
Effect of Bacillus Additives on Fermentation Quality and Bacterial Community during the Ensiling Process of Whole-Plant Corn Silage

Abstract
:1. Introduction
2. Materials and Methods
2.1. Inoculant Preparation
2.2. Silage Production
2.3. pH and Organic Acid Analyses
2.4. Nutrient Analyses
2.5. DON Analyses
2.6. Bacteria Community Analyses
2.7. Statistical Analyses
3. Results
3.1. Dynamic Changes in pH and Organic Acids during the Ensiling Process
3.2. Dynamic Changes in Chemical Compositions during the Ensiling Process
3.3. Dynamic Changes in DON Content during the Ensiling Process
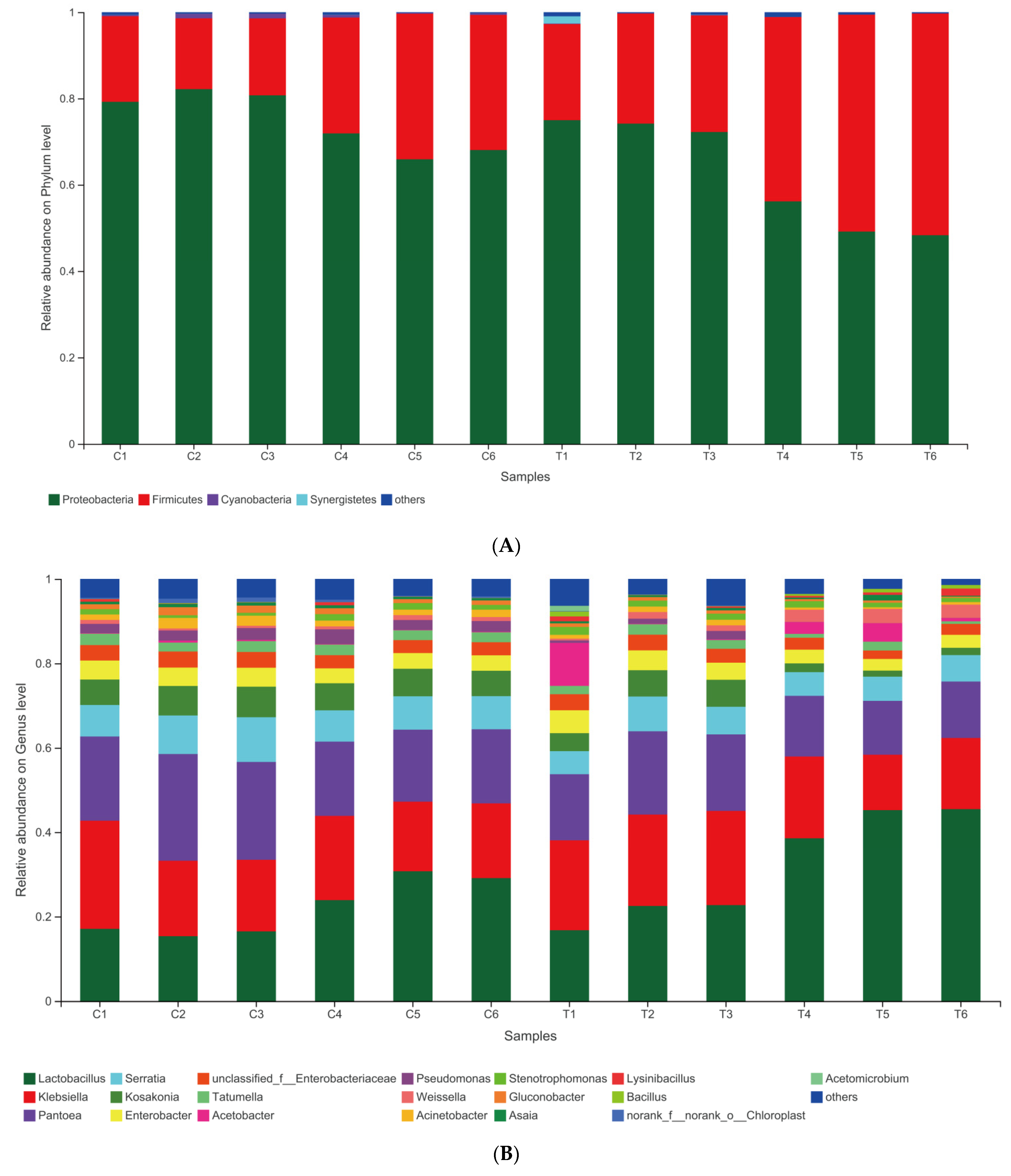
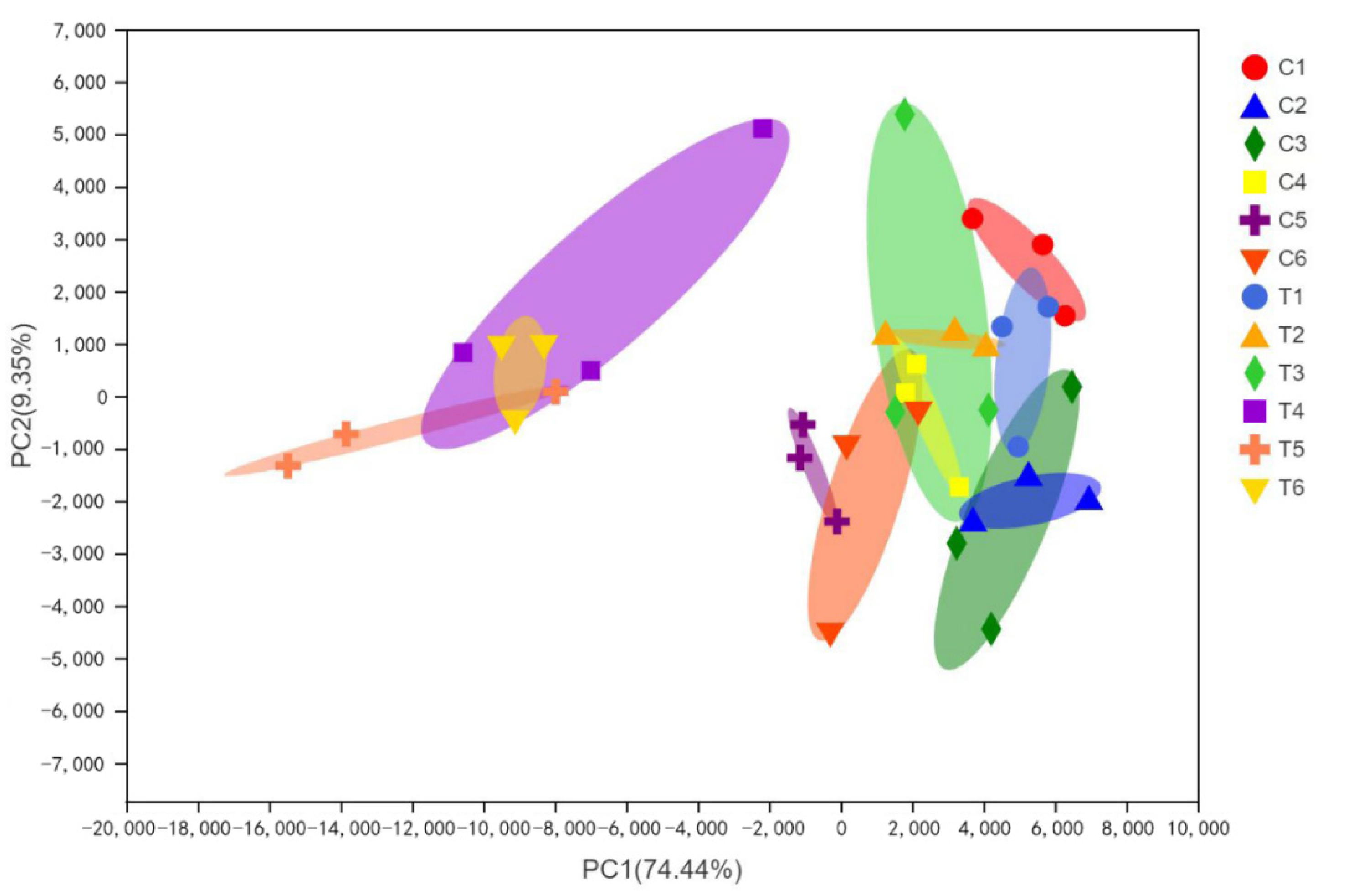
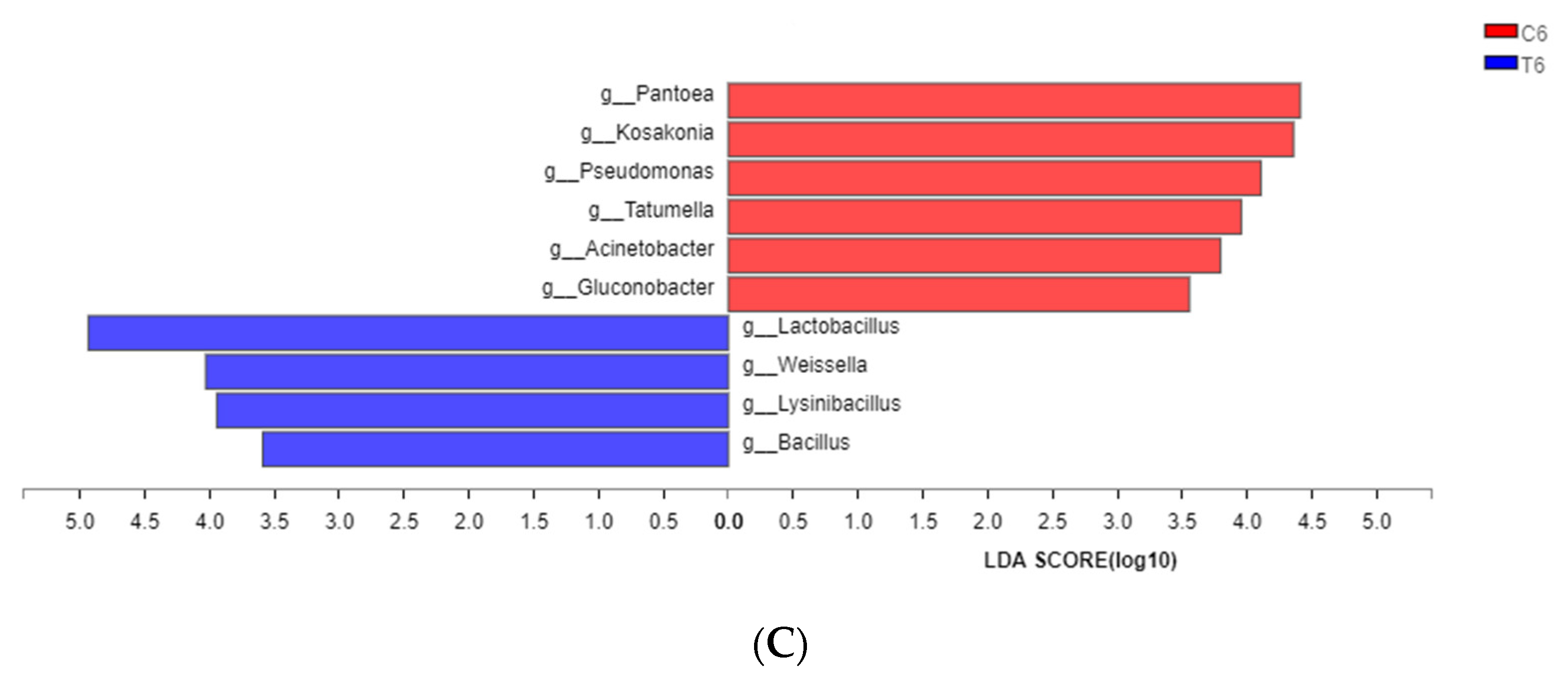
3.4. Dynamic Changes in Bacterial Communities during the Ensiling Process
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Z.; Xu, H.; Sun, P.; Zhao, B.; Dong, S. Research Progress on Harmless Treatment of Dead Livestock and Poultry with Alkaline Hydrolysis in China. Anim. Husb. Feed. Sci. 2017, 6, 10–12. [Google Scholar]
- Ni, K.; Wang, F.; Zhu, B.; Yang, J.; Zhou, G.; Pan, Y.; Tao, Y.; Zhong, J. Effects of lactic acid bacteria and molasses additives on the microbial community and fermentation quality of soybean silage. Bioresour. Technol. 2017, 238, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Sun, L.; Xu, H.; Na, N.; Yin, G.; Liu, S.; Jiang, Y.; Xue, Y. Microbial Communities, Metabolites, Fermentation Quality and Aerobic Stability of Whole-Plant Corn Silage Collected from Family Farms in Desert Steppe of North China. Processes 2021, 9, 784. [Google Scholar] [CrossRef]
- Ferraretto, L.F.; Shaver, R.D. Effects of whole-plant corn silage hybrid type on intake, digestion, ruminal fermentation, and lactation performance by dairy cows through a meta-analysis. J. Dairy Sci. 2015, 98, 2662–2675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nkosi, B.D.; Meeske, R.; Palic, D.; Langa, T.; Leeuw, K.J.; Groenewald, I.B. Effects of ensiling whole crop maize with bacterial inoculants on the fermentation, aerobic stability, and growth performance of lambs. Anim. Feed Sci. Technol. 2009, 154, 193–203. [Google Scholar] [CrossRef]
- Raffrenato, E.; Fievisohn, R.; Cotanch, K.W.; Grant, R.J.; Chase, L.E.; Van Amburgh, M.E. Effect of lignin linkages with other plant cell wall components on in vitro and in vivo neutral detergent fiber digestibility and rate of digestion of grass forages. J. Dairy Sci. 2017, 100, 8119–8131. [Google Scholar] [CrossRef] [Green Version]
- Wambacq, E.; Vanhoutte, I.; Audenaert, K.; Gelder, L.D.; Haesaert, G. Occurrence, prevention and remediation of toxigenic fungi and mycotoxins in silage: A review. J. Sci. Food Agric. 2016, 96, 2284–2302. [Google Scholar] [CrossRef]
- Dell’Orto, V.; Baldi, G.; Cheli, F. Mycotoxins in silage: Checkpoints for effective management and control. World Mycotoxin J. 2015, 8, 603–617. [Google Scholar] [CrossRef]
- Zain, M.E. Impact of mycotoxins on humans and animals. J. Saudi Chem. Soc. 2011, 15, 129–144. [Google Scholar] [CrossRef] [Green Version]
- Jensen, T.; Boevre, M.D.; Saeger, S.D.; Preuke, N.; Frank, D.S.; Kramer, E.; Klink, H.; Verreet, J.A.; Birr, T. Effect of ensiling duration on the fate of deoxynivalenol, zearalenone and their derivatives in maize silage. Mycotoxin Res. 2020, 36, 127–136. [Google Scholar] [CrossRef]
- Marta, L.; Andreia, F.; Ana Sanches, S.; Jorge, B.; Fernando, R. Maize food chain and mycotoxins: A review on occurrence studies. Trends Food Sci. Technol. 2021, 115, 307–331. [Google Scholar]
- Muck, R.E.; Nadeau, E.; Mcallister, T.A.; Contreras-Govea, F.E.; Kung, L.J. Silage review: Recent advances and future uses of silage additives. J. Dairy Sci. 2018, 101, 3980–4000. [Google Scholar] [CrossRef] [PubMed]
- Ferraretto, L.F.; Shaver, R.D.; Luck, B.D. Silage review: Recent advances and future technologies for whole-plant and fractionated corn silage harvesting. J. Dairy Sci. 2018, 101, 3937–3951. [Google Scholar] [CrossRef] [PubMed]
- Hassan, Z.U.; Thani, R.A.; Alsafran, M.; Migheli, Q.; Jaoua, S. Selection of Bacillus spp. with decontamination potential on multiple Fusarium mycotoxins. Food Control 2021, 127, 108119. [Google Scholar] [CrossRef]
- Guo, W.; Guo, X.J.; Zhu, B.C.; Guo, Y.Y.; Zhou, X. In situ degradation, ruminal fermentation, and the rumen bacterial community of cattles fed corn stover fermented by lignocellulolytic microorganisms. Anim. Feed Sci. Technol. 2019, 248, 10–19. [Google Scholar] [CrossRef]
- Xiao, J.W.; Guo, X.J.; Qiao, X.L.; Zhang, X.C.; Chen, X.M.; Zhang, D.D. Activity of fengycin and Iturin A isolated from Bacillus subtilis Z-14 on Gaeumannomyces graminis var. tritici and soil microbial diversity. Front. Microbiol. 2021, 12, 682437. [Google Scholar] [CrossRef]
- Ren, F.Y.; He, R.C.; Zhou, X.K.; Gu, Q.C.; Xia, Z.S.; Liang, M.Z.; Zhou, J.H.; Lin, B.; Zou, C.X. Dynamic changes in fermentation profiles and bacterial community composition during sugarcane top silage fermentation: A preliminary study. Bioresour. Technol. 2019, 285, 121315. [Google Scholar] [CrossRef]
- Wang, S.R.; Zhao, J.; Dong, Z.H.; Li, J.F.; Kaka, N.A.; Shao, T. Sequencing and microbiota transplantation to determine the role of microbiota on the fermentation type of oat silage. Bioresour. Technol. 2020, 309, 123371. [Google Scholar] [CrossRef]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Bai, J.; Xu, D.; Xie, D.; Wang, M.; Li, Z.; Guo, X. Effects of antibacterial peptide-producing Bacillus subtilis and Lactobacillus buchneri on fermentation, aerobic stability, and microbial community of alfalfa silage. Bioresour. Technol. 2020, 315, 123881. [Google Scholar] [CrossRef]
- Li, J.; Yuan, X.; Dong, Z.; Mugabe, W.; Shao, T. The effects of fibrolytic enzymes, cellulolytic fungi and bacteria on the fermentation characteristics, structural carbohydrates degradation, and enzymatic conversion yields of Pennisetum sinese silage. Bioresour. Technol. 2018, 264, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.S.; Weinberg, Z.G.; Ogunade, I.M.; Cervantes, A.A.P.; Arriola, K.G.; Yun, J.; Kim, D.; Li, X.; Goncalves, M.C.M.; Vyas, D. Meta-analysis of effects of inoculation with homofermentative and facultative heterofermentative lactic acid bacteria on silage fermentation, aerobic stability, and the performance of dairy cows. J. Dairy Sci. 2017, 100, 4587–4603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kung, L.M.; Shaver, R.D.; Grant, R.J.; Schmidt, R.J. Silage review: Interpretation of chemical, microbial, and organoleptic components of silages. J. Dairy Sci. 2018, 101, 4020–4033. [Google Scholar] [CrossRef] [PubMed]
- Lara, E.C.; Basso, F.C.; De Assis, F.B.; Souza, F.A.; Berchielli, T.T.; Reis, R.A. Changes in the nutritive value and aerobic stability of corn silages inoculated with Bacillus subtilis alone or combined with Lactobacillus plantarum. Anim. Prod. Sci. 2016, 56, 1867–1874. [Google Scholar] [CrossRef] [Green Version]
- Adesogan, A.T.; Krueger, N.; Salawu, M.B.; Dean, D.B.; Staples, C.R. The influence of treatment with dual purpose bacterial inoculants or soluble carbohydrates on the fermentation and aerobic stability of bermudagrass. J. Dairy Sci. 2004, 87, 3407–3416. [Google Scholar] [CrossRef] [Green Version]
- Ellis, J.L.; Hindrichsen, I.K.; Klop, G.; Kinley, R.D.; Milora, N.; Bannink, A.; Dijkstra, J. Effects of lactic acid bacteria silage inoculation on methane emission and productivity of Holstein Friesian dairy cattle. J. Dairy Sci. 2016, 99, 7159–7174. [Google Scholar] [CrossRef]
- Larsen, S.U.; Hjort-Gregersen, K.; Vazifehkhoran, A.H.; Triolo, J.M. Co-ensiling of straw with sugar beet leaves increases the methane yield from straw. Bioresour. Technol. 2017, 245, 106–115. [Google Scholar] [CrossRef]
- Ilavenil, S.; Soo, P.H.; Sathya, R.; Ravikumar, S.; Choon, C.K. Application and Future Prospective of Lactic Acid Bacteria as Natural Additives for Silage Production—A Review. Appl. Sci. 2021, 11, 8127. [Google Scholar]
- Bai, J.; Franco, M.; Ding, Z.T.; Hao, L.; Ke, W.C.; Wang, M.S.; Xie, D.M.; Li, Z.Q.; Ai, L.; Guo, X.S. Effect of Bacillus amyloliquefaciens and Bacillus subtilis on fermentation, dynamics of bacterial community and their functional shifts of whole-plant corn silage. J. Anim. Sci. Biotechnol. 2022, 13, 7. [Google Scholar] [CrossRef]
- Ning, T.; Wang, H.; Zheng, M.; Niu, D.; Zuo, S.; Xu, C. Effects of microbial enzymes on starch and hemicellulose degradation in total mixed ration silages. Asian Austral. J. Anim. Sci. 2017, 30, 171–180. [Google Scholar] [CrossRef]
- Tian, X.Y.; Song, F.P.; Zhang, J.; Liu, R.M.; Zhang, X.P.; Duan, J.Y.; Shu, C.L. Diversity of gut bacteria in larval Protaetia brevitarsis (Coleoptera:Scarabaedia) fed on corn stalk. Acta Entomol. Sin. 2017, 60, 632–641. [Google Scholar]
- Dogi, C.A.; Fochesato, A.; Armando, R.; Pribull, B.; de Souza, M.M.S.; da Silva Coelho, I.; Araújo de Melo, D.; Dalcero, A.; Cavaglieri, L. Selection of lactic acid bacteria to promote an efficient silage fermentation capable of inhibiting the activity of Aspergillus parasiticus and Fusarium gramineraum and mycotoxin production. J. Appl. Microbiol. 2013, 114, 1650–1660. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Fancello, F.; Ghilardelli, F.; Zara, S.; Spanghero, M. Effects of several commercial or pure lactic acid bacteria inoculants on fermentation and mycotoxin levels in high-moisture corn silage. Anim. Feed Sci. Technol. 2022, 286, 115256. [Google Scholar] [CrossRef]
- Gao, X.; Ma, Q.; Zhao, L.; Lei, Y.; Shan, Y.; Cheng, J. Isolation of Bacillus subtilis: Screening for aflatoxins B1, M1, and G1 detoxification. Eur. Food Restechnol. 2011, 232, 957–962. [Google Scholar] [CrossRef]
- Yi, P.J.; Pai, C.K.; Liu, J.R. Isolation and characterization of a Bacillus licheniformis strain capable of degrading zearalenone. World J. Microbol. Biotechnol. 2011, 27, 1035–1043. [Google Scholar] [CrossRef]
- Dunière, L.; Sindou, J.; Chaucheyras-Durand, F.; Chevallier, I.; Thévenot- Sergentet, D. Silage processing and strategies to prevent persistence of undesirable microorganisms. Anim. Feed Sci. Technol. 2013, 182, 1–15. [Google Scholar] [CrossRef]
- Muck, R.E. Recent advances in silage microbiology. Agric. Food Sci. 2013, 22, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Eikmeyer, F.G.; Köfinger, P.; Poschenel, A.; Jünemann, S.; Zakrzewski, M.; Heinl, S.; Mayrhuber, E.; Grabherr, R.; Pühler, A.; Schwab, H.; et al. Metagenome analyses reveal the influence of the inoculant Lactobacillus buchneri CD034 on the microbial community involved in grass ensiling. J. Biotechnol. 2013, 167, 334–343. [Google Scholar] [CrossRef]
- Yang, L.L.; Yuan, X.J.; Li, J.F.; Dong, Z.H.; Shao, T. Dynamics of microbial community and fermentation quality during ensiling of sterile and nonsterile alfalfa with or without Lactobacillus plantarum inoculant. Bioresour. Technol. 2019, 275, 280–287. [Google Scholar] [CrossRef]
- Keshri, J.; Chen, Y.; Pinto, R.; Kroupitski, Y.; Weinberg, Z.G.; Sela Saldinger, S. Microbiome dynamics during ensiling of corn with and without Lactobacillus plantarum inoculant. Appl. Microbiol. Biot. 2018, 102, 4025–4037. [Google Scholar] [CrossRef]
- Ni, K.; Zhao, J.; Zhu, B.; Su, R.; Pan, Y.; Ma, J.; Zhou, G.; Tao, Y.; Liu, X.; Zhong, J. Assessing the fermentation quality and microbial community of the mixed silage of forage soybean with crop corn or sorghum. Bioresour. Technol. 2018, 265, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.H.; Li, X.M.; Guan, H.; Huang, L.K.; Ma, X.; Peng, Y.; Li, Z.; Nie, G.; Zhou, J.Q.; Yang, W.Y.; et al. Microbial community and fermentation characteristic of Italian ryegrass silage prepared with corn stover and lactic acid bacteria. Bioresour. Technol. 2019, 279, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Guan, H.; Yan, Y.H.; Li, X.L.; Li, X.M.; Shuai, Y.; Feng, G.Y.; Ran, Q.F.; Cai, Y.M.; Li, Y.; Zhang, X.Q. Microbial communities and natural fermentation of corn silages prepared with farm bunker-silo in Southwest China. Bioresour. Technol. 2018, 265, 282–290. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Days | pH Values | LA (%) | AA (%) | BA (%) |
|---|---|---|---|---|---|
| C | 1 | 5.64 a | 1.95 a | 0.32 a | - |
| 3 | 4.25 c | 2.62 b | 0.35 a | - | |
| 5 | 3.95 de | 2.79 b | 0.32 a | - | |
| 10 | 3.83 fg | 3.11 c | 0.38 a | - | |
| 30 | 3.88 def | 3.89 d | 0.43 a | - | |
| 45 | 3.85 ef | 3.82 d | 0.44 a | - | |
| T | 1 | 4.55 b | 2.66 b | 0.75 b | - |
| 3 | 3.97 d | 5.02 e | 1.21 c | - | |
| 5 | 3.74 g | 5.78 f | 1.40 d | - | |
| 10 | 3.79 fg | 6.32 g | 1.45 d | - | |
| 30 | 3.81 fg | 7.88 h | 1.92 e | - | |
| 45 | 3.80 fg | 7.95 h | 1.98 e | - | |
| SEM | 0.072 | 0.268 | 0.078 | - | |
| p-Value | |||||
| G | <0.01 | <0.01 | <0.01 | ||
| D | <0.01 | <0.01 | <0.01 | ||
| G × D | <0.01 | <0.01 | <0.01 |
| Groups | Days | NDF (%) | ADF (%) | CP (%) | Starch (%) |
|---|---|---|---|---|---|
| C | 1 | 40.51 a | 29.21 a | 8.88 | 35.62 |
| 3 | 38.83 b | 28.53 ab | 8.72 | 34.49 | |
| 5 | 37.23 cd | 28.44 b | 8.57 | 35.12 | |
| 10 | 36.28 de | 27.92 b | 8.35 | 35.54 | |
| 30 | 35.21 ef | 27.22 c | 8.35 | 35.26 | |
| 45 | 34.92 f | 26.88 c | 8.05 | 35.89 | |
| T | 1 | 39.52 ab | 28.24 b | 8.88 | 35.15 |
| 3 | 37.55 c | 26.52 c | 8.95 | 36.25 | |
| 5 | 35.62 ef | 24.98 d | 8.91 | 35.25 | |
| 10 | 32.54 g | 22.24 e | 8.98 | 35.99 | |
| 30 | 31.56 h | 21.93 f | 9.01 | 35.62 | |
| 45 | 30.25 i | 21.55 f | 9.00 | 36.20 | |
| SEM | 0.403 | 0.330 | 0.115 | 0.518 | |
| p-Value | |||||
| G | <0.01 | <0.01 | 0.204 | 0.208 | |
| D | <0.01 | <0.01 | 0.085 | 0.105 | |
| G × D | <0.01 | <0.01 | 0.125 | 0.226 |
| Groups | Days | DON (μg/kg) |
|---|---|---|
| C | 1 | 32.67 a |
| 3 | 150.11 c | |
| 5 | 401.32 fg | |
| 10 | 499.42 h | |
| 30 | 415.32 g | |
| 45 | 382.51 f | |
| T | 1 | 42.52 a |
| 3 | 102.42 b | |
| 5 | 289.56 e | |
| 10 | 298.93 e | |
| 30 | 191.31 d | |
| 45 | 205.67 d | |
| SEM | 19.94 | |
| p-Value | ||
| G | <0.001 | |
| D | <0.001 | |
| G × D | <0.001 |
| Groups | Days | Reads | OTU | Shannon | Chao1 | Coverage (%) |
|---|---|---|---|---|---|---|
| C | 1 | 62,067.33 | 898 | 3.04 | 758.67 | 99.65 |
| 3 | 61,457.67 | 705 | 3.09 | 632.37 | 99.72 | |
| 5 | 63,284.33 | 678 | 3.08 | 613.16 | 99.73 | |
| 10 | 61,296.67 | 903 | 3.11 | 707.33 | 99.68 | |
| 30 | 64,128.33 | 526 | 2.93 | 467.68 | 99.82 | |
| 45 | 56,734.67 | 702 | 2.99 | 570.96 | 99.74 | |
| T | 1 | 59,743.00 | 848 | 3.25 | 640.05 | 99.71 |
| 3 | 63,531.67 | 624 | 2.94 | 518.76 | 99.79 | |
| 5 | 63,846.33 | 834 | 3.08 | 617.08 | 99.77 | |
| 10 | 69,875.00 | 648 | 2.73 | 533.82 | 99.82 | |
| 30 | 72,961.67 | 559 | 2.63 | 459.89 | 99.86 | |
| 45 | 68,830.33 | 479 | 2.60 | 453.64 | 99.84 |
| Days | Relative Abundance (%) | SEM | p-Value | |
|---|---|---|---|---|
| C | T | |||
| 1 | 0.049 | 1.088 | 0.254 | <0.001 |
| 3 | 0.052 | 1.111 | 0.202 | <0.001 |
| 5 | 0.043 | 1.003 | 0.215 | <0.001 |
| 10 | 0.052 | 1.251 | 0.203 | <0.001 |
| 30 | 0.053 | 1.439 | 0.185 | <0.001 |
| 45 | 0.054 | 1.433 | 0.198 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, X.; Guo, W.; Yang, M.; Sun, Y.; Wang, Y.; Yan, Y.; Zhu, B. Effect of Bacillus Additives on Fermentation Quality and Bacterial Community during the Ensiling Process of Whole-Plant Corn Silage. Processes 2022, 10, 978. https://0-doi-org.brum.beds.ac.uk/10.3390/pr10050978
Guo X, Guo W, Yang M, Sun Y, Wang Y, Yan Y, Zhu B. Effect of Bacillus Additives on Fermentation Quality and Bacterial Community during the Ensiling Process of Whole-Plant Corn Silage. Processes. 2022; 10(5):978. https://0-doi-org.brum.beds.ac.uk/10.3390/pr10050978
Chicago/Turabian StyleGuo, Xiaojun, Wei Guo, Ming Yang, Yuelong Sun, Yujing Wang, Yan Yan, and Baocheng Zhu. 2022. "Effect of Bacillus Additives on Fermentation Quality and Bacterial Community during the Ensiling Process of Whole-Plant Corn Silage" Processes 10, no. 5: 978. https://0-doi-org.brum.beds.ac.uk/10.3390/pr10050978