
Lentinan Impairs the Early Development of Zebrafish Embryos, Possibly by Disrupting Glucose and Lipid Metabolism


1
School of Public Health, Zhejiang University School of Medicine, Hangzhou 310058, China
2
Hunter Biotechnology, Inc., Hangzhou 310051, China
*
Author to whom correspondence should be addressed.
Processes 2022, 10(1), 120; https://0-doi-org.brum.beds.ac.uk/10.3390/pr10010120
Submission received: 20 December 2021
/
Revised: 5 January 2022
/
Accepted: 5 January 2022
/
Published: 7 January 2022
(This article belongs to the Special Issue Advances of Food Safety and Toxicology in Food Processes and Engineering)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:LNT is the major biologically active substance extracted from Lentinus edodes (L. edodes). Although functional and pharmacological studies have demonstrated that LNT has multiple benefits for animals and humans, the safety assessment is far from sufficient. To evaluate the potential safety risk, larval zebrafish were continuously exposed to varying concentrations of LNT for 120 h. The 96 h LC50 of LNT was determined to be 1228 μg/mL, and morphological defects including short body length, reduced eye and swim bladder sizes and yolk sac edema were observed. In addition, LNT exposure significantly reduced the blood flow velocity and locomotor activity of larval zebrafish. The biochemical parameters were also affected, showing reduced glucose, triglyceride and cholesterol levels in zebrafish larvae after being exposed to LNT. Correspondingly, the genes involved in glucose and lipid metabolism were disrupted. In conclusion, the present study demonstrates the adverse potential of high concentrations of LNT on the development of zebrafish larvae in the early life stage.
1. Introduction
Lentinan (LNT), a neutral β-glucan, consisting of repeated β-(1,6) branched β-(1,3)-glucan units, is the main biologically active compound that is fully extracted from Lentinus edodes (L. edodes) [1,2]. The beneficial effects of LNT and other β-glucans have been widely studied using modern pharmacological and biological strategies, revealing that β-glucans possess multiple functions. The researchers from Japan first demonstrated the anti-tumor activity of LNT in the 1970s [3,4,5]. Further positive biological activities were uncovered, including anti-oxidative activity [6,7], antibacterial activity [8], anti-virus activity [9], anti-inflammation activity [10], reno-protective activity [11], and most importantly, immunomodulatory activity [12,13,14,15]. Interestingly, the biological activities of β -glucans have been shown in both vertebrates, such as humans, chickens, dogs, and fishes [16,17,18,19], and invertebrates, like shrimp and drosophila [20,21].
Because of their functions and taste, β-glucans are widely used as medicine, antimicrobial agents, dietary supplements and nutrition enhancers. For instance, two kinds of β-glucans, LNT and krestin, have been approved as anti-tumor drugs for gastric cancer treatment in Japan [22]. Extensive studies have been conducted to elucidate the mechanism underlying the anti-tumor activity of LNT. Two chief mechanisms have identified: (1) enhancing the immunity against tumors, including cellular and humoral immunities [23,24]; (2) preventing cancer cell proliferation or metastasis through direct induction of cell apoptosis and death [25]. Besides the applications in cancer therapy, LNT has been approved as a safe food ingredient in the European Union and used in various products, such as dietary supplements, processed foods and drinks [26]. The efficacy and effectiveness studies carried out in animals and humans partially support the safety of LNT, however, only limited value was provided. The LD50 values of LNT were determined to be 250–500 mg/kg and >2500 mg/kg in mice (intravenously) and rats (intraperitoneally, subcutaneously and orally), respectively [27]. In a 6-month study, different doses of LNT (0.01, 0.1, 1, 10 mg/kg/day) were intravenously injected into the tail vein of rats to assess the chronic toxicity of LNT. All treated groups showed different degrees of arteritis, pulmonary hemorrhage and hypospermatogenesis, and the maximum no-effect level was lower than 0.01 mg/kg/day [28]. Based on this result and considering the direct cellular toxicity on cancer cells and the anti-bacterial effect [8,25,28], LNT may exert adverse effects on receivers. Thus, the health risk of LNT should be assessed adequately in different species and the underlying mechanism should be elucidated.
Before being administered to humans, the safety assessments of medicines, food ingredients or chemicals are essential. Currently, most of the toxicity assays are using animals, especially rodents. To improve animal welfare, test methods meeting the 3R principles (replacement, reduction, and refinement) are encouraged [29,30]. Zebrafish are becoming popular as an alternative animal model in safety assessment because of their particular advantages that partially obey the 3R principles. For instance, zebrafish are lower sentient vertebrates compared with rodents, which means the suffering during the experiments would be on a lower degree. The use of zebrafish for early screening could largely reduce the number of rodents in the subsequent research phase. Moreover, the fecundity of zebrafish is high and the embryos can be handled easily in vitro, which significantly increases the efficiency of experiments [31]. Zebrafish larva is a prominent model organism used in developmental biology due to its transparency, which allows easy staging under optical microscopy. A large number of studies have used zebrafish larvae to evaluate the developmental toxicity of chemicals [32,33,34]. More importantly, since the genome of zebrafish has been sequenced [35], the mechanism underlying the toxic phenotype is able to be elucidated at the molecular level.
In the present study, larval zebrafish were used to investigate the impacts of high concentrations of LNT on the developmental process in the early life stage. The hatching rate at 72 and 96 h post fertilization (hpf) and the median lethal concentration (LC50) at 96 hpf were determined. Furthermore, the morphological changes, heartbeat, blood flow velocity and locomotor activity of larval zebrafish that were continuously exposed to varying concentrations of LNT were studied at 120 hpf. In addition, the biochemical indexes reflecting energy metabolism were evaluated. To reveal the underlying mechanism, the genes involved in glucose and lipid metabolism were quantified at the transcriptional level. This study evaluated the potential adverse effects of LNT by using larval zebrafish for the first time. Moreover, these findings provided evidence that high concentrations of LNT would alter the energy metabolism process.
2. Materials and Methods
2.1. Research Compound
Lentinan (LNT, CAS NO. 37339-90-5, purity ≥98%) was obtained from YuanYe Bio-technology (Shanghai, China). The LNT was dissolved in E3 medium at 100 mg/mL and stored at −20 °C for further use.
2.2. Zebrafish Maintenance and Embryo Collection
Wild-type AB strain zebrafish were housed in a light- and temperature-controlled aquaculture facility with a light/dark photoperiod of 14:10 h. The fish were maintained in fish water at 28 °C (0.2% Instant Ocean Salt in deionized water, pH 6.9–7.2, conductivity 480–510 μS/cm and hardness 53.7–71.6 mg/L CaCO3) and were fed with live brine shrimp twice a day. Male and female zebrafish were set up for natural mating. All healthy embryos from different pairs of zebrafish were mixed and washed with E3 medium for further experiments. The experiment was performed following the ethical guideline of Experimental Animal Ethics Committee of Hunter Biotechnology, Inc. (AAALAC 001458).
2.3. Embryo Acute Toxicity Test
Fertilized eggs before 16 cell-stage were selected and randomly transferred into the six-well plates with 30 embryos per well, containing 3 mL of E3 medium. The embryos were exposed to series concentrations of LNT (600, 800, 1000, 1200, 1400, 1600, 1800, 2000, and 4000 μg/mL) continuously for 96 h. The media were refreshed every day. During the exposure period, the lethality was recorded according to the OECD guidelines [36], and the dead embryos were removed. The number of dead larvae at each concentration was plotted and a concentration–response curve was fitted. The LC50 value was calculated from independent experiments performed in triplicate. According to the 96 h LC50 value, four LNT concentrations (30, 100, 300, and 900 μg/mL) were set for further evaluation.
2.4. Pathological Alteration Analyses
The embryos at 6 hpf were exposed to series concentrations of LNT (30, 100, 300, and 900 μg/mL), and larval zebrafish at 120 hpf were collected. The numbers of hatched embryos at 72 and 96 hpf were recorded. The hatching rate was summarized from three independent assays. For morphological observation, the zebrafish larvae were anesthetized and images were captured using optical microscopy (SZX7, OLYMPUS, Japan). The body length, swim bladder size, eye area, and yolk sac area were analyzed using NIS-ElementsD 3.20 software (Nikon, Japan). The relative values were calculated by the following formula: relative values = individual values/mean value of control group. The zebrafish larvae were not anesthetized and the heartbeats in 20 s were recorded manually. The blood flow velocity was calculated using MicroZebraLab (Viewpoint, Lyon, France). For behavioral analyses, the zebrafish larvae were assigned individually into a 96-well microplate. After being acclimated for 5 min under the lighting conditions, the swimming ability in the light/dark cycle (5 min/5 min) was recorded for 25 min using Zebrabox (ViewPoint, Lyon, France).
2.5. Biochemical Indexes Determination
A total of 30 zebrafish larvae at 120 hpf were homogenized with 60 μL of ice-cold PBS using a homogenizer, centrifuged at 5000× g for 10 min at 4 °C, and the supernatant was collected for further analyses. The contents of Glucose (Glu), triglyceride (TG), and total cholesterol (T-CHO) were measured using the commercial kits obtained from Nanjing Jiancheng Institute of Biotechnology (Nanjing, China) according to the manufactures’ instructions. Briefly, 2.5 μL of supernatant from each sample was used to measure the glucose content by using the glucose oxidase method (GOD). Similarly, 2.5 μL of supernatant from each sample was used to measure the triglyceride content by using the GPO-PAP method and 2.5 μL of supernatant from each sample was used to measure the total cholesterol content by using the COD-PAP method.
2.6. Quantitative Real Time-Polymerase Chain Reaction (qRT-PCR) Analyses
Total RNA was isolated from 30 zebrafish larvae at 120 hpf using RNA-easy Isolation Reagent (Vazyme Biotech, Nanjing, China). Briefly, 30 larvae were homogenized with 300 μL of lysis buffer using a homogenizer, centrifuged at 12,000× g for 15 min, and the supernatant was transferred to a new tube. The same volume of isopropanol was added to precipitate the RNA. After being centrifuged at 12,000× g for 10 min, the pellet was washed with 75% ethanol. The RNA was dissolved with RNase-free ddH2O and the purity was checked using Nano Drop 2000 (Thermo Scientific, WALTHAM, MA, USA). A total of 2 μg RNA was reverse transcribed into cDNA by using FastKing RT kit (Tiangen Biotech, Beijing, China). Gene expressions were detected by qRT-PCR using iTaq Universal SYBR Green (Bio-Rad, Hercules, CA, USA) on the CFX Connect Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA). The primer sequences were referred to other studies and listed in Table S1 [37,38,39,40,41]. The relative expression levels of the genes were normalized to actb2, and calculated using the 2−ΔΔCT method.
2.7. Statistical Analysis
All data in this study were presented as mean ± standard error (SE), and analyzed by one-way analysis of variance (ANOVA) followed by the Dunnett’s multiple comparisons test using GraphPad Prism 7. Compared with the control group, p < 0.05 was considered significant.
3. Results
3.1. Lethal Effect and Developmental Toxicity of LNT in Larval Zebrafish
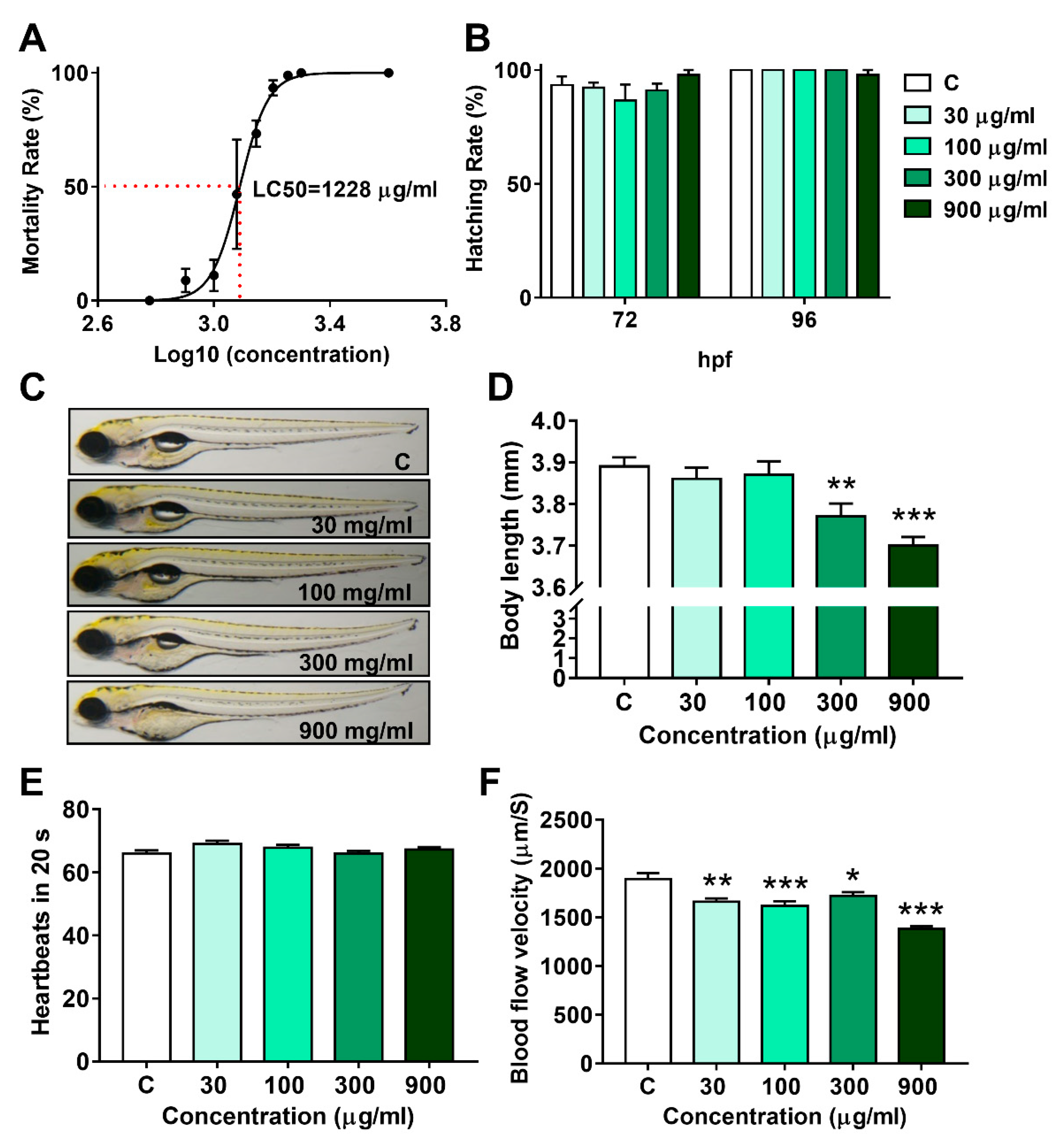
To investigate the lethal effect of LNT, fertilized eggs were exposed to series concentrations of LNT for 96 h. By analyzing the fitting curve shown in Figure 1A, the 96 h LC50 value of LNT was determined to be 1228 μg/mL.
Based on the LC50 value, LNT at concentrations of 30, 100, 300, and 900 μg/mL (1/36, 1/12, 1/4, and 3/4 LC50, respectively) were set for further evaluations. As shown in Figure 1B, the hatching rates at 72 h and 96 h were not affected by LNT treatment, even in the 900 μg/mL group. Except for yolk sac edema, no obvious deformity was observed in all LNT treated groups (Figure 1C). Nevertheless, the body length of zebrafish larvae decreased significantly in a dose-dependent manner (Figure 1D), suggesting that LNT exposure inhibited the growth of individuals. Interestingly, the blood flow velocity was remarkably reduced in all LNT treated groups, without any effect on the heartbeats in 20 s (Figure 1E,F).
3.2. LNT Induced Pathological Alterations in Larval Zebrafish
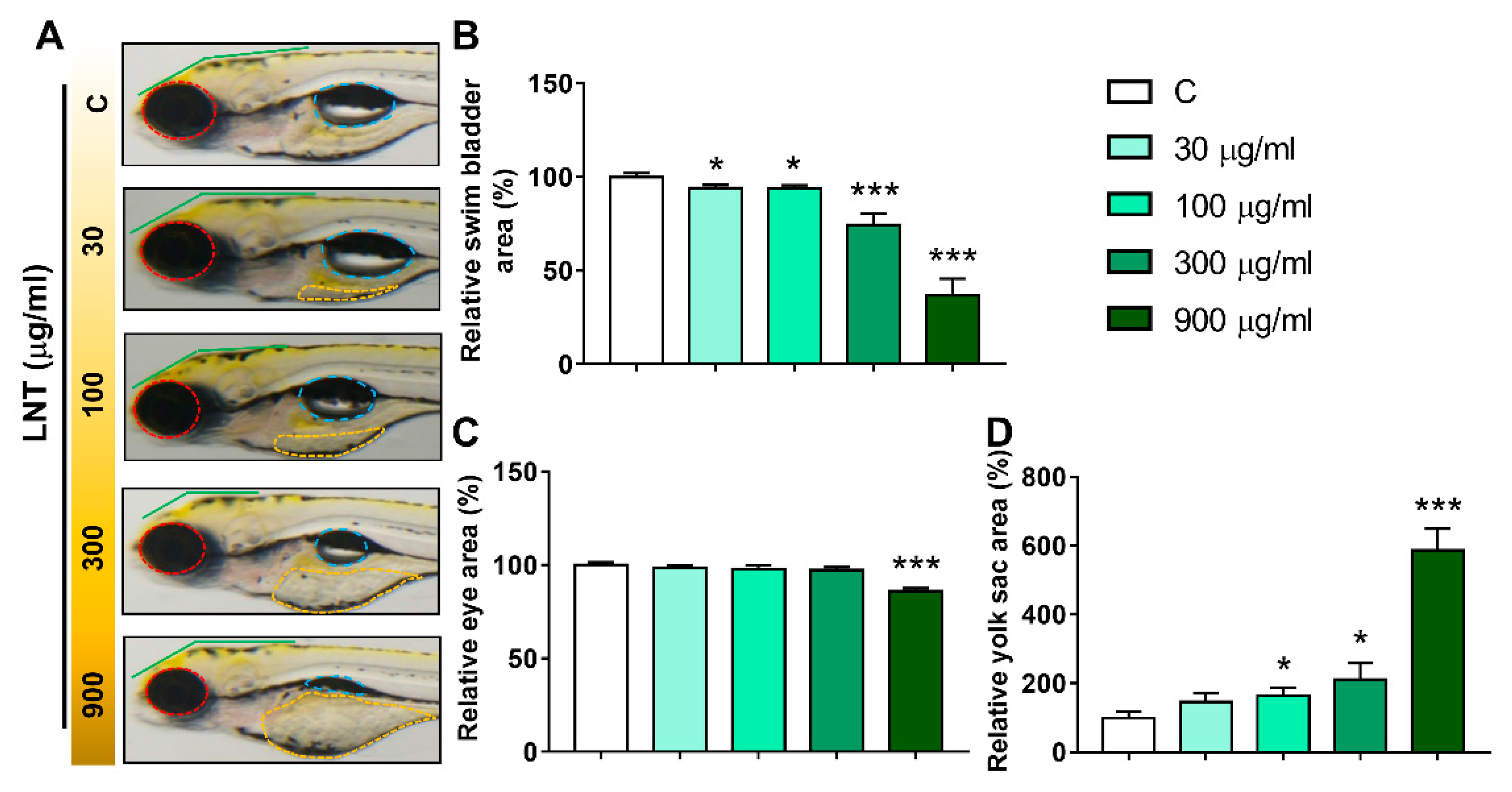
To further assess the adverse effects of LNT exposure on the development of larval zebrafish, more detailed characteristics, including eye and swim bladder sizes and yolk sac area, were quantified (Figure 2A). Inflated swim bladders (Figure 2A, blue dotted lines) were observed in the control group, while the sizes of the swim bladders were dose-dependently decreased in the LNT treated groups (Figure 2B). Notably, almost 50% of the zebrafish larvae completely lost the inflated swim bladders in the 900 μg/mL group. In addition, the eyes (Figure 2A, red dotted lines) in the 900 μg/mL group were significantly smaller than those in the control group (Figure 2C). Another obvious deformity observed was yolk sac edema. As shown in Figure 2D, the yolk sac area clearly increased along with the concentrations of LNT. All these morphological changes, including body length, eye and swim bladder sizes and yolk sac area, demonstrate that high concentrations of LNT inhibited the normal development of the larval zebrafish.
3.3. Effects of LNT Exposure on the Locomotor Activity in Larval Zebrafish
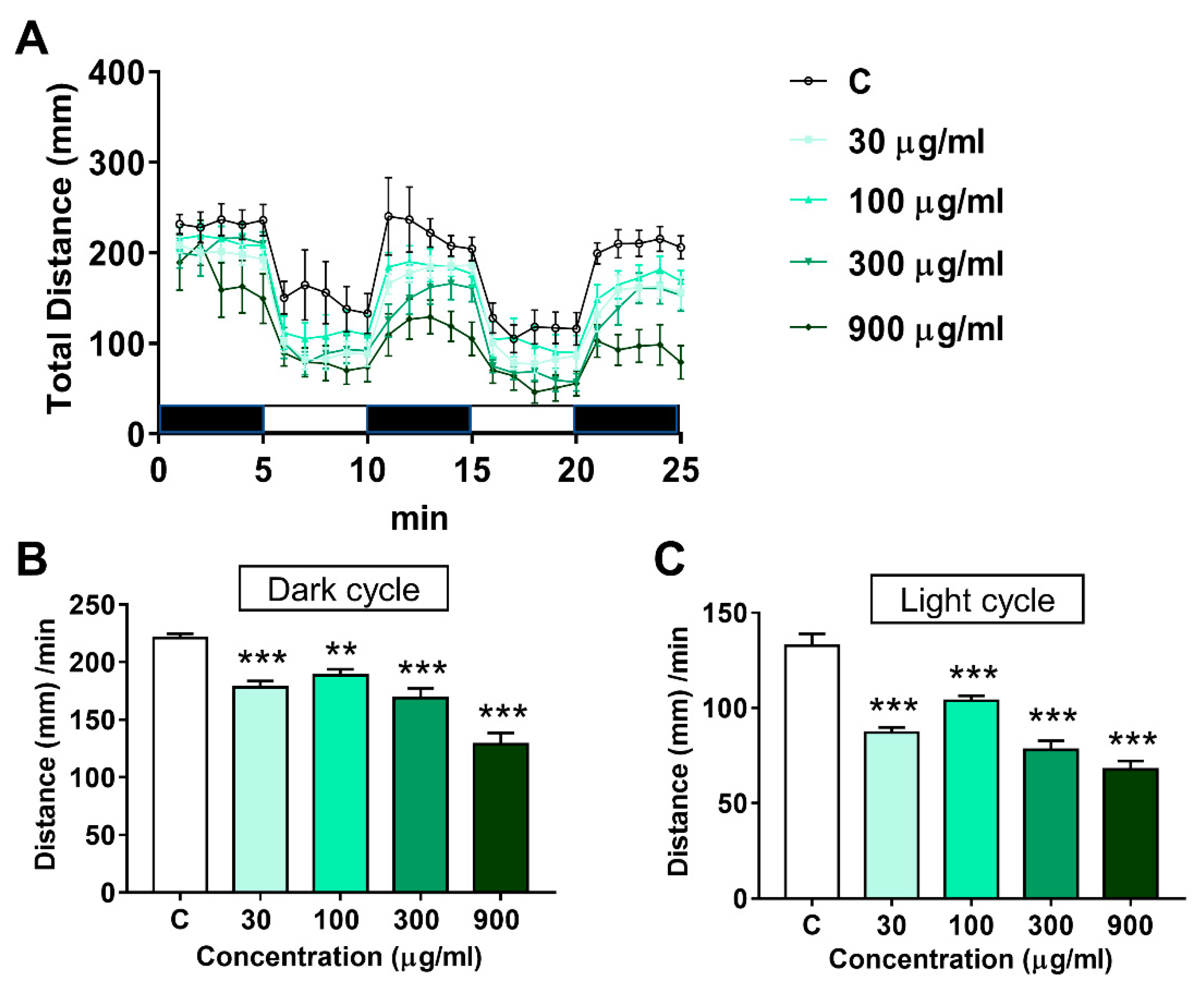
The impact of LNT on locomotor activity in larval zebrafish is shown in Figure 3. A light/dark rhythm was observed in the control group and all LNT treated groups (Figure 3A), suggesting that LNT exposure didn’t disrupt the response of zebrafish larvae to light and dark stimulation. Whereas, the total distance moved during the light and dark phases by the zebrafish larvae in the LNT treated groups decreased remarkably (Figure 3B,C).
3.4. Effects of LNT Exposure on the Glu, TG and T-CHO Levels in Larval Zebrafish
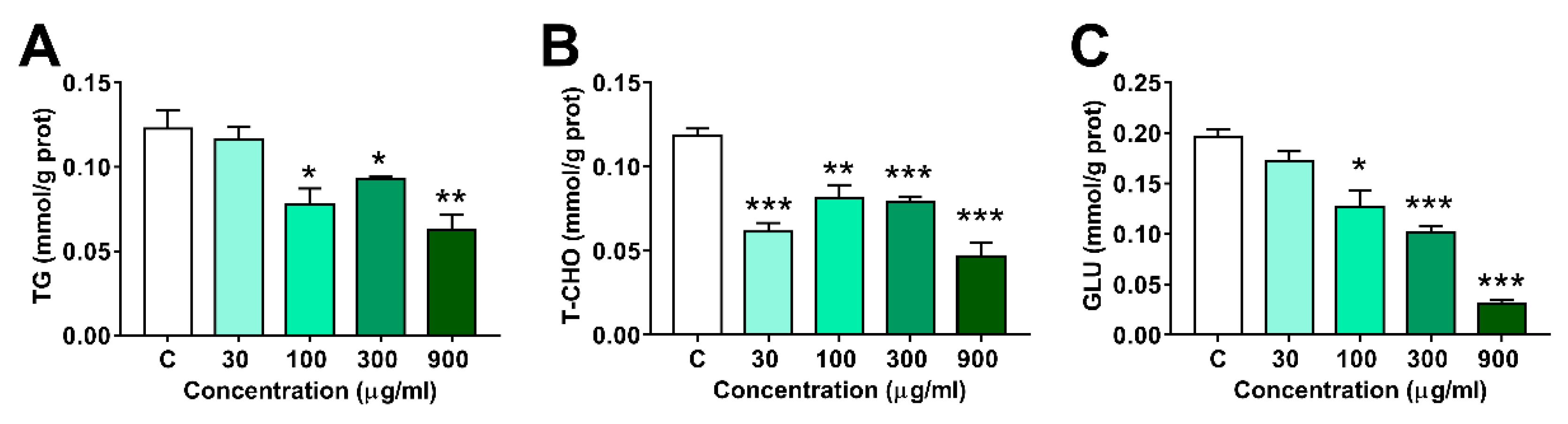
As shown in Figure 4, the contents of Glu and TG decreased dramatically in 100, 300 and 900 μg/mL groups, and T-CHO levels decreased in all LNT treated groups.
3.5. Effects of LNT Exposure on the Genes Involved in Glucose, Lipid and Cholesterol Metabolism in Larval Zebrafish
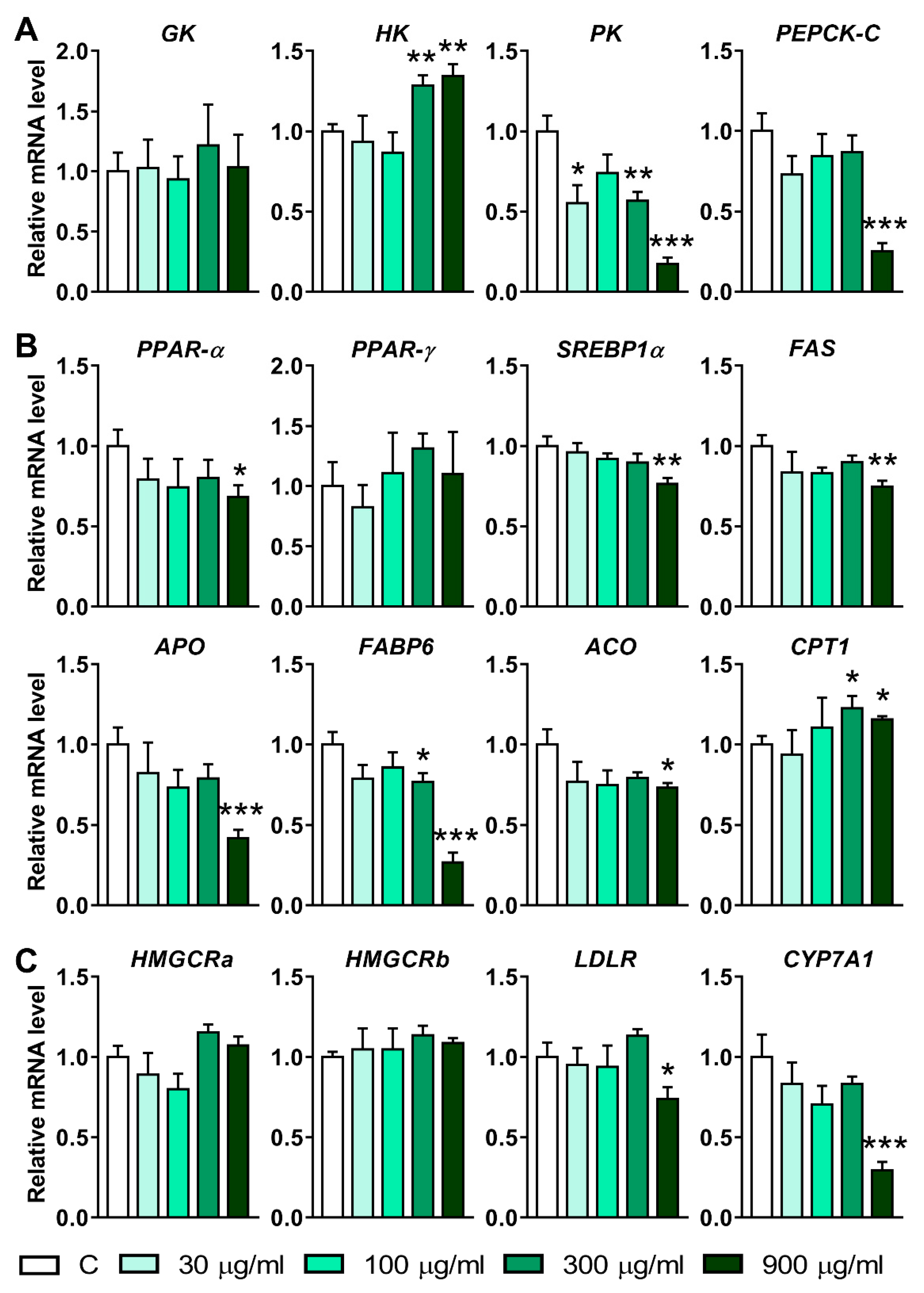
To elucidate the mechanism underlying the decrease in glucose, triglyceride, and cholesterol induced by LNT, the mRNA levels of the genes involved in the metabolism of these substances were quantified using qRT-PCR. The results from Figure 5A showed that the expression of Hexokinase (HK1) increased significantly after being treated with 300 and 900 μg/mL of LNT. On the contrary, the mRNA levels of pyruvate kinase (PK) in the 30, 300, and 900 μg/mL groups were lower than that in the control group. Additionally, the transcription level of cytosolic phosphoenolpyruvate carboxy kinase (PEPCK-C) decreased significantly in the 900 μg/mL group. These results suggest that LNT exposure affected the genes related to glucose metabolism.
In addition, the transcription levels of the genes involved in the lipid metabolism process were also disrupted by LNT exposition. The mRNA levels of peroxisome proliferator-activated receptor-α (PPAR-α), sterol regulatory element binding protein 1α (SREBP1α), fatty acid synthase (FAS), acyl-oxidase (ACO), and apolipoprotein A-IV (Apo) were downregulated after being treated with 900 μg/mL of LNT (Figure 5B). The mRNA levels of fatty acid binding protein 6 (FABP6) decreased in both the 300 and 900 μg/mL groups. Nevertheless, the transcription of carnitine palmitoyl transferase 1 (CPT1) was induced by 300 and 900 μg/mL of LNT (Figure 5B).
Finally, hydroxymethyl glutaryl coenzyme A reductase a (HMGCRa), HMGCRb, low-density lipoprotein receptor (LDLR) and cholesterol 7α-hydroxylase (CYP7A1), the genes related to the cholesterol metabolism, were quantified. The results showed that 900 μg/mL of LNT markedly inhibited the expressions of LDLR and CYP7A1 (Figure 5C). HMGCRa and HMGCRb were not affected by LNT.
4. Discussion
Natural medicines and functional foods derived from natural products, such as plants and fungi, are of huge value to humans. Compared to synthetic compounds, natural products are supposed to be lower toxic to the receivers [42]. In addition, one natural product contains diverse bioactive components, which may enhance the efficacy due to the synergistic effect [43]. In recent decades, comprehensive studies on natural products have been conducted in many countries, examining various aspects including preparation, bioavailability, and bioactivity. Glucans are products that are used as both medicines and functional foods. However, compared with the research evaluating their function and efficacy, adequate safety assessments are far from sufficient. Therefore, the present study was conducted to evaluate the toxicity and the underlying mechanism of high concentrations of LNT on larval zebrafish. Our results demonstrated that LNT exposure inhibited the development of larval zebrafish and caused morphological and locomotor defects. The glucose, triglyceride, and cholesterol levels were significantly decreased in the LNT treated groups. Further qRT-PCR assays revealed that transcriptions of the genes involved in glucose, lipid, and cholesterol metabolism were disrupted by LNT.
After being established as a model organism, zebrafish embryos have been widely used in scientific research, especially in toxicology [44]. The Organization for Economic Co-operation and Development (OECD) adopted the Fish Embryo Acute Toxicity (FET) Test as a method for the testing of chemicals in 2013 [36], which greatly promoted the use of zebrafish embryos for safety assessment. Recently, Wan-Mohtar et al. investigated the toxicity of the polysaccharides extracted from European Ganoderma applanatum mushrooms, and reported that the LC50 of exopolysaccharide (EPS) and endopolysaccharide (ENS) were 1410 and 870 μg/mL respectively [45]. Here, the half-lethal concentration of LNT at 96 h was demonstrated to be 1228 μg/mL (Figure 1A), suggesting that the toxicities of glucans differ by species and even by portions. From another view, these results also reinforced that the FET test is a reliable and sensitive method to assess the safety of natural product extracts.
The hatching rate was not significantly affected by LNT at concentrations of 30, 100, 300, and even 900 μg/mL. Interestingly, several other studies also suggested that the glucans appear to have little effect on the hatching rate of embryos at concentrations lower than 1000 μg/mL [45,46]. However, different degrees of morphological defects were observed in the larval zebrafish, manifesting as short body length, slow blood flow velocity, small eyes and swim bladder, and yolk sac edema, which could be summarized as developmental retardation [47]. It is well known that energy is essential for living organisms to carry out activities such as growth, development, movement, and reproduction [48]. We hypothesized that the energy balance might be disrupted by LNT in the zebrafish embryos. Indeed, the levels of glucose, triglyceride, and cholesterol were significantly lower in the LNT treated groups. The LNT induced growth retardation but did not affect the hatching rate, suggesting that the zebrafish embryos are more resistant to LNT than the larvae, which may attribute to the protective effect of the chorion [49].
Locomotor activity plays a crucial role in the survival of animals, because it has been closely related to migration and predator avoidance [50,51]. It is well known that locomotor activity is influenced by several factors, such as the nervous system, feeding, and growth [50]. Altered locomotor activity has been used as an indicator for investigating neurotoxicity, developmental toxicity and some diseases [52]. Here, we observed that LNT exposure remarkably inhibited the locomotor activity in larval zebrafish (Figure 3). Combined with the abnormal morphological changes, all these results suggest that LNT adversely affected the growth of larval zebrafish in the early stage. In addition, because movement is an energy cost behavior, the decrease in glucose, lipid, and cholesterol induced by LNT also could directly affect the locomotor activity of zebrafish larvae.
Without any doubt, the nutrients, such as glucose and lipids, are the foundation of development for all organisms [53]. Hence, the contents of these substances were measured in the larval zebrafish at 5 dpf. In accordance with our hypothesis, the glucose, triglyceride, and cholesterol levels were dramatically downregulated by LNT (Figure 4). Indeed, it was reported that β-glucans from different sources reduced the blood glucose level in both healthy and diabetic individuals [54,55,56]. In a mouse model, the β-glucan from barley was demonstrated to have the potential to reduce blood glucose and serum lipid [56]. Further investigation revealed that the genes involved in lipid metabolism were significantly altered after the β-glucan treatment [57]. Similarly, several key genes related to glucose and lipid metabolism were found to be regulated by LNT in the present study. HK, the initial enzyme in the glycolysis pathway [58], was induced by LNT. On the contrary, PEPCK-C, the key enzyme in glucose synthesis [59], was suppressed at the transcription level by LNT. These results indicated that the consumption of glucose was increased, while the production of glucose was decreased. As a consequence, the glucose content reduced significantly in the larval zebrafish. Interestingly, PK, the enzyme involved in the last step of glycolysis [60], was inhibited at the transcription level by LNT, which may lead to a deficiency in energy. In addition, LNT suppressed the expression of genes involved in lipid synthesis, such as SREBP1α and FAS, and enhanced the expression of genes related to catabolism of lipids, like CPT1. Similar results were observed in a previous study [57], showing that barley β-glucan induced the expression of CPT1 and reduced the expression of SREBP1α.
The cholesterol-lowering effect of β-glucan has been demonstrated in humans and animal models previously and different mechanisms were elucidated [61,62,63]. For instance, by-products of β-glucan digestion were reported to affect the synthesis of cholesterol [64]. Moreover, metabolism and circulation of bile acid were also influenced by β-glucan, which may be responsible for the lowering of cholesterol [63]. Our results showed that HMGCRa and HMGCRb, two key genes involved in cholesterol synthesis, were not altered by LNT. Whereas, the mRNA level of CYP7A1 was significantly inhibited by LNT, indicating that LNT regulated the cholesterol through the bile acid system, instead of direct regulation of cholesterol synthesis. However, the detailed mechanism remains unclear.
LNT has been tested in several clinical trials. For instance, LNT significantly prolonged the survival of gastric cancer patients treated with fluoropyrimidine-based chemotherapy. The ratio of granulocytes/lymphocytes was enhanced by LNT in the patients receiving LNT compared to the individuals with chemotherapy alone, supporting the immunomodulatory effect of LNT [65]. A meta-analysis also suggested that LNT prolonged the survival of advanced gastric cancer patients treated with standard chemotherapy [66]. The beneficial effects of β-glucans from mushrooms on the cardio-metabolic system have been reported in in vitro and animal experiments, with different mechanisms which were not completely understood [67,68]. However, the data from humans supporting the beneficial effect of LNT on lipid and cholesterol metabolism is scarce. A clinical trial showed that an eight-week intervention with β-glucans obtained from Shiitake mushrooms did not alter the lipid- or cholesterol-related parameters in hypercholesterolemic subjects receiving β-glucans or placebo [69]. Nevertheless, our results showed that LNT significantly affected the metabolism of glucose, triglyceride, and cholesterol at both biochemical and transcriptional levels. Although the exact reasons involved in the different results observed in humans and zebrafish are still to be elucidated, several potential factors may have contributed to the uncertain and inconsistent findings: (1) the type of subjects included in the studies (healthy, hypercholesterolemic, and diabetic subjects); (2) the different mushroom species used as the source of β-glucans; (3) different digestion processes in humans and zebrafish.
5. Conclusions
LNT exposure had toxic effects on the zebrafish embryos. The 96 h LC50 value was estimated to be 1228 μg/mL in the present study. LNT at the concentrations of 30, 100, 300, and 900 μg/mL did not alter the hatching rate of embryos at 72 and 96 hpf. However, the development of the embryos was impaired, which was evidenced by shortened body length, slowed blood flow velocity, small eyes and swim bladder, and yolk sac edema. In addition, the locomotor activity was also inhibited by LNT. Detection of the biochemical indexes and the gene transcriptions revealed that LNT significantly disrupted the glucose, lipid, and cholesterol metabolism, which might contribute to the LNT-induced energy deficiency. All these results suggested that high concentrations of LNT induced developmental toxicity in the zebrafish embryos, which can possibly be attributed to the disruption of energy metabolism induced by LNT.
Supplementary Materials
The following supporting information can be downloaded at: https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/pr10010120/s1, Table S1: The primer sequence pairs used in the qRT-PCR. Table S2: Effects of LNT on the GLU, TG and T-CHO in zebrafish larvae.
Author Contributions
Conceptualization, W.M. and C.L.; data curation, W.M.; funding acquisition, W.M.; investigation, W.M., L.H. and T.Z.; methodology, W.M., L.H. and C.L.; project administration, W.M.; supervision, C.L.; writing—original draft, W.M.; writing—review & editing, W.M. and C.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Scientific Research Fund of Zhejiang Provincial Education Department (Y201940954).
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Ethics Committee of Hunter Biotechnology, Inc. (AAALAC 001458; protocol code IACUC-2021-0012 and 26 November 2021).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available within the article and Supplementary Material.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Wang, Y.; Jin, H.; Yu, J.; Qu, C.; Wang, Q.; Yang, S.; Ma, S.; Ni, J. Quality control and immunological activity of lentinan samples produced in China. Int. J. Biol. Macromol. 2020, 159, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Zhang, G.; Kuai, J.; Fan, P.; Wang, X.; Zhou, P.; Yang, D.; Zheng, X.; Liu, X.; Wu, Q.; et al. Lentinan inhibits tumor angiogenesis via interferon γ and in a T cell independent manner. J. Exp. Clin. Cancer Res. 2018, 37, 260. [Google Scholar] [CrossRef] [Green Version]
- Maeda, Y.Y.; Hamuro, J.; Chihara, G. The mechanisms of action of anti-tumour polysaccharides. I. The effects of antilymphocyte serum on the anti-tumour activity of lentinan. Int. J. Cancer 1971, 8, 41–46. [Google Scholar] [CrossRef]
- Sasaki, T.; Takasuka, N. Further study of the structure of lentinan, an anti-tumor polysaccharide from Lentinus edodes. Carbohydr. Res. 1976, 47, 99–104. [Google Scholar] [CrossRef]
- Haba, S.; Hamaoka, T.; Takatsu, K.; Kitagawa, M. Selective suppression of t-cell activity in tumor-bearing mice and its improvement by lentinan, a potent anti-tumor polysaccharide. Int. J. Cancer 1976, 18, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Kozarski, M.; Klaus, A.; Nikšić, M.; Vrvić, M.M.; Todorović, N.; Jakovljević, D.; Van Griensven, L.J.L.D. Antioxidative activities and chemical characterization of polysaccharide extracts from the widely used mushrooms Ganoderma applanatum, Ganoderma lucidum, Lentinus edodes and Trametes versicolor. J. Food Compos. Anal. 2012, 26, 144–153. [Google Scholar] [CrossRef]
- Morales, D.; Rutckeviski, R.; Villalva, M.; Abreu, H.; Soler-Rivas, C.; Santoyo, S.; Iacomini, M.; Smiderle, F.R. Isolation and comparison of α- and β-Dglucans from shiitake mushrooms (Lentinula edodes) with different biological activities. Carbohydr. Polym. 2020, 229, 115521. [Google Scholar] [CrossRef] [PubMed]
- Hatvani, N. Antibacterial effect of the culture fluid of Lentinus edodes mycelium grown in submerged liquid culture. Int. J. Antimicrob. Agents 2001, 17, 71–74. [Google Scholar] [CrossRef]
- Zhao, Y.M.; Yang, J.M.; Liu, Y.H.; Zhao, M.; Wang, J. Ultrasound assisted extraction of polysaccharides from Lentinus edodes and its anti-hepatitis B activity in vitro. Int. J. Biol. Macromol. 2018, 107, 2217–2223. [Google Scholar] [CrossRef]
- Han, D.; Lee, H.T.; Lee, J.B.; Kim, Y.; Lee, S.J.; Yoon, J.W. A Bioprocessed Polysaccharide from Lentinus edodes Mycelia Cultures with Turmeric Protects Chicks from a Lethal Challenge of Salmonella gallinarum. J. Food Prot. 2017, 80, 245–250. [Google Scholar] [CrossRef]
- Song, X.; Ren, Z.; Wang, X.; Jia, L.; Zhang, C. Antioxidant, anti-inflammatory and renoprotective effects of acidic-hydrolytic polysaccharides by spent mushroom compost (Lentinula edodes) on LPS-induced kidney injury. Int. J. Biol. Macromol. 2020, 151, 1267–1276. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Li, J.; Kong, F.; Lin, J.; Gao, Y. Induction of immunomodulating cytokines by a new polysaccharide-peptide complex from culture mycelia of Lentinus edodes. Immunopharmacology 1998, 40, 187–198. [Google Scholar] [CrossRef]
- Jeff, I.B.; Fan, E.; Tian, M.; Song, C.; Yan, J.; Zhou, Y. In vivo anticancer and immunomodulating activities of mannogalactoglucan-type polysaccharides from Lentinus edodes (Berkeley) Singer. Cent. Eur. J. Immunol. 2016, 41, 47–53. [Google Scholar] [CrossRef]
- Wang, T.; He, H.; Liu, X.; Liu, C.; Liang, Y.; Mei, Y. Mycelial polysaccharides of Lentinus edodes (shiitake mushroom) in submerged culture exert immunoenhancing effect on macrophage cells via MAPK pathway. Int. J. Biol. Macromol. 2019, 130, 745–754. [Google Scholar] [CrossRef]
- Chen, S.; Liu, C.; Huang, X.; Hu, L.; Huang, Y.; Chen, H.; Fang, Q.; Dong, N.; Li, M.; Tang, W.; et al. Comparison of immunomodulatory effects of three polysaccharide fractions from Lentinula edodes water extracts. J. Funct. Foods 2020, 66, 103791. [Google Scholar] [CrossRef]
- Murphy, E.A.; Davis, J.M.; Carmichael, M.D. Immune modulating effects of beta-glucan. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 656–661. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Shao, Y.; Wang, Z.; Guo, Y. Effects of dietary yeast β-glucans supplementation on growth performance, gut morphology, intestinal Clostridium perfringens population and immune response of broiler chickens challenged with necrotic enteritis. Anim. Feed Sci. Technol. 2016, 215, 144–155. [Google Scholar] [CrossRef]
- Vetvicka, V.; Oliveira, C. beta(1-3)(1-6)-d-Glucans modulate immune status in pigs: Potential importance for efficiency of commercial farming. Ann. Transl. Med. 2014, 2, 16. [Google Scholar] [PubMed]
- Pilarski, F.; Ferreira de Oliveira, C.A.; Darpossolo de Souza, F.P.B.; Zanuzzo, F.S. Different beta-glucans improve the growth performance and bacterial resistance in Nile tilapia. Fish Shellfish Immunol. 2017, 70, 25–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.S.; Liau, S.Y.; Huang, C.T.; Nan, F.H. Beta 1,3/1,6-glucan and vitamin C immunostimulate the non-specific immune response of white shrimp (Litopenaeus vannamei). Fish Shellfish Immunol. 2016, 57, 269–277. [Google Scholar] [CrossRef]
- Kim, Y.S.; Ryu, J.H.; Han, S.J.; Choi, K.H.; Nam, K.B.; Jang, I.H.; Lemaitre, B.; Brey, P.T.; Lee, W.J. Gram-negative bacteria-binding protein, a pattern recognition receptor for lipopolysaccharide and beta-1,3-glucan that mediates the signaling for the induction of innate immune genes in Drosophila melanogaster cells. J. Biol. Chem. 2000, 275, 32721–32727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ina, K.; Kataoka, T.; Ando, T. The Use of Lentinan for Treating Gastric Cancer. Anticancer Agents Med. Chem. 2013, 13, 681–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, L.J.; Qi, M.; Li, N.; Lei, Y.H.; Zhang, D.M.; Chen, J.X. Natural products and their derivatives: Promising modulators of tumor immunotherapy. J. Leukoc. Biol. 2020, 108, 493–508. [Google Scholar] [CrossRef]
- Liu, Z.; He, J.; Zhu, T.; Hu, C.; Bo, R.; Wusiman, A.; Hu, Y.; Wang, D. Lentinan-Functionalized Graphene Oxide Is an Effective Antigen Delivery System That Modulates Innate Immunity and Improves Adaptive Immunity. ACS Appl. Mater. Interfaces 2020, 12, 39014–39023. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Y.; Zhou, Y.; Zheng, Z.; Tang, W.; Song, M.; Wang, J.; Wang, K. Lentinan inhibited colon cancer growth by inducing endoplasmic reticulum stress-mediated autophagic cell death and apoptosis. Carbohydr. Polym. 2021, 267, 118154. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on the safety of “Lentinus edodes extract” as a Novel Food ingredient. EFSA J. 2010, 8, 1685. [Google Scholar] [CrossRef]
- Moriyuki, H.; Ichimura, M. Acute toxicity of lentinan in mice and rats. J. Toxicol. Sci. 1980, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Shimazu, H.; Takeda, K.; Onodera, C.; Makita, I.; Hashi, T.; Yamazoe, T.; Kokuba, Y.; Tanigawa, H.; Ohkuma, S.; Shinpo, K.; et al. Intravenous chronic toxicity of lentinan in rats: 6-month treatment and 3-month recovery. J. Toxicol. Sci. 1980, 5, 33–57. [Google Scholar] [CrossRef]
- Russell, W.M.S.; Burch, R.L. The Principles of Humane Experimental Technique; Universities Federation for Animal Welfare: Wheathamsted, UK, 1959. [Google Scholar]
- Sewell, F.; Edwards, J.; Prior, H.; Robinson, S. Opportunities to Apply the 3Rs in Safety Assessment Programs. ILAR J. 2016, 57, 234–245. [Google Scholar] [CrossRef] [Green Version]
- Geisler, R.; Köhler, A.; Dickmeis, T.; Strähle, U. Archiving of zebrafish lines can reduce animal experiments in biomedical research. EMBO Rep. 2017, 18, 1–2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wei, L.; Wang, Y.; He, B.; Kong, B.; Zhu, J.; Jin, Y.; Fu, Z. Evaluation of development, locomotor behavior, oxidative stress, immune responses and apoptosis in developing zebrafish (Danio rerio) exposed to TBECH (tetrabromoethylcyclohexane). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2018, 217, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, R.; Jin, Y. Differential responses of larval zebrafish to the fungicide propamocarb: Endpoints at development, locomotor behavior and oxidative stress. Sci. Total Environ. 2020, 731, 139136. [Google Scholar] [CrossRef] [PubMed]
- Weng, Y.; Huang, Z.; Wu, A.; Yu, Q.; Lu, H.; Lou, Z.; Lu, L.; Bao, Z.; Jin, Y. Embryonic toxicity of epoxiconazole exposure to the early life stage of zebrafish. Sci. Total Environ. 2021, 778, 146407. [Google Scholar] [CrossRef]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OECD. OECD Guidelines for the Testing of Chemicals. Fish Embryo Acute Toxicity (FET) Test; OECD: Paris, France, 2013. [Google Scholar]
- Mu, X.; Wang, K.; Chai, T.; Zhu, L.; Yang, Y.; Zhang, J.; Pang, S.; Wang, C.; Li, X. Sex specific response in cholesterol level in zebrafish (Danio rerio) after long-term exposure of difenoconazole. Environ. Pollut. 2015, 197, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Qi, S.; Wang, D.; Wang, K.; Zhu, L.; Chai, T.; Wang, C. Toxic effects of thifluzamide on zebrafish (Danio rerio). J. Hazard. Mater. 2016, 307, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Cocco, A.; Ronnberg, A.M.; Jin, Z.; Andre, G.I.; Vossen, L.E.; Bhandage, A.K.; Thörnqvist, P.-E.; Birnir, B.; Winberg, S. Characterization of the gamma-aminobutyric acid signaling system in the zebrafish (Danio rerio Hamilton) central nervous system by reverse transcription-quantitative polymerase chain reaction. Neuroscience 2017, 343, 300–321. [Google Scholar] [CrossRef] [PubMed]
- Al-Habsi, A.A.; Massarsky, A.; Moon, T.W. Exposure to gemfibrozil and atorvastatin affects cholesterol metabolism and steroid production in zebrafish (Danio rerio). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2016, 199, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Bao, Z.; Wan, Z.; Fu, Z.; Jin, Y. Polystyrene microplastic exposure disturbs hepatic glycolipid metabolism at the physiological, biochemical, and transcriptomic levels in adult zebrafish. Sci. Total Environ. 2020, 710, 136279. [Google Scholar] [CrossRef] [PubMed]
- Markman, M. Safety issues in using complementary and alternative medicine. J. Clin. Oncol. 2002, 20, 39–41. [Google Scholar]
- Lahlou, M. Screening of natural products for drug discovery. Expert Opin. Drug Discov. 2007, 2, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Willett, C.; Fremgen, T. Zebrafish: An Animal Model for Toxicological Studies. Curr. Protoc. Toxicol. 2003, 17, 1.7.1–1.7.18. [Google Scholar] [CrossRef] [PubMed]
- Wan-Mohtar, W.A.A.Q.I.; Ilham, Z.; Jamaludin, A.A.; Rowan, N. Use of Zebrafish Embryo Assay to Evaluate Toxicity and Safety of Bioreactor-Grown Exopolysaccharides and Endopolysaccharides from European Ganoderma applanatum Mycelium for Future Aquaculture Applications. Int. J. Mol. Sci. 2021, 22, 1675. [Google Scholar] [CrossRef] [PubMed]
- Taufek, N.M.; Harith, H.H.; Abd Rahim, M.H.; Ilham, Z.; Rowan, N.; Wan, W.A.A.Q.I. Performance of mycelial biomass and exopolysaccharide from Malaysian Ganoderma lucidum for the fungivore red hybrid Tilapia (Oreochromis sp.) in Zebrafish embryo. Aquac. Rep. 2020, 17, 100322. [Google Scholar] [CrossRef]
- Park, H.; Lee, J.Y.; Park, S.; Song, G.; Lim, W. Developmental toxicity of fipronil in early development of zebrafish (Danio rerio) larvae: Disrupted vascular formation with angiogenic failure and inhibited neurogenesis. J. Hazard Mater. 2020, 385, 121531. [Google Scholar] [CrossRef]
- Zhu, J.J.; Thompson, C.B. Metabolic regulation of cell growth and proliferation. Nat. Rev. Mol. Cell Biol. 2019, 20, 436–450. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-Y.; Li, N.-J.; Cheng, F.-Y.; Hsueh, J.-F.; Huang, C.-C.; Lu, F.-I.; Fu, T.-F.; Yan, S.-J.; Lee, Y.-H.; Wang, Y.-J. The Effect of the Chorion on Size-Dependent Acute Toxicity and Underlying Mechanisms of Amine-Modified Silver Nanoparticles in Zebrafish Embryos. Int. J. Mol. Sci. 2020, 21, 2864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brodin, T.; Piovano, S.; Fick, J.; Klaminder, J.; Heynen, M.; Jonsson, M. Ecological effects of pharmaceuticals in aquatic systems—impacts through behavioural alterations. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130580. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Liang, X.; Zhao, Y.; Gu, X.; Mao, Z.; Zeng, Q.; Chen, H.; Martyniuk, C.J. Molecular and behavioral responses of zebrafish embryos/larvae after sertraline exposure. Ecotoxicol. Environ. Saf. 2021, 208, 111700. [Google Scholar] [CrossRef]
- Selderslaghs, I.W.; Hooyberghs, J.; De Coen, W.; Witters, H.E. Locomotor activity in zebrafish embryos: A new method to assess developmental neurotoxicity. Neurotoxicol. Teratol. 2010, 32, 460–471. [Google Scholar] [CrossRef]
- Nafikov, R.; Beitz, D.C. Carbohydrate and Lipid Metabolism in Farm Animals. J. Nutr. 2007, 137, 702–705. [Google Scholar] [CrossRef] [Green Version]
- Behall, K.M.; Scholfield, D.J.; Hallfrisch, J.G.; Liljeberg-Elmståhl, H.G. Consumption of Both Resistant Starch and -Glucan Improves Postprandial Plasma Glucose and Insulin in Women. Diabetes Care 2006, 29, 976–981. [Google Scholar] [CrossRef]
- Granfeldt, Y.; Nyberg, L.; Björck, I. Muesli with 4 g oat β-glucans lowers glucose and insulin responses after a bread meal in healthy subjects. Eur. J. Clin. Nutr. 2007, 62, 600–607. [Google Scholar] [CrossRef] [Green Version]
- El Khoury, D.; Cuda, C.; Luhovyy, B.L.; Anderson, G.H. Beta Glucan: Health Benefits in Obesity and Metabolic Syndrome. J. Nutr. Metab. 2012, 2012, 851362. [Google Scholar] [CrossRef] [Green Version]
- Aoe, S.; Mio, K.; Yamanaka, C.; Kuge, T. Low Molecular Weight Barley β-Glucan Affects Glucose and Lipid Metabolism by Prebiotic Effects. Nutrients 2021, 13, 130. [Google Scholar] [CrossRef]
- Tan, V.P.; Miyamoto, S. HK2/hexokinase-II integrates glycolysis and autophagy to confer cellular protection. Autophagy 2015, 11, 963–964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Méndez-Lucas, A.; Duarte, J.A.; Sunny, N.E.; Satapati, S.; He, T.; Fu, X.; Bermúdez, J.; Burgess, S.C.; Perales, J.C. Share PEPCK-M expression in mouse liver potentiates, not replaces, PEPCK-C mediated gluconeogenesis. J. Hepatol. 2013, 59, 105–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schormann, N.; Hayden, K.L.; Lee, P.; Banerjee, S.; Chattopadhyay, D. An overview of structure, function, and regulation of pyruvate kinases. Protein Sci. 2019, 28, 1771–1784. [Google Scholar] [CrossRef]
- Wolever, T.M.; Tosh, S.M.; Gibbs, A.L.; Brand-Miller, J.; Duncan, A.M.; Hart, V.; Lamarche, B.; Thomson, B.A.; Duss, R.; Wood, P.J. Physicochemical properties of oat beta-glucan influence its ability to reduce serum LDL cholesterol in humans: A randomized clinical trial. Am. J. Clin. Nutr. 2010, 92, 723–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Harding, S.V.; Eck, P.; Thandapilly, S.J.; Gamel, T.H.; Abdel-Aal, E.S.M.; Crow, G.H.; Tosh, S.M.; Jones, P.J.H.; Ames, N.P. High-molecular-weight beta-glucan decreases serum cholesterol differentially based on the CYP7A1 rs3808607 polymorphism in mildly hypercholesterolemic adults. J. Nutr. 2016, 146, 720–727. [Google Scholar] [CrossRef]
- Wang, Y.; Harding, S.V.; Thandapilly, S.J.; Tosh, S.M.; Jones, P.J.; Ames, N.P. Barley β-glucan reduces blood cholesterol levels via interrupting bile acid metabolism. Br. J. Nutr. 2017, 118, 822–829. [Google Scholar] [CrossRef] [Green Version]
- Gunness, P.; Gidley, M.J. Mechanisms underlying the cholesterol-lowering properties of soluble dietary fibre polysaccharides. Food Funct. 2010, 1, 149–155. [Google Scholar] [CrossRef]
- Ina, K.; Furuta, R.; Kataoka, T.; Kayukawa, S.; Yoshida, T.; Miwa, T.; Yamamura, Y.; Takeuchi, Y. Lentinan prolonged survival in patients with gastric cancer receiving S-1-based chemotherapy. World J. Clin. Oncol. 2011, 2, 339–343. [Google Scholar] [CrossRef] [PubMed]
- Oba, K.; Kobayashi, M.; Matsui, T.; Kodera, Y.; Sakamoto, J. Individual patient based meta-analysis of lentinan for unresectable/recurrent gastric cancer. Anticancer. Res. 2009, 29, 2739–2745. [Google Scholar]
- Sima, P.; Vannucci, L.; Vetvicka, V. β-glucans and cholesterol (Review). Int. J. Mol. Med. 2018, 41, 1799–1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wouk, J.; Dekker, R.F.H.; Queiroz, E.A.I.F.; Barbosa-Dekker, A.M. β-Glucans as a panacea for a healthy heart? Their roles in preventing and treating cardiovascular diseases. Int. J. Biol. Macromol. 2021, 177, 176–203. [Google Scholar] [CrossRef] [PubMed]
- Morales, D.; Shetty, S.A.; López-Plaza, B.; Gómez-Candela, C.; Smidt, H.; Marín, F.R.; Soler-Rivas, C. Modulation of human intestinal microbiota in a clinical trial by consumption of a β-d-glucan-enriched extract obtained from Lentinula edodes. Eur. J. Nutr. 2021, 60, 3249–3265. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Lethal effect and developmental toxicity of LNT in larval zebrafish. (A) Fitting curve of 96 h LC50 (each group n = 30, triplicate); (B) hatching rate at 72 and 96 hpf (each group n = 30, triplicate); (C) representative images of larvae at 120 hpf; (D) body length of larvae at 120 hpf (each group n = 20); (E) heartbeats of larvae at 120 hpf (each group n = 10); (F) blood flow velocity of larvae at 120 hpf (each group n = 10). All data were compared with the control group, *, p < 0.05; **, p < 0.01; ***, p < 0.001.
Figure 1.
Lethal effect and developmental toxicity of LNT in larval zebrafish. (A) Fitting curve of 96 h LC50 (each group n = 30, triplicate); (B) hatching rate at 72 and 96 hpf (each group n = 30, triplicate); (C) representative images of larvae at 120 hpf; (D) body length of larvae at 120 hpf (each group n = 20); (E) heartbeats of larvae at 120 hpf (each group n = 10); (F) blood flow velocity of larvae at 120 hpf (each group n = 10). All data were compared with the control group, *, p < 0.05; **, p < 0.01; ***, p < 0.001.

Figure 2.
LNT-induced pathological alterations in larval zebrafish. (A) Representative images of larvae at 120 hpf (red dotted line: eyes; blue dotted line: swim bladder; yellow dotted line: yolk sac); (B) relative swim bladder area; (C) relative eye area; (D) relative yolk sac area. Each group n = 20. All data were compared with the control group, *, p < 0.05; ***, p < 0.001.
Figure 2.
LNT-induced pathological alterations in larval zebrafish. (A) Representative images of larvae at 120 hpf (red dotted line: eyes; blue dotted line: swim bladder; yellow dotted line: yolk sac); (B) relative swim bladder area; (C) relative eye area; (D) relative yolk sac area. Each group n = 20. All data were compared with the control group, *, p < 0.05; ***, p < 0.001.

Figure 3.
Effects of LNT exposure on the locomotor activity in larval zebrafish. (A) Total distance moved by zebrafish larvae in 25 min; (B) distance moved by zebrafish larvae in one min in the dark cycle; (C) distance moved by zebrafish larvae in one min in the light cycle. Each group n = 12. All data were compared with the control group, **, p < 0.01; ***, p < 0.001.
Figure 3.
Effects of LNT exposure on the locomotor activity in larval zebrafish. (A) Total distance moved by zebrafish larvae in 25 min; (B) distance moved by zebrafish larvae in one min in the dark cycle; (C) distance moved by zebrafish larvae in one min in the light cycle. Each group n = 12. All data were compared with the control group, **, p < 0.01; ***, p < 0.001.

Figure 4.
Effects of LNT exposure on the Glu, TG and T-CHO levels in larval zebrafish. (A) TG content; (B) T-CHO content, (C) GLU content. Each group n = 4. All data were compared with the control group, *, p < 0.05; **, p < 0.01; ***, p < 0.001.
Figure 4.
Effects of LNT exposure on the Glu, TG and T-CHO levels in larval zebrafish. (A) TG content; (B) T-CHO content, (C) GLU content. Each group n = 4. All data were compared with the control group, *, p < 0.05; **, p < 0.01; ***, p < 0.001.

Figure 5.
Effects of LNT on the mRNA levels of metabolism-related genes in larval zebrafish. (A) Genes related to glucose metabolism; (B) genes related to lipid metabolism; (C) genes related to cholesterol metabolism. Each group n = 6. All data were compared with the control group, *, p < 0.05; **, p < 0.01; ***, p < 0.001.
Figure 5.
Effects of LNT on the mRNA levels of metabolism-related genes in larval zebrafish. (A) Genes related to glucose metabolism; (B) genes related to lipid metabolism; (C) genes related to cholesterol metabolism. Each group n = 6. All data were compared with the control group, *, p < 0.05; **, p < 0.01; ***, p < 0.001.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Miao, W.; He, L.; Zhang, T.; Li, C. Lentinan Impairs the Early Development of Zebrafish Embryos, Possibly by Disrupting Glucose and Lipid Metabolism. Processes 2022, 10, 120. https://0-doi-org.brum.beds.ac.uk/10.3390/pr10010120
AMA Style
Miao W, He L, Zhang T, Li C. Lentinan Impairs the Early Development of Zebrafish Embryos, Possibly by Disrupting Glucose and Lipid Metabolism. Processes. 2022; 10(1):120. https://0-doi-org.brum.beds.ac.uk/10.3390/pr10010120
Chicago/Turabian StyleMiao, Wenyu, Lingling He, Tao Zhang, and Chunqi Li. 2022. "Lentinan Impairs the Early Development of Zebrafish Embryos, Possibly by Disrupting Glucose and Lipid Metabolism" Processes 10, no. 1: 120. https://0-doi-org.brum.beds.ac.uk/10.3390/pr10010120
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.