Alpha Amylase from Bacillus pacificus Associated with Brown Algae Turbinaria ornata: Cultural Conditions, Purification, and Biochemical Characterization



Abstract
:1. Introduction
2. Results
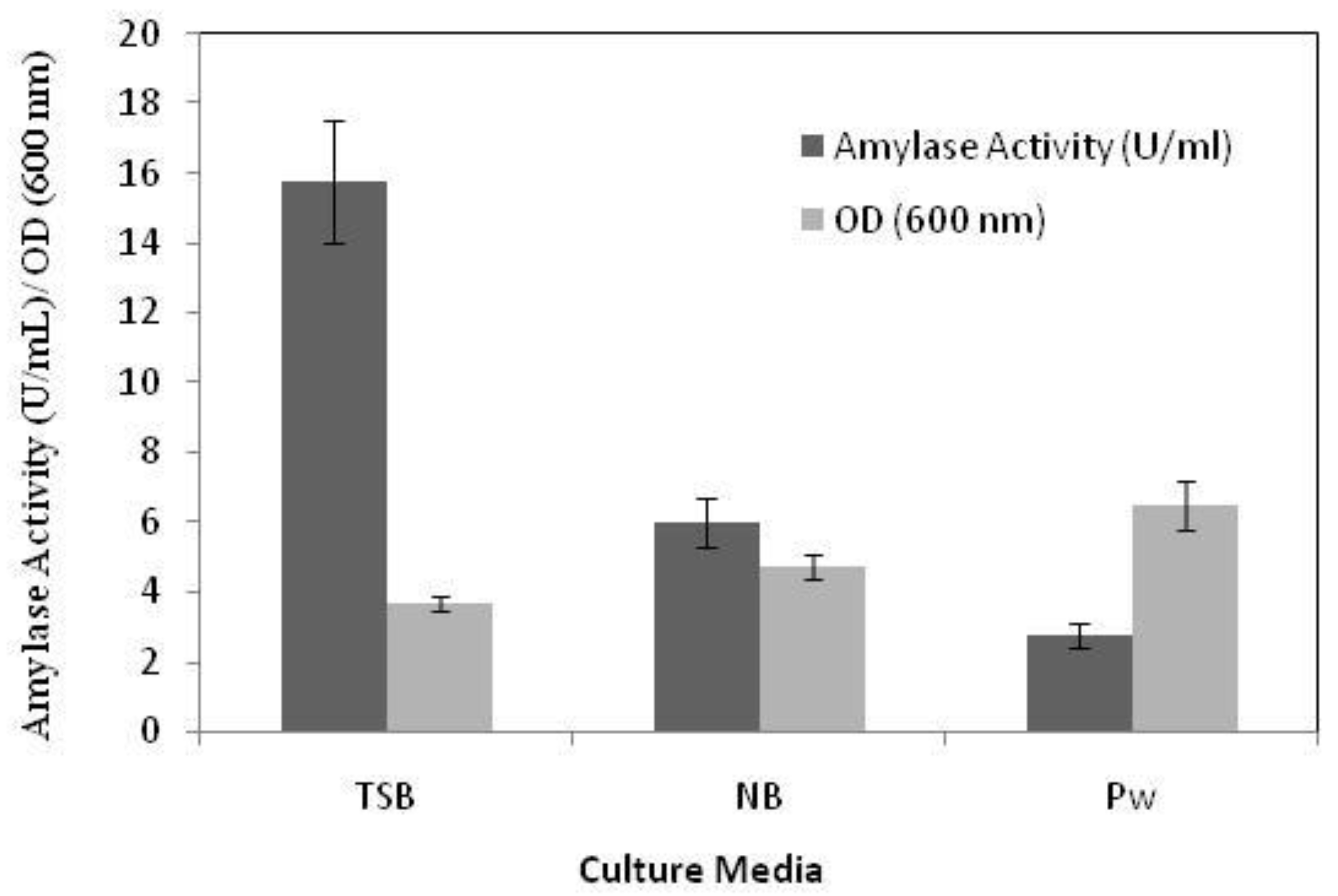
2.1. Cultivation Conditions of Marine Bacterial Amylase
Optimization of the Fermentation Media
2.2. Effect of Physical Parameters
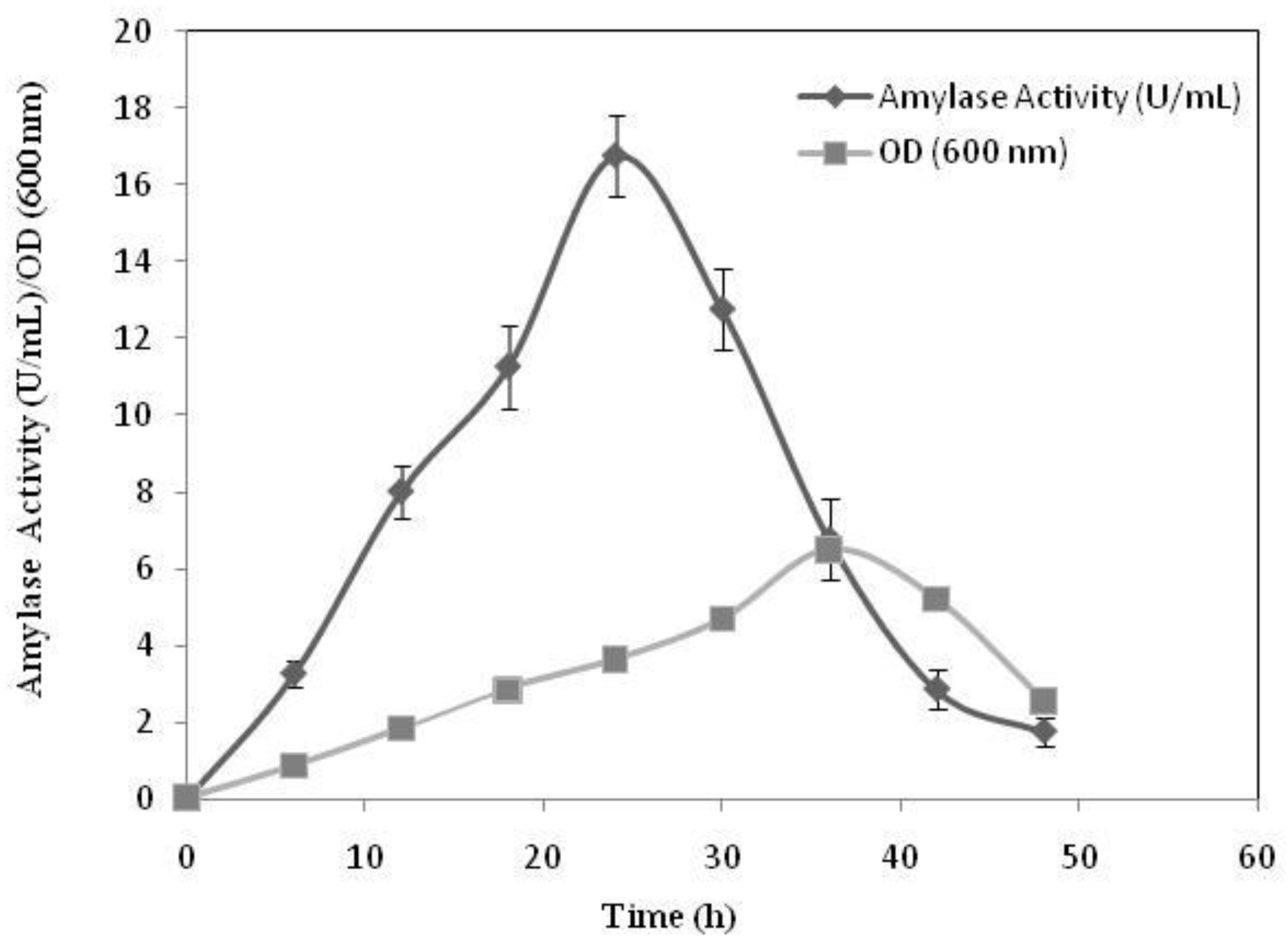
2.2.1. The Effect of Incubation Time on Cell Growth and Amylase Productivity
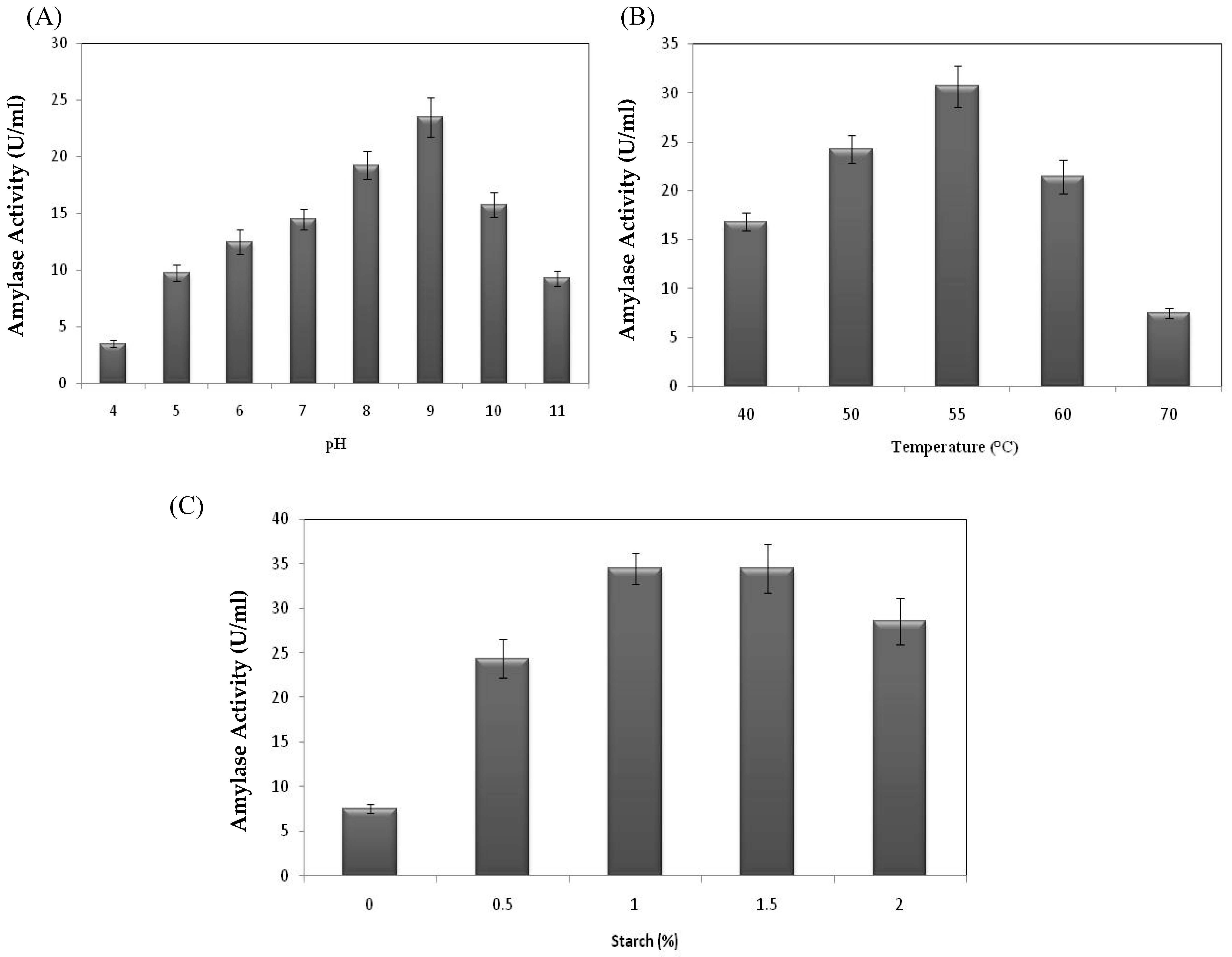
2.2.2. Effect of Initial pH, Temperature, and Substrate Concentration on Bacterial Amylase Activity
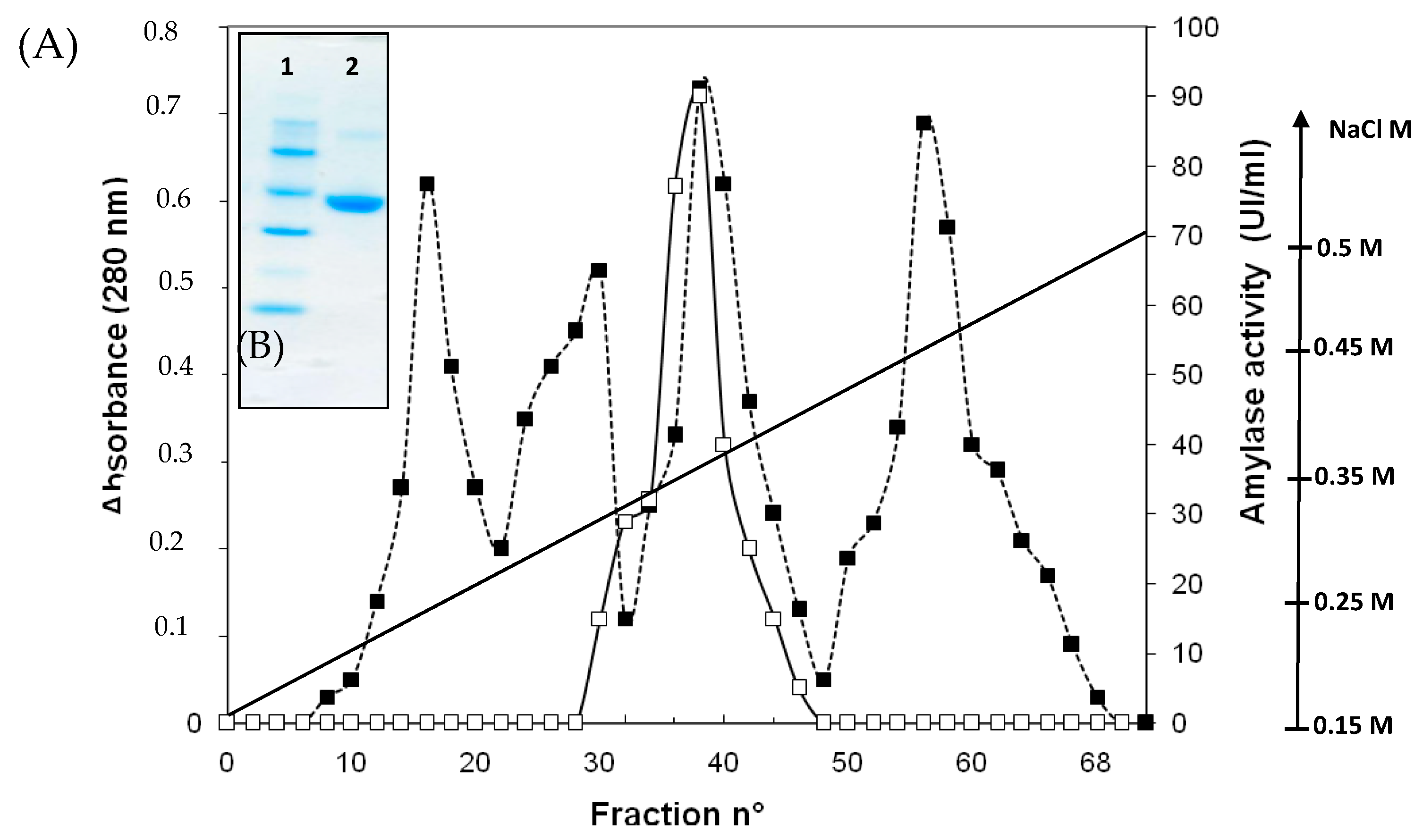
2.3. Purification of Amylase from Bacillus pacificus amB.p
2.4. Pure Amylase amB.p Characterization
The Effect of pH and Temperature on Amylase Activity and Stability
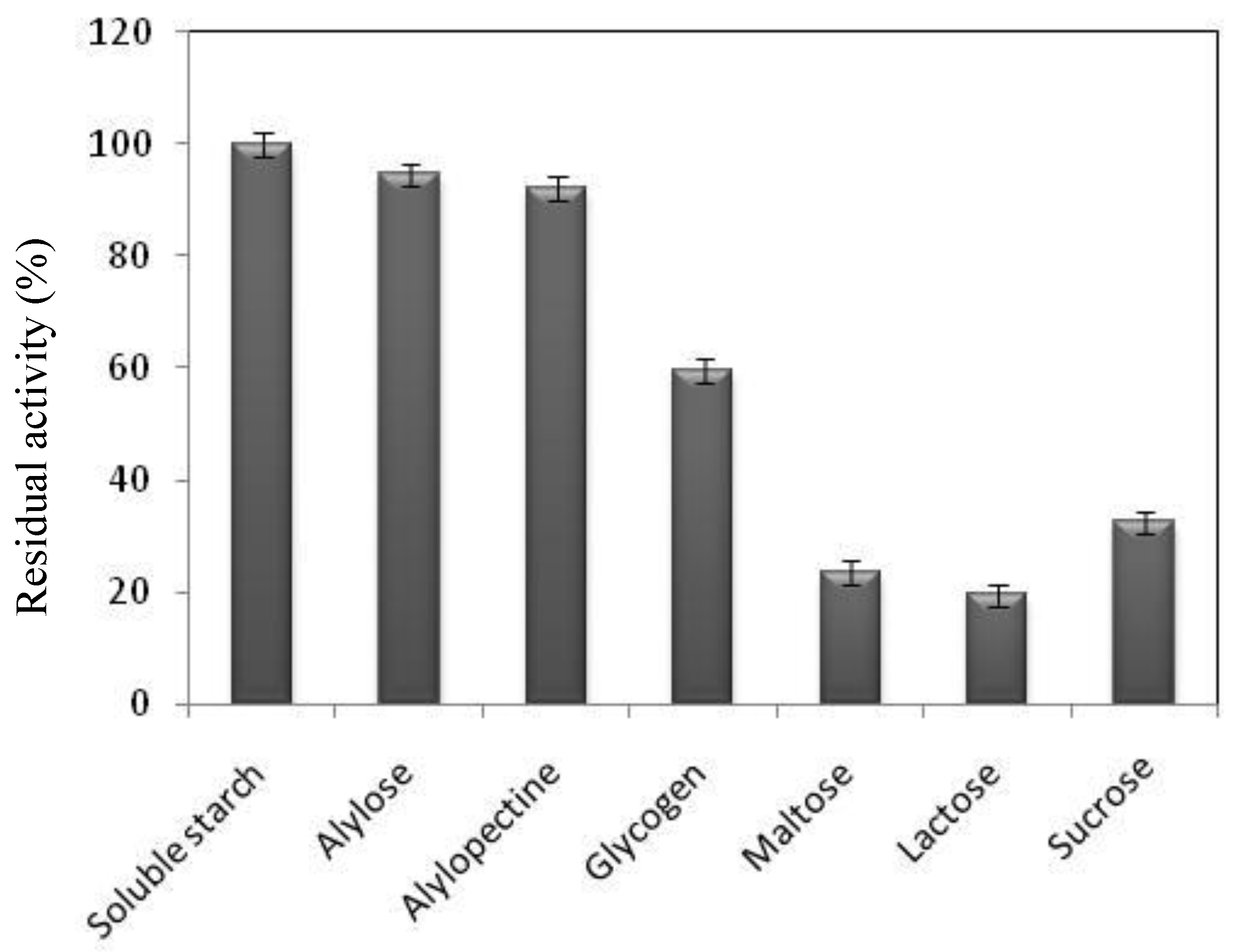
2.5. Substrate Specificity of B. pacificus α-Amylase amB.p
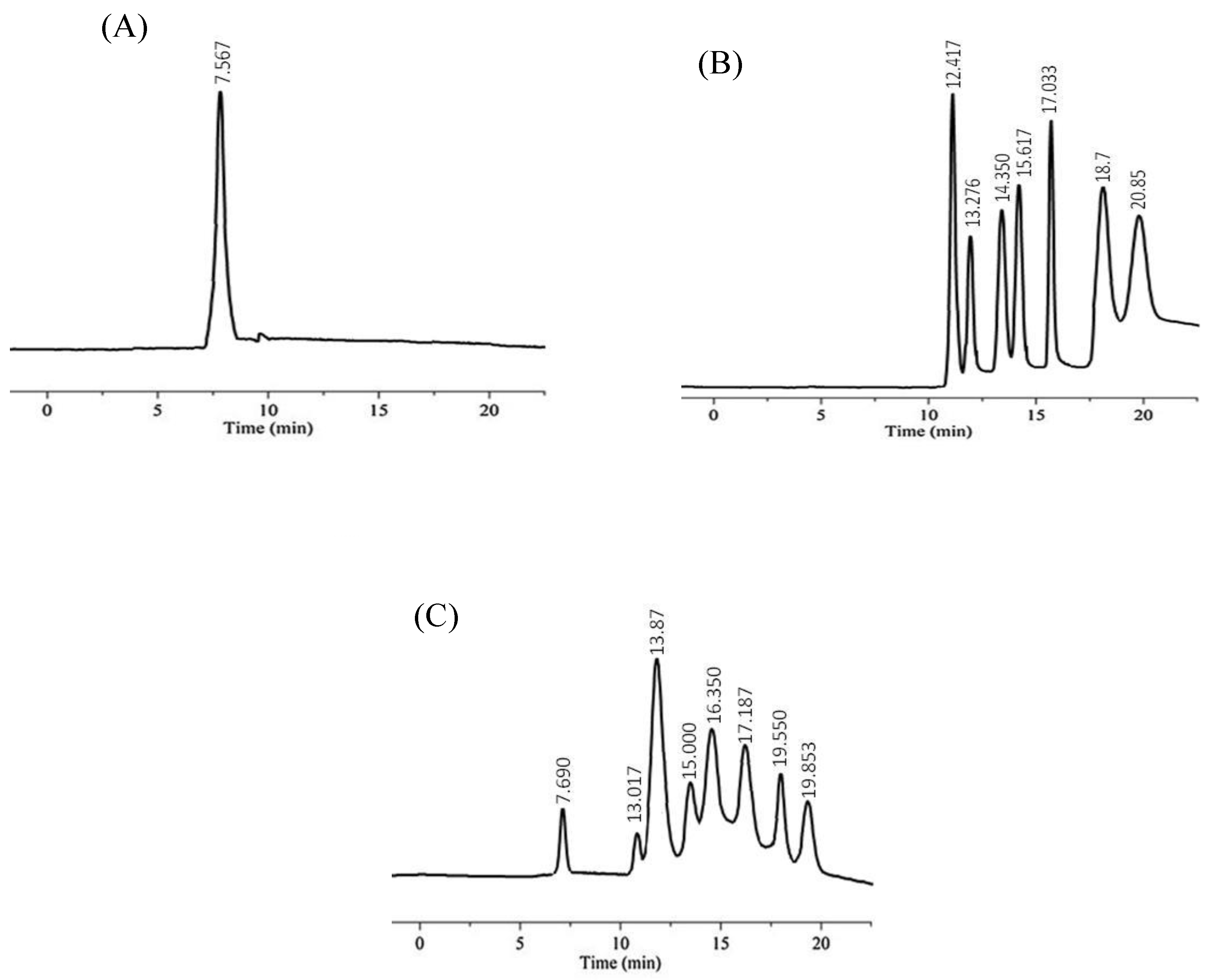
2.6. Analysis of the Starch Hydrolysis Products
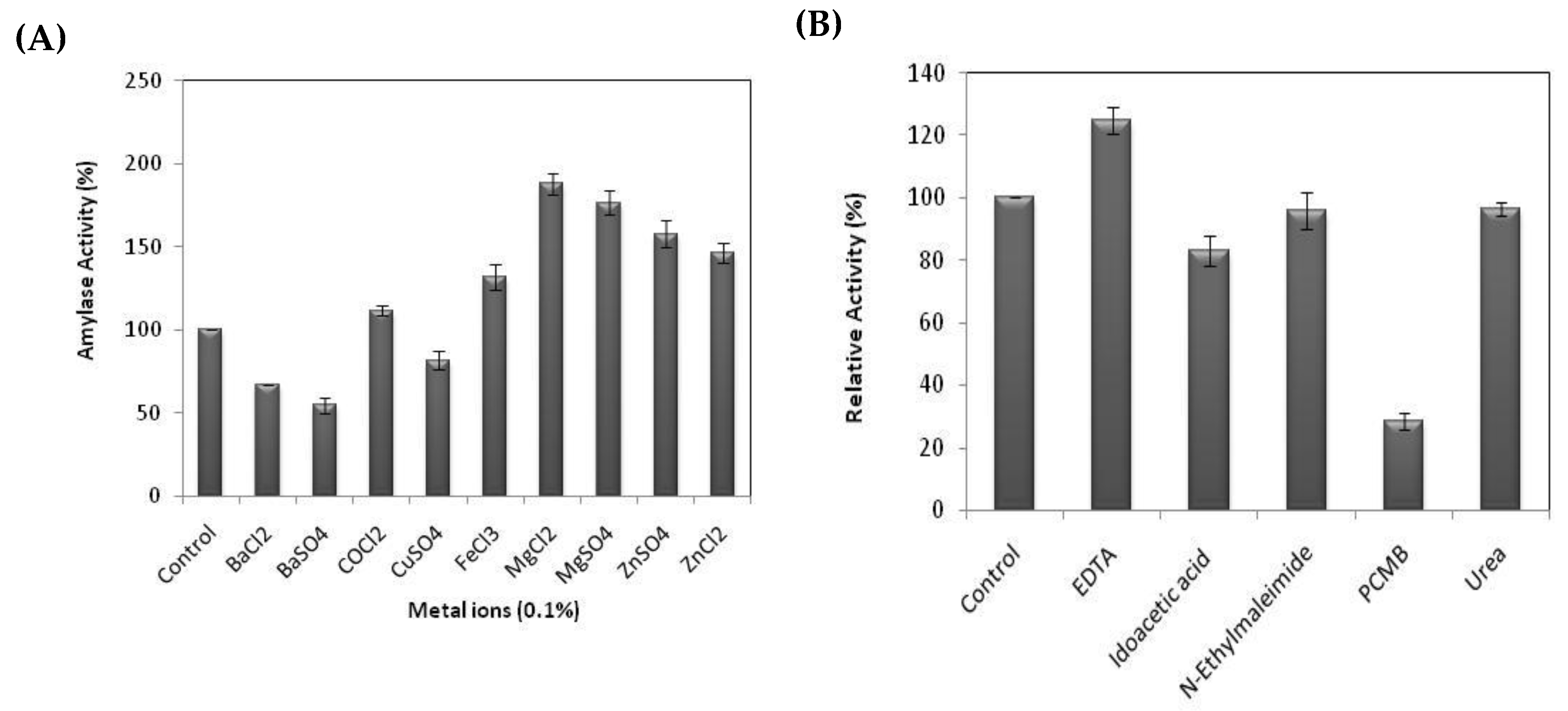
2.7. Effect of Metal Ions and Chemical Reagents on Pure α-Amylase Activity
3. Discussion
4. Materials and Methods
4.1. Microorganism and Cultivation Conditions
4.2. α-Amylase Assay
4.3. Inoculum Size
4.4. The Effect of Incubation Time
4.5. Influence of Initial pH and Temperature
4.6. Effect of Substrate Concentration
4.7. α-Amylase Purification
4.8. Protein Analysis
4.9. HPLC Analysis of the Starch Hydrolysis Products
4.10. Metal Ions and Chemical Reagents Effects on α-Amylase Activity
4.11. Substrate Specificity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statements
Acknowledgments
Conflicts of Interest
References
- Tester, R.F.; Karkalas, J.; Qi, X. Starch-composition, fine structure and architecture. J. Cereal Sci. 2004, 39, 151–165. [Google Scholar] [CrossRef]
- Gupta, R.; Gigras, P.; Mohapatra, H.; Goswami, V.K.; Chauhan, B. Microbial α-amylases: A biotechnological perspective. Process. Biochem. 2003, 38, 1599–1616. [Google Scholar] [CrossRef]
- Ficko-Blean, E.; Stuart, C.P.; Boraston, A.B. Structural analysis of CPF_2247, a novel α-amylase from Clostridium perfringens. Proteins 2011, 79, 2771–2777. [Google Scholar] [CrossRef] [PubMed]
- Sagu, S.T.; Huschek, G.; Bonick, J.; Homann, T.; Rawel, H.M. A New Approach of Extraction of alpha-Amylase/trypsin Inhibitors from Wheat (Triticum aestivum L.), Based on Optimization Using Plackett-Burman and Box-Behnken Designs. Molecules 2019, 24, 3589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratanakhanokchai, K.; Kaneko, J.; Kamio, Y.; Izaki, K. Purification and properties of a maltotetraose and maltotriose producing amylase from Chloroflexus aurantiacus. Appl. Environ. Microbiol. 1992, 58, 2490–2494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalpana, B.J.; Pandian, S.K. Halotolerant, acid-alkali stable, chelator resistant and raw starch digesting α-amylase from a marine bacterium Bacillus subtilis S8–18. J. Basic Microbiol. 2014, 8, 802–811. [Google Scholar] [CrossRef]
- Dou, S.; Chi, N.; Zhou, X.; Zhang, Q.; Pang, F.; Xiu, Z. Molecular cloning, expression, and biochemical characterization of a novel cold-active α-amylase from Bacillus sp. dsh19-1. Extremophiles 2018, 22, 739–749. [Google Scholar] [CrossRef]
- Puspasari, F.; Radjasa, O.K.; Noer, A.S.; Nurachman, Z.; Syah, Y.M.; Van Der Maarel, M.; Dijkhuizen, L.; Janeček, Š.; Natalia, D. Raw starch-degrading α-amylase from Bacillus aquimaris MKSC 6.2: Isolation and expression of the gene, bioinformatics and biochemical characterization of the recombinant enzyme. J. Appl. Microbiol. 2013, 114, 108–120. [Google Scholar] [CrossRef] [Green Version]
- Brown, E.D.; Yada, R.; Marangoni, A.G. The dependence of the lipolytic activity of Rhizopus arrhizus lipase on surfactant concentration in Aerosol-OT/isooctane reverse micelles and its relationship to enzyme structure. Biochim. Biophys. Acta 1993, 1161, 66–72. [Google Scholar] [CrossRef]
- Mobini, D.M.; Javann, A.F. Application of alpha-amylase in biotechnology. J. Biol. Today World 2012, 1, 39–50. [Google Scholar]
- Nisha, M.; Satyanarayana, T. Recombinant bacterial amylopellulanase developments ad perspectives. Bioingeneered 2013, 4, 388–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, L.; Chyau, C.C.; Hsu, W.H. Production and properties of a raw starch-degrading amylase from the thermophilic and alkaliphilic Bacillus sp. TS-23. Biotechnol. Appl. Biochem. 1998, 28, 61–68. [Google Scholar]
- Gomes, I.; Gomes, J.; Steiner, W. Highly thermostable amylase and pullulanase of the extreme thermophilic eubacterium Rhodothermus marinus: Production and partial characterization. Bioresour. Technol. 2003, 90, 207–214. [Google Scholar] [CrossRef]
- Saxena, R.K.; Dutt, K.; Agarwal, L.; Nayyar, P. A highly thermostable and alkaline amylase from a Bacillus spPN5. J. Biortech. 2007, 98, 260–265. [Google Scholar]
- Teodoro, C.E.D.S.; Martins, M.L.L. Culture conditions for the production of thermostable amylase by Bacillus sp. Braz. J. Microbiol. 2000, 31, 298–302. [Google Scholar] [CrossRef]
- Cripwell, R.A.; Van Zyl, W.H.; Viljoen-Bloom, M. Amylases and their applications. Afr. J. Biotechnol. 2005, 4, 1525–1529. [Google Scholar]
- Vijayaraghavan, P.; Kalaiyarasi, M.; Vincent, S.G.P. Cow dung is an ideal fermentation medium for amylase production in solid-state fermentation by Bacillus cereus. J. Genet. Eng. Biotechnol. 2015, 13, 111–117. [Google Scholar] [CrossRef] [Green Version]
- Asad, W.; Asif, M.; Rasool, S.A. Rasool. Extracellular enzyme production by indigenous thermophilic bacteria: Partial purification and characterization of α-amylase by Bacillus sp. WA21. Pak. J. Bot. 2011, 43, 1045–1052. [Google Scholar]
- Bajpai, P.; Bajpai, P.K. High-temperature alkaline α-amylase from Bacillus licheniformis TCRDC-B13. Biotech. Bioeng. 1989, 33, 72–78. [Google Scholar] [CrossRef]
- Pandey, A.; Nigam, P.S.N.; Soccol, C.R.; Soccol, V.T.; Singh, D.; Mohan, R. Advances in microbial amylases. Biotechnol. Appl. Biochem. 2000, 31, 135–152. [Google Scholar] [CrossRef]
- Normurodova, K.T.; Nurmatov, S.K.; Alimova, B.K.; Pulatova, O.M.; Akhmedova, Z.R.; Makhsumkhanov, A.A. Isolation and characteristics of highly active α-amylase from Bacillus subtilis-150. Chem. Nat. Comp. 2007, 43, 454–457. [Google Scholar] [CrossRef]
- Singh, R.N.; Bahuguna, A.; Chauhan, P.; Sharma, V.K.; Kaur, S.; Singh, S.K.; Khan, A. Production, purification and characterization of thermostable α-amylase from soil isolate Bacillus sp. strain B-10. J. Biosci. Biotechnol. 2016, 5, 37–43. [Google Scholar]
- Paul, J.S.; Lall, B.M.; Jadhav, S.K.; Tiwari, K.L. Parameter’s optimization and kinetics study of α-amylase enzyme of Bacillus sp. MB6 isolated from vegetable waste. Process. Biochem. 2017, 52, 123–129. [Google Scholar] [CrossRef]
- Haq, I.; Ali, S.; Javed, M.M.; Hameed, U.; Saleem, A.; Adnan, F.; Qadeer, M.A. Production of α amylase from a randomly induced mutant strain of Bacillus amyloliquefaciens and its application as a desizer in textile industry. Pak. J. Bot. 2010, 42, 473–484. [Google Scholar]
- Zambare, V. Optimization of amylase production from Bacillus sp. using statistics based experimental design. Emir. J. Food Agric. 2011, 23, 37–47. [Google Scholar] [CrossRef] [Green Version]
- Raplong, H.H.; Odeleye, P.O.; Chrinius, H. Production of Alpha Amylase by Bacillus cereus in Submerged Fermentation. J. Aceh Int. J. Sci. Technol. 2014, 3, 124–130. [Google Scholar] [CrossRef] [Green Version]
- Demirkan, E.S.; Mikami, B.; Adachi, M.; Higasa, T.; Utsumi, S. α-Amylase from B. amyloliquefaciens: Purification, characterization, raw starch degradation and expression in E. coli. Process. Biochem. 2005, 40, 2629–2636. [Google Scholar] [CrossRef]
- Kusuda, M.; Nagai, M.; Hur, T.-C.; Ueda, M.; Terashita, T. Purification and some properties of α-amylase from an ectomycorrhizal fungus, Tricholoma matsutake. Mycoscience 2003, 44, 311–317. [Google Scholar] [CrossRef]
- Mohamed, S.A.; Azhar, E.I.; Ba-Akdah, M.M.; Tashk, N.R.; Kumosani, T.A. Production, purification and characterization of α-amylase from Trichoderma harzianum grown on mandarin peel. Afr. J. Microbiol. Res. 2011, 5, 930–940. [Google Scholar]
- Busi, M.V.; Barchiesi, J.; Martín, M.; Gomez-Casati, D.F. Starch metabolism in green algae. Starch 2014, 66, 28–40. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Hewick, R.M.; Hunkapiller, M.W.; Hood, L.E.; Dreyer, W.J. A gas-liquid solid phase peptide and protein sequenator. J. Biol. Chem. 1981, 256, 7990–7997. [Google Scholar] [PubMed]
- Sarikaya, E.; Higasa, T.; Adachi, M.; Mikamib, B. Comparison of degradation abilities of a and b-amylases on raw starch granules. Process. Biochem. 2000, 35, 711–715. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Step | Total Activity (Units) | Protein (Mg) | Specific Activity (U/Mg) | Activity Recovery (%) | Purification Factor |
|---|---|---|---|---|---|
| Crude extract | 6210 | 477.7 | 13 | 100 | 1 |
| Heat treatment (70 °C, 10 min) | 4937 | 133.4 | 37 | 79.5 | 2.8 |
| Ammonium sulfate Fractionation (40–85%) | 3160 | 58.5 | 54 | 58 | 4.1 |
| Mono Q-Sephadex | 1585 | 15.1 | 105 | 25.5 | 8.07 |
| DAILHAFNWKYSRVTANAEQKAAAG (Present Work) |
|---|
| DAIVHAFNWKYSRVTANAEHKAAAG (α-amylase from Tepidimonas fonticaldi QBN20693.1) |
| DAILHAFNWKYSDVTANAEHIAAAG (α-amylase from unclassified Vibrio WP161425494.1) DAILHAFNWKYSDVTANAEHIAAAG (α-amylase from Vibrio sp. A8-1 WP_170905550.1) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alonazi, M.; Karray, A.; Badjah-Hadj-Ahmed, A.Y.; Ben Bacha, A. Alpha Amylase from Bacillus pacificus Associated with Brown Algae Turbinaria ornata: Cultural Conditions, Purification, and Biochemical Characterization. Processes 2021, 9, 16. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9010016
Alonazi M, Karray A, Badjah-Hadj-Ahmed AY, Ben Bacha A. Alpha Amylase from Bacillus pacificus Associated with Brown Algae Turbinaria ornata: Cultural Conditions, Purification, and Biochemical Characterization. Processes. 2021; 9(1):16. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9010016
Chicago/Turabian StyleAlonazi, Mona, Aida Karray, Ahmed Yacine Badjah-Hadj-Ahmed, and Abir Ben Bacha. 2021. "Alpha Amylase from Bacillus pacificus Associated with Brown Algae Turbinaria ornata: Cultural Conditions, Purification, and Biochemical Characterization" Processes 9, no. 1: 16. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9010016