
Distinct Roles of Vav Family Members in Adaptive and Innate Immune Models of Arthritis

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Antigen-Induced Arthritis
2.3. Histological Determinations
2.4. Cell Isolation
2.5. Flow Cytometry
2.6. Quantitative RT-PCR
2.7. T and B Cell Activity Assays
2.8. Zymosan A-Induced Arthritis
2.9. Statistical Analyses
3. Results
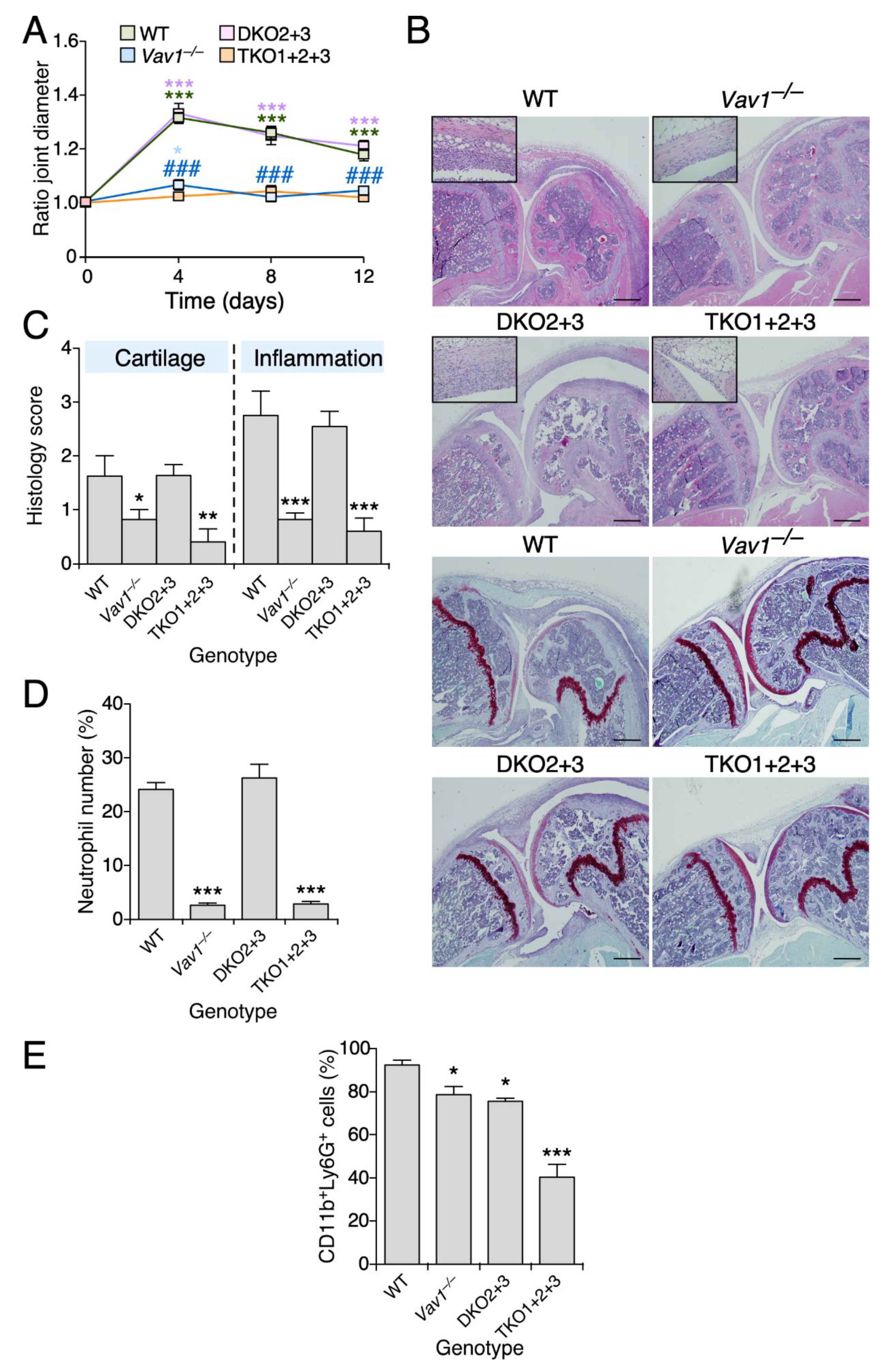
3.1. Vav1 Deficiency Decreases the Severity of Antigen-Induced Arthritis
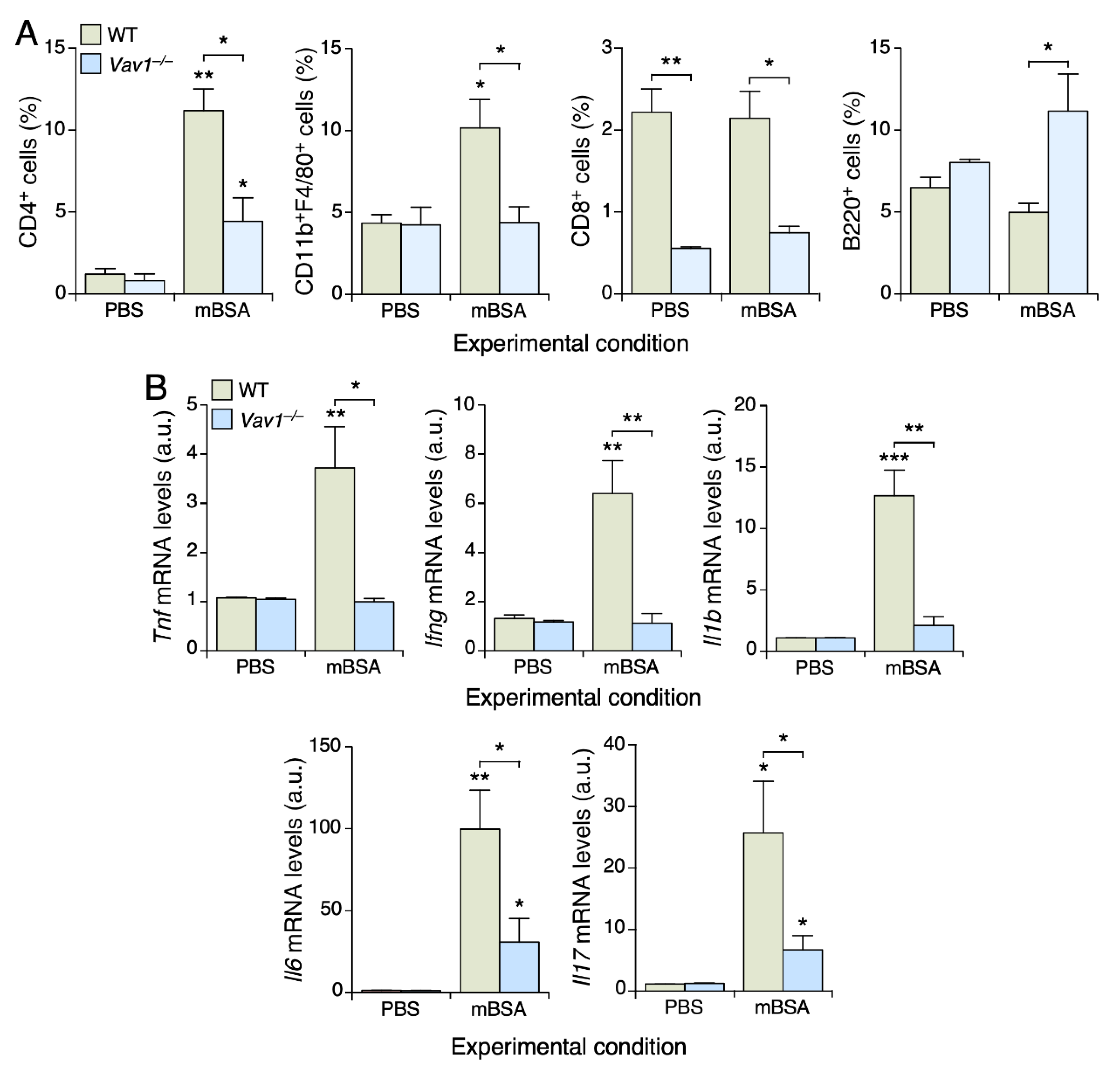
3.2. Vav1 Deficiency Impairs the Infiltration of CD4+ T Cells and Macrophages in mBSA-Treated Joints
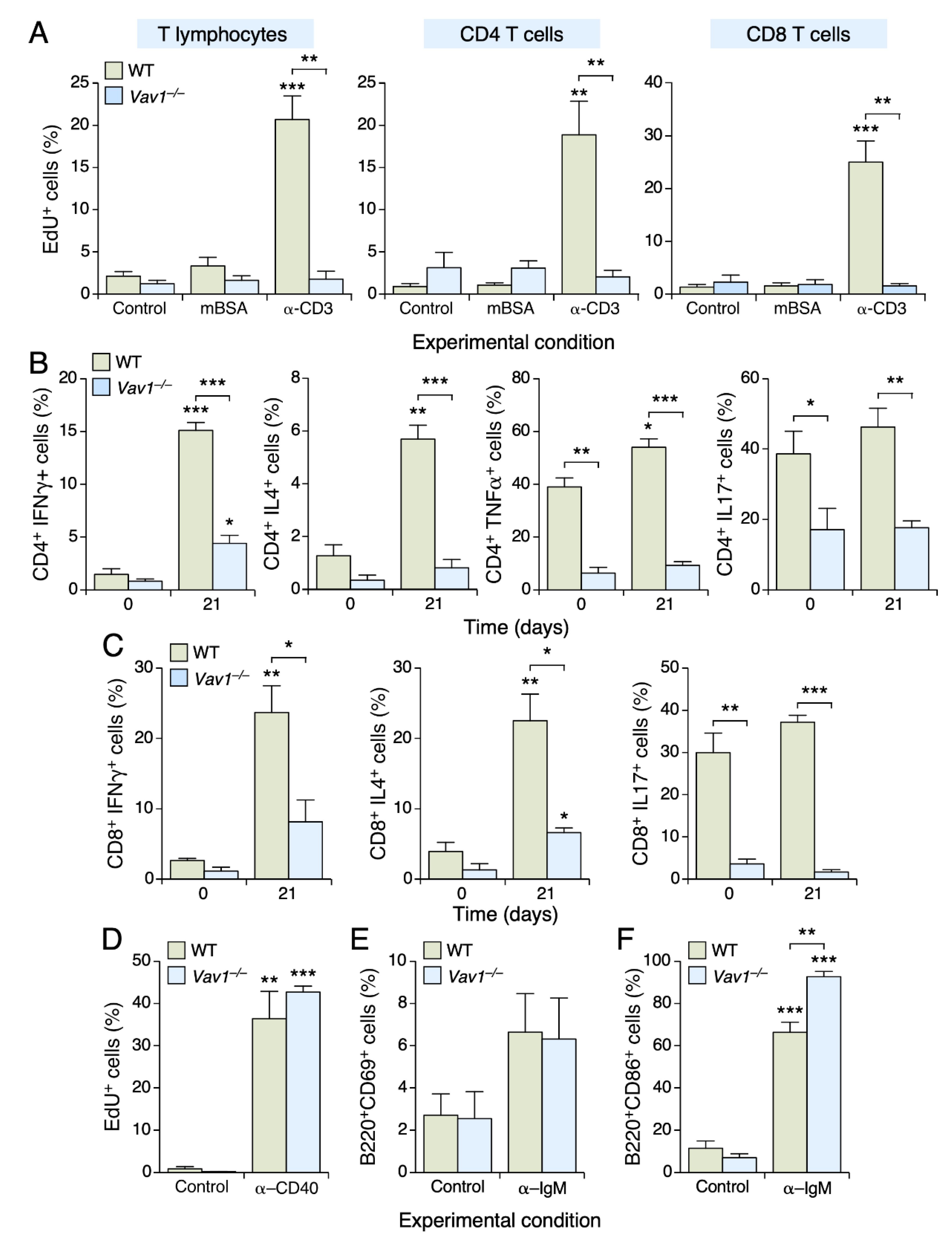
3.3. The Vav1 Deficiency Impairs T Cell Proliferation and Differentiation
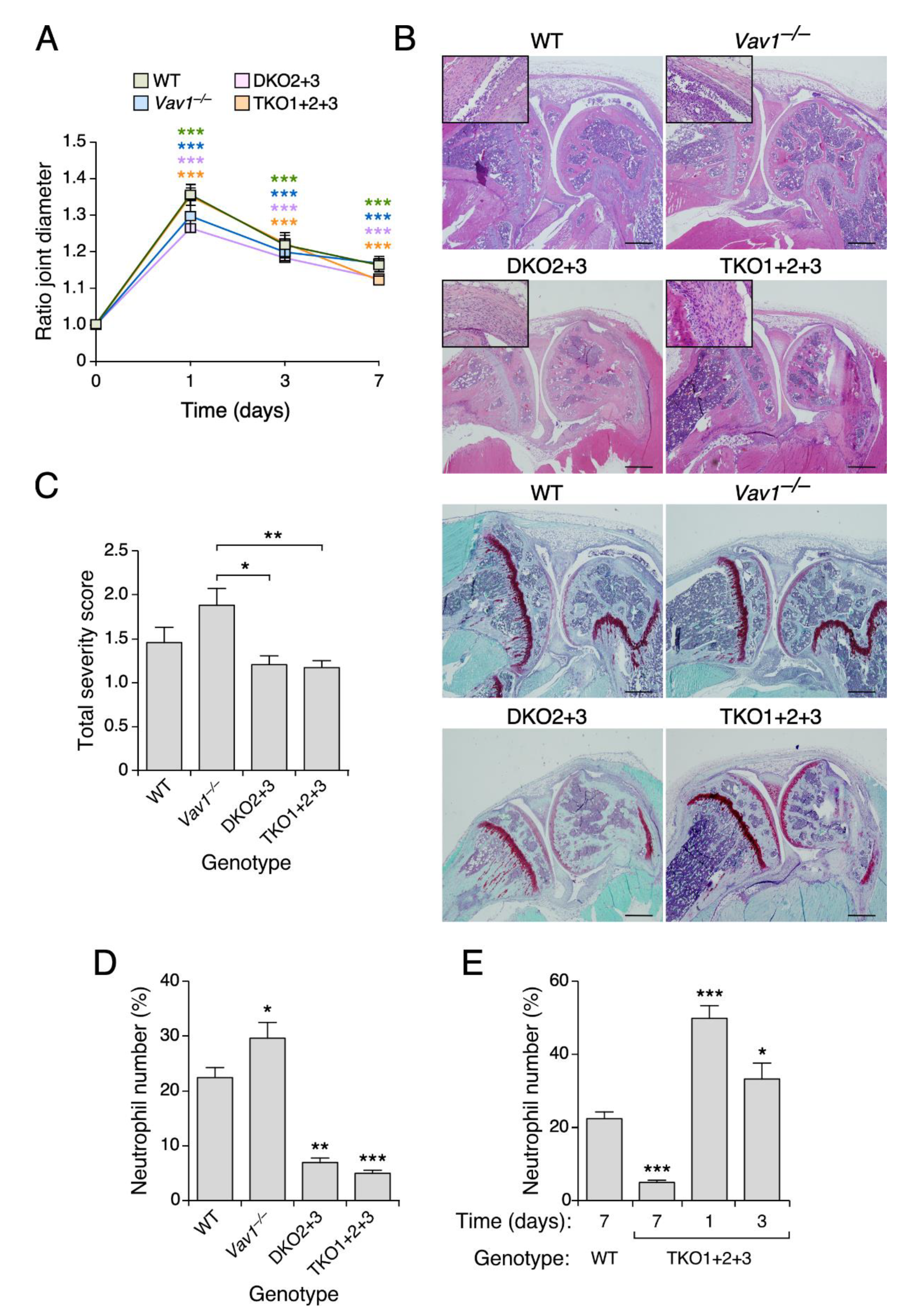
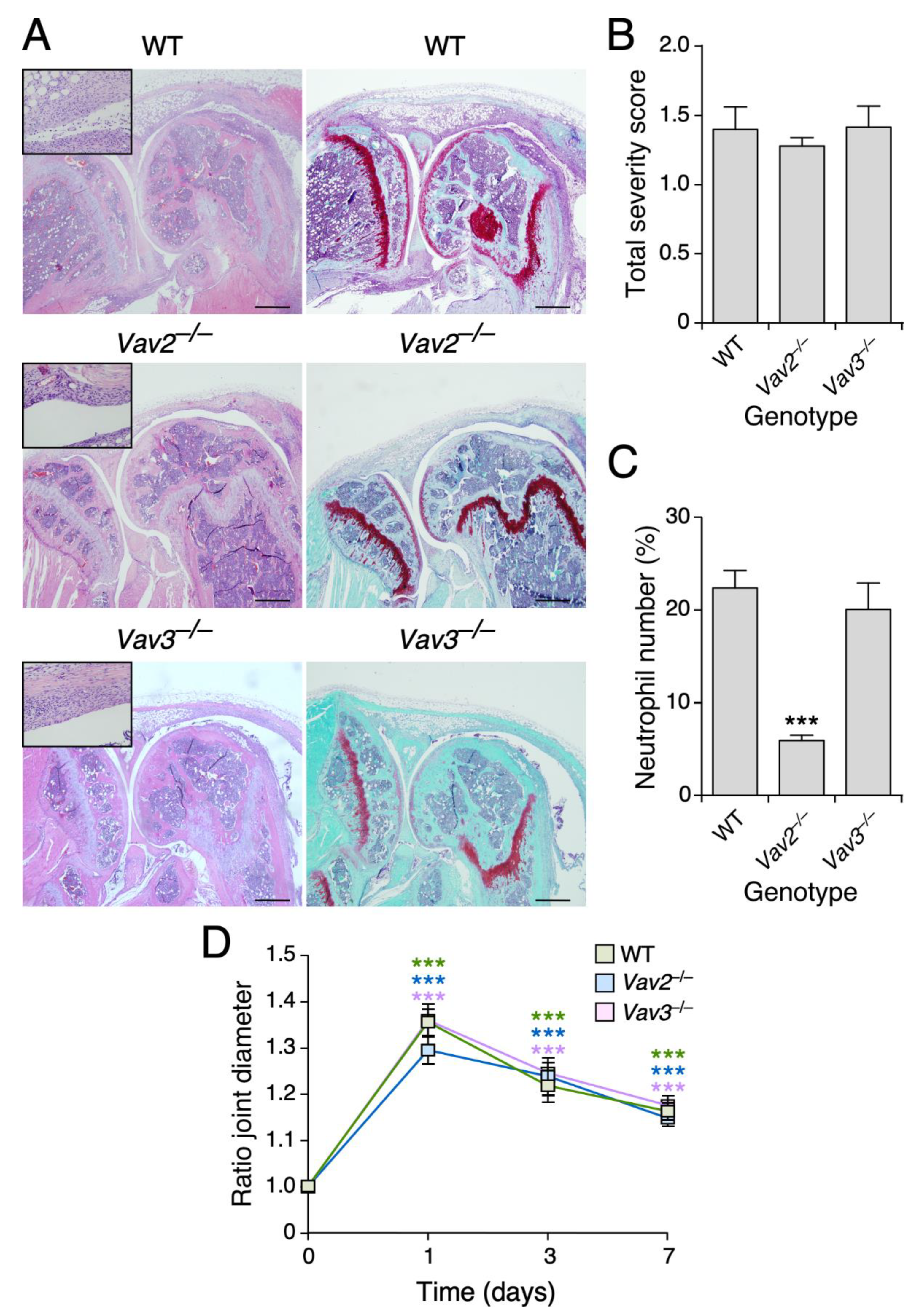
3.4. Vav2, but Not the Rest of Vav Family Proteins, Plays Roles in Zymosan A-Induced Arthritis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rodríguez-Fdez, S.; Bustelo, X.R. The Vav GEF Family: An Evolutionary and Functional Perspective. Cells 2019, 8, 465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustelo, X.R.; Dosil, M. The Vav family. In Encyclopedia of Signaling Molecules; Choi, S., Ed.; Springer: New York, NY, USA, 2016; pp. 1–15. [Google Scholar]
- Bustelo, X.R. Vav family exchange factors: An integrated regulatory and functional view. Small GTPases 2014, 5, e973757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katzav, S.; Martin-Zanca, D.; Barbacid, M. vav, a novel human oncogene derived from a locus ubiquitously expressed in hematopoietic cells. EMBO J. 1989, 8, 2283–2290. [Google Scholar] [CrossRef]
- Bustelo, X.R.; Rubin, S.D.; Suen, K.L.; Carrasco, D.; Barbacid, M. Developmental expression of the vav protooncogene. Cell Growth Differ. Mol. Boil. J. Am. Assoc. Cancer Res. 1993, 4, 297–308. [Google Scholar]
- Schuebel, K.E.; Bustelo, X.R.; Nielsen, D.A.; Song, B.J.; Barbacid, M.; Goldman, D.; Lee, I.J. Isolation and characterization of murine vav2, a member of the vav family of proto-oncogenes. Oncogene 1996, 13, 363–371. [Google Scholar]
- Movilla, N.; Bustelo, X.R. Biological and Regulatory Properties of Vav-3, a New Member of the Vav Family of Oncoproteins. Mol. Cell. Biol. 1999, 19, 7870–7885. [Google Scholar] [CrossRef] [Green Version]
- Crespo, P.; Schuebel, K.E.; Ostrom, A.A.; Gutkind, J.S.; Bustelo, X.R. Phosphotyrosine-dependent activation of Rac-1 GDP/GTP exchange by the vav proto-oncogene product. Nat. Cell Biol. 1997, 385, 169–172. [Google Scholar] [CrossRef]
- Schuebel, K.E.; Movilla, N.; Rosa, J.L.; Bustelo, X.R. Phosphorylation-dependent and constitutive activation of Rho proteins by wild-type and oncogenic Vav-2. EMBO J. 1998, 17, 6608–6621. [Google Scholar] [CrossRef] [Green Version]
- Pawlik, A.; Malinowski, D.; Paradowska-Gorycka, A.; Safranow, K.; Dziedziejko, V. Gene Polymorphisms in Patients with Rheumatoid Arthritis. Int. J. Environ. Res. Public Health 2020, 5, 3214. [Google Scholar] [CrossRef]
- Guerreiro-Cacais, A.O.; Rheumatoid Arthritis Consortium International (RACI); Norin, U.; Gyllenberg, A.; Berglund, R.; Beyeen, A.D.; Petit-Teixeira, E.; Cornélis, F.; Saoudi, A.; Fournié, G.J.; et al. VAV1 regulates experimental autoimmune arthritis and is associated with anti-CCP negative rheumatoid arthritis. Genes Immun. 2017, 18, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Jagodic, M.; Colacios, C.; Nohra, R.; Dejean, A.S.; Beyeen, A.D.; Khademi, M.; Casemayou, A.; Lamouroux, L.; Duthoit, C.; Papapietro, O.; et al. A Role for VAV1 in Experimental Autoimmune Encephalomyelitis and Multiple Sclerosis. Sci. Transl. Med. 2009, 1, 10ra21. [Google Scholar] [CrossRef]
- Cremasco, V.; Graham, D.B.; Novack, D.V.; Swat, W.; Faccio, R. Vav/Phospholipase Cgamma2-mediated control of a neutrophil-dependent murine model of rheumatoid arthritis. Arthritis Rheum. 2008, 58, 2712–2722. [Google Scholar] [CrossRef] [Green Version]
- Fabbiano, S.; Menacho-Márquez, M.; Robles-Valero, J.; Pericacho, M.; Matesanz-Marín, A.; García-Macías, C.; Sevilla, M.A.; Montero, M.J.; Alarcón, B.; López-Novoa, J.M.; et al. Immunosuppression-Independent Role of Regulatory T Cells against Hypertension-Driven Renal Dysfunctions. Mol. Cell. Biol. 2015, 35, 3528–3546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauzeau, V.; Jerkic, M.; Lopez-Novoa, J.M.; Bustelo, X.R. Loss of Vav2 Proto-Oncogene Causes Tachycardia and Cardiovascular Disease in Mice. Mol. Biol. Cell 2007, 18, 943–952. [Google Scholar] [CrossRef] [Green Version]
- Sauzeau, V.; Sevilla, M.A.; Rivas-Elena, J.V.; de Alava, E.; Montero, M.J.; Lopez-Novoa, J.M.; Bustelo, X.R. Vav3 proto-oncogene deficiency leads to sympathetic hyperactivity and cardiovascular dysfunction. Nat. Med. 2006, 12, 841–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, M.; Mee, P.; Walters, A.E.; Quinn, M.E.; Mellor, A.L.; Zamoyska, R.; Tybulewicz, V.L. A Requirement for the Rho-Family GTP Exchange Factor Vav in Positive and Negative Selection of Thymocytes. Immunity 1997, 7, 451–460. [Google Scholar] [CrossRef] [Green Version]
- Bessis, N.; Decker, P.; Assier, E.; Semerano, L.; Boissier, M.-C. Arthritis models: Usefulness and interpretation. Semin. Immunopathol. 2017, 39, 469–486. [Google Scholar] [CrossRef]
- Petrelli, A.; van Wijk, F. CD8(+) T cells in human autoimmune arthritis: The unusual suspects. Nat. Rev. Rheumatol. 2016, 12, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Chemin, K.; Gerstner, C.; Malmström, V. Effector Functions of CD4+ T Cells at the Site of Local Autoimmune Inflammation-Lessons from Rheumatoid Arthritis. Front Immunol. 2019, 10, 353. [Google Scholar] [CrossRef] [Green Version]
- Tybulewicz, V.L. Analysis of antigen receptor signalling using mouse gene targeting. Curr. Opin. Cell Biol. 1998, 10, 195–204. [Google Scholar] [CrossRef]
- Dorner, T.; Lipsky, P.E. B cells: Depletion or functional modulation in rheumatic diseases. Curr. Opin. Rheumatol. 2014, 26, 228–236. [Google Scholar] [CrossRef]
- Frasnelli, M.E.; Tarussio, D.; Chobaz-Péclat, V.; Busso, N.; So, A. TLR2 modulates inflammation in zymosan-induced arthritis in mice. Arthritis Res. 2005, 7, R370–R379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brackertz, D.; Mitchell, G.F.; Mackay, I.R. Antigen-induced arthritis in mice. I. Induction of arthritis in various strains of mice. Arthritis Rheum. 1977, 20, 841–850. [Google Scholar] [CrossRef]
- Tarakhovsky, A.; Turner, M.; Schaal, S.; Mee, P.J.; Duddy, L.P.; Rajewsky, K.; Tybulewicz, V.L.J. Defective antigen receptor-mediated proliferation of B and T cells in the absence of Vav. Nat. Cell Biol. 1995, 374, 467–470. [Google Scholar] [CrossRef] [PubMed]
- Tybulewicz, V.L.J.; Ardouin-Bataille, L.; Prisco, A.; Reynolds, L.F. Vav1: A key signal transducer downstream of the TCR. Immunol. Rev. 2003, 192, 42–52. [Google Scholar] [CrossRef]
- Fujikawa, K.; Miletic, A.V.; Alt, F.W.; Faccio, R.; Brown, T.; Hoog, J.; Swat, W. Vav1/2/3-null mice define an essential role for Vav family proteins in lymphocyte development and activation but a differential requirement in MAPK signaling in T and B cells. J. Exp. Med. 2003, 198, 1595–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Underhill, D.M. Macrophage recognition of zymosan particles. J Endotoxin Res. 2003, 9, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Underhill, D.M.; Ozinsky, A.; Hajjar, A.; Stevens, A.; Wilson, C.B.; Bassetti, M.; Aderem, A. The Toll-like receptor 2 is recruited to macrophage phagosomes and discriminates between pathogens. Nat. Cell Biol. 1999, 401, 811–815. [Google Scholar] [CrossRef]
- Cook, A.D.; Turner, A.L.; Braine, E.L.; Pobjoy, J.; Lenzo, J.C.; Hamilton, J.A. Regulation of systemic and local myeloid cell subpopulations by bone marrow cell-derived granulocyte-macrophage colony-stimulating factor in experimental inflammatory arthritis. Arthritis Rheum. 2011, 63, 2340–2351. [Google Scholar] [CrossRef]
- Wipke, B.T.; Allen, P.M. Essential Role of Neutrophils in the Initiation and Progression of a Murine Model of Rheumatoid Arthritis. J. Immunol. 2001, 167, 1601–1608. [Google Scholar] [CrossRef] [Green Version]
- Tseng, C.C.; Chang, S.J.; Tsai, W.C.; Ou, T.T.; Wu, C.C.; Sung, W.Y.; Yen, J.H. Increased incidence of rheumatoid arthritis in multiple sclerosis: A nationwide cohort study. Medicine 2016, 95, e3999. [Google Scholar] [CrossRef]
- Kassem, S.; Gaud, G.; Bernard, I.; Benamar, M.; Dejean, A.S.; Liblau, R.; Fournié, G.J.; Colacios, C.; Malissen, B.; Saoudi, A. A Natural Variant of the T Cell Receptor-Signaling Molecule Vav1 Reduces Both Effector T Cell Functions and Susceptibility to Neuroinflammation. PLoS Genet. 2016, 12, e1006185. [Google Scholar] [CrossRef] [PubMed]
- McInnes, I.B.; Schett, G. Cytokines in the pathogenesis of rheumatoid arthritis. Nat. Rev. Immunol. 2007, 7, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Saveliev, A.; Vanes, L.; Ksionda, O.; Rapley, J.; Smerdon, S.J.; Rittinger, K.; Tybulewicz, V.L.J. Function of the Nucleotide Exchange Activity of Vav1 in T Cell Development and Activation. Sci. Signal. 2009, 2, ra83. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conde, J.; Fernández-Pisonero, I.; Cuadrado, M.; Abad, A.; Robles-Valero, J.; Bustelo, X.R. Distinct Roles of Vav Family Members in Adaptive and Innate Immune Models of Arthritis. Biomedicines 2021, 9, 695. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9060695
Conde J, Fernández-Pisonero I, Cuadrado M, Abad A, Robles-Valero J, Bustelo XR. Distinct Roles of Vav Family Members in Adaptive and Innate Immune Models of Arthritis. Biomedicines. 2021; 9(6):695. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9060695
Chicago/Turabian StyleConde, Javier, Isabel Fernández-Pisonero, Myriam Cuadrado, Antonio Abad, Javier Robles-Valero, and Xosé R. Bustelo. 2021. "Distinct Roles of Vav Family Members in Adaptive and Innate Immune Models of Arthritis" Biomedicines 9, no. 6: 695. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9060695