367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, T.; Stange, D.E.; Ferrante, M.; Vries, R.G.J.; van Es, J.H.; van den Brink, S.; van Houdt, W.J.; Pronk, A.; Gorp, J.; Siersema, P.D.; et al. Long-term expansion of epithelial organoids from human colon, adenoma, adenocarcinoma, and Barrett’s epithelium. Gastroenterology 2011, 141, 1762–1772. [Google Scholar] [CrossRef] [PubMed]
- Yeung, M.L.; Jia, L.; Yip, C.C.Y.; Chan, J.F.W.; Teng, J.L.L.; Chan, K.H.; Cai, J.; Zhang, C.; Zhang, A.J.; Wong, W.; et al. Human tryptophanyl-tRNA synthetase is an IFN-γ-inducible entry factor for Enterovirus. J. Clin. Investig. 2018, 128, 5163–5177. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.; Chan, J.F.-W.; Yuen, T.T.-T.; Shuai, H.; Yuan, S.; Wang, Y.; Hu, B.; Yip, C.C.; Tsang, J.O.; Huang, X.; et al. Comparative tropism, replication kinetics, and cell damage profiling of SARS-CoV-2 and SARS-CoV with implications for clinical manifestations, transmissibility, and laboratory studies of COVID-19: An observational study. Lancet Microbe. 2020, 1, e14–e23. [Google Scholar] [CrossRef]
- Chan, J.F.W.; Chik, K.K.H.; Yuan, S.; Yip, C.C.Y.; Zhu, Z.; Tee, K.M.; Tsang, J.O.; Chung-Sing, C.; Vincent, C.; Poon, K.; et al. Novel antiviral activity and mechanism of bromocriptine as a Zika virus NS2B-NS3 protease inhibitor. Antiviral Res. 2017, 141, 29–37. [Google Scholar] [CrossRef]
- Yuan, S.; Chan, J.F.-W.; Den-Haan, H.; Chik, K.K.-H.; Zhang, A.J.; Chan, C.C.-S.; Poon, V.K.-M.; Yip, C.C.Y.; Mak, W.W.-N.; Zhu, Z.; et al. Structure-based discovery of clinically approved drugs as Zika virus NS2B-NS3 protease inhibitors that potently inhibit Zika virus infection in vitro and in vivo. Antiviral Res. 2017, 145, 33–43. [Google Scholar] [CrossRef]
- Yuan, S.; Chu, H.; Chan, J.F.W.; Ye, Z.W.; Wen, L.; Yan, B.; Lai, P.; Tee, K.; Huang, J.; Che, D.; et al. SREBP-dependent lipidomic reprogramming as a broad-spectrum antiviral target. Nat. Commun. 2019, 10, 120. [Google Scholar] [CrossRef]
- Yuan, S.; Chu, H.; Huang, J.; Zhao, X.; Ye, Z.W.; Lai, P.M.; Wen, L.; Cai, J.P.; Mo, Y.; Cao, J.; et al. Viruses harness YxxØ motif to interact with host AP2M1 for replication: A vulnerable broad-spectrum antiviral target. Sci. Adv. 2020, 6, eaba7910. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.; Tsang, J.O.L.; Yan, B.; Chik, K.K.H.; Chan, C.C.Y.; Cao, J.; Liang, R.; Tang, K.; Yin, F.; Ye, Z.-W.; et al. Metabolic profiling reveals significant perturbations of intracellular glucose homeostasis in enterovirus-infected cells. Metabolites 2020, 10, 302. [Google Scholar] [CrossRef] [PubMed]
- Yan, B.; Zou, Z.; Chu, H.; Chan, G.; Tsang, J.O.L.; Lai, P.M.; Yuan, S.; Yip, C.C.-Y.; Yin, F.; Kao, R.Y.; et al. Lipidomic profiling reveals significant perturbations of intracellular lipid homeostasis in enterovirus-infected cells. Int. J. Mol. Sci. 2019, 20, 5952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, J.F.W.; Yip, C.C.Y.; Tsang, J.O.L.; Tee, K.M.; Cai, J.P.; Chik, K.K.H.; Zhu, Z.; Chan, C.C.; Choi, G.K.; Sridhar, S.; et al. Differential cell line susceptibility to the emerging Zika virus: Implications for disease pathogenesis, non-vector-borne human transmission and animal reservoirs. Emerg. Microbes Infect. 2016, 5, e93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamayoshi, S.; Yamashita, Y.; Li, J.; Hanagata, N.; Minowa, T.; Takemura, T.; Koike, S. Scavenger receptor B2 is a cellular receptor for enterovirus 71. Nat. Med. 2009, 15, 798–801. [Google Scholar] [CrossRef]
- Nishimura, Y.; Shimojima, M.; Tano, Y.; Miyamura, T.; Wakita, T.; Shimizu, H. Human P-selectin glycoprotein ligand-1 is a functional receptor for enterovirus 71. Nat. Med. 2009, 15, 794–797. [Google Scholar] [CrossRef]
- Bergelson, J.M.; Chan, M.; Solomon, K.R.; St. John, N.F.; Lin, H.; Finberg, R.W. Decay-accelerating factor (CD55), a glycosylphosphatidylinositol-anchored complement regulatory protein, is a receptor for several echoviruses. Proc. Natl. Acad. Sci. USA 1994, 91, 6245–6248. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Zhang, G.; Liu, S.; Chen, X.; Peng, R.; Dai, L.; Qu, X.; Li, S.; Song, H.; Gao, Z.; et al. Human Neonatal Fc Receptor Is the Cellular Uncoating Receptor for Enterovirus, B. Cell 2019, 177, 1553–1565. [Google Scholar] [CrossRef]
- Bergelson, J.M.; Cunningham, J.A.; Droguett, G.; Kurt-Jones, E.A.; Krithivas, A.; Hong, J.S.; Horwitz, M.S.; Crowell, R.L.; Finberg, R.W. Isolation of a common receptor for coxsackie B viruses and adenoviruses 2 and 5. Science 1997, 275, 1320–1323. [Google Scholar] [CrossRef]
- Kaplan, G.; Freistadt, M.S.; Racaniello, V.R. Neutralization of poliovirus by cell receptors expressed in insect cells. J. Virol. 1990, 64, 4697–4702. [Google Scholar] [CrossRef] [Green Version]
- Wei, W.; Guo, H.; Chang, J.; Yu, Y.; Liu, G.; Zhang, N.; Willard, S.H.; Zheng, S.; Yu, X.-F. ICAM-5/Telencephalin Is a Functional Entry Receptor for Enterovirus D68. Cell Host Microbe 2016, 20, 631–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, W.; Qian, S.; Fang, L.; Han, Y.; Zheng, C. Association study of inflammatory cytokine and chemokine expression in hand foot and mouth disease. Oncotarget 2017, 8, 79425–79432. [Google Scholar] [CrossRef] [PubMed]
- Messacar, K.; Abzug, M.J.; Dominguez, S.R. The Emergence of Enterovirus-D68. Emerg. Infect. 2016, 10, 4. [Google Scholar]
- Oberste, M.S.; Maher, K.; Schnurr, D.; Flemister, M.R.; Lovchik, J.C.; Peters, H.; Sessions, W.; Kirk, C.; Chatterjee, N.; Fuller, S.; et al. Enterovirus 68 is associated with respiratory illness and shares biological features with both the enteroviruses and the rhinoviruses. J. Gen. Virol. 2004, 85, 2577–2584. [Google Scholar] [CrossRef]
- Rausell, A.; Muñoz, M.; Martinez, R.; Roger, T.; Telenti, A.; Ciuffi, A. Innate immune defects in HIV permissive cell lines. Retrovirology 2016, 13, 43. [Google Scholar] [CrossRef] [Green Version]
- Kolawole, A.O.; Mirabelli, C.; Hill, D.R.; Svoboda, S.A.; Janowski, A.B.; Passalacqua, K.D.; Rodriguez, B.N.; Dame, M.K.; Freiden, P.; Berger, R.P.; et al. Astrovirus replication in human intestinal enteroids reveals multi-cellular tropism and an intricate host innate immune landscape. PLoS Pathog. 2019, 15, e1008057. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.M.; Lei, H.Y.; Liu, C.C. Cytokine immunopathogenesis of enterovirus 71 brain stem encephalitis. Clin. Dev. Immunol. 2012, 2012, 876241. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Lei, H.; Huang, K.; Wu, J.; Wang, J.; Yu, C.; Su, I.-J.; Liu, C.C. Pathogenesis of Enterovirus 71 Brainstem Encephalitis in Pediatric Patients: Roles of Cytokines and Cellular Immune Activation in Patients with Pulmonary Edema. J. Infect Dis. 2003, 188, 564–570. [Google Scholar] [CrossRef] [Green Version]
- Wieczorek, M.; Ciąćka, A.; Witek, A.; Kuryk, Ł.; Żuk-Wasek, A. Environmental Surveillance of Non-polio Enteroviruses in Poland, 2011. Food Environ. Virol. 2015, 7, 224–231. [Google Scholar] [CrossRef]
- Kuryk, L.; Wieczorek, M.; Diedrich, S.; Böttcher, S.; Witek, A.; Litwińska, B. Genetic analysis of poliovirus strains isolated from sewage in Poland. J. Med. Virol. 2014, 86, 1243–1248. [Google Scholar] [CrossRef]






| CC50 (μM) | IC50 (μM) | Selectivity Index | |
|---|---|---|---|
| Human small intestinal organoids | 6.63 | 0.58 ± 0.055 | 11.43 |
| RD | >25 | 1.20 ± 0.073 | >20.83 |
| Caco-2 | >25 | 0.45 ± 0.056 | >55.56 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsang, J.O.-L.; Zhou, J.; Zhao, X.; Li, C.; Zou, Z.; Yin, F.; Yuan, S.; Yeung, M.-L.; Chu, H.; Chan, J.F.-W. Development of Three-Dimensional Human Intestinal Organoids as a Physiologically Relevant Model for Characterizing the Viral Replication Kinetics and Antiviral Susceptibility of Enteroviruses. Biomedicines 2021, 9, 88. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9010088
Tsang JO-L, Zhou J, Zhao X, Li C, Zou Z, Yin F, Yuan S, Yeung M-L, Chu H, Chan JF-W. Development of Three-Dimensional Human Intestinal Organoids as a Physiologically Relevant Model for Characterizing the Viral Replication Kinetics and Antiviral Susceptibility of Enteroviruses. Biomedicines. 2021; 9(1):88. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9010088
Chicago/Turabian StyleTsang, Jessica Oi-Ling, Jie Zhou, Xiaoyu Zhao, Cun Li, Zijiao Zou, Feifei Yin, Shuofeng Yuan, Man-Lung Yeung, Hin Chu, and Jasper Fuk-Woo Chan. 2021. "Development of Three-Dimensional Human Intestinal Organoids as a Physiologically Relevant Model for Characterizing the Viral Replication Kinetics and Antiviral Susceptibility of Enteroviruses" Biomedicines 9, no. 1: 88. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9010088