
Palm Oil Decanter Cake Wastes as Alternative Nutrient Sources and Biomass Support Particles for Production of Fungal Whole-Cell Lipase and Application as Low-Cost Biocatalyst for Biodiesel Production

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Decanter Cake Waste from Palm Oil Mill
2.2. Fungal Strains
2.3. Selection of Whole-Cell Lipase Producing Fungi
2.4. Optimization of DCW-Based Nutrient Composition and Culture Conditions for SSF
2.5. Optimization of Whole-Cell Lipase Production
2.6. Application of Whole-Cell Lipase as a Biocatalyst for Biodiesel Production
2.6.1. Standard Procedures for Determination of Lipase Activity
Determination of Hydrolytic Activity
Determination of Transesterification Activity
Determination of Esterification Activity
Analysis of FAME by TLC-FID
2.6.2. Optimization of Biodiesel Production by Whole-Cell Biocatalyst
2.6.3. Scale up of Whole-Cell Lipase and Biodiesel Production
2.7. Analytical Methods
2.8. Statistical Analysis
3. Results and Discussion
3.1. Selection of Whole-Cell Lipase Producing Fungi
3.2. Optimization of Medium Component and Culture Conditions for Whole-Cell Lipase Production
3.2.1. Optimization of Medium Components
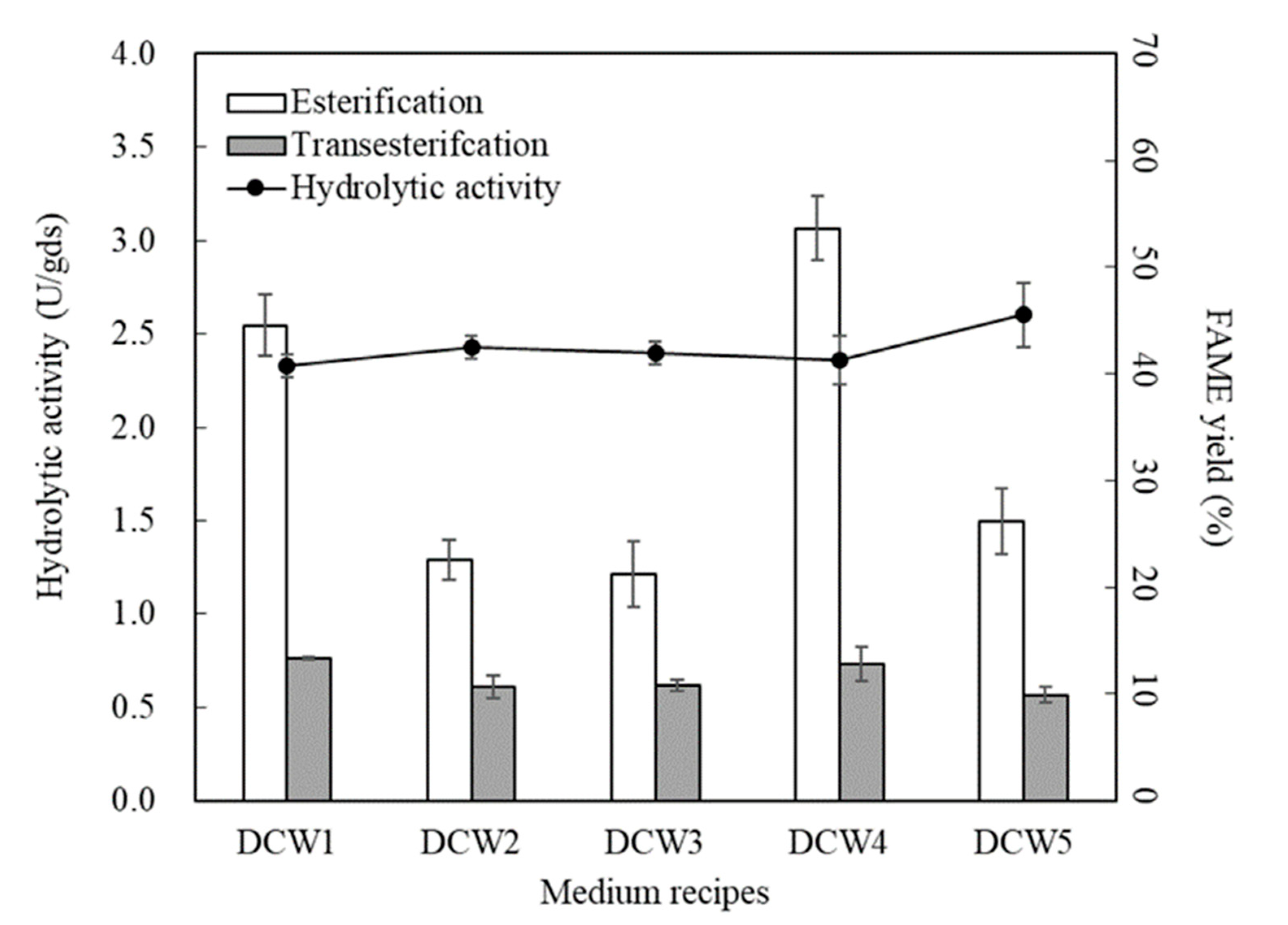
3.2.2. Addition of Co-Carbon and Inorganic Nitrogen Sources
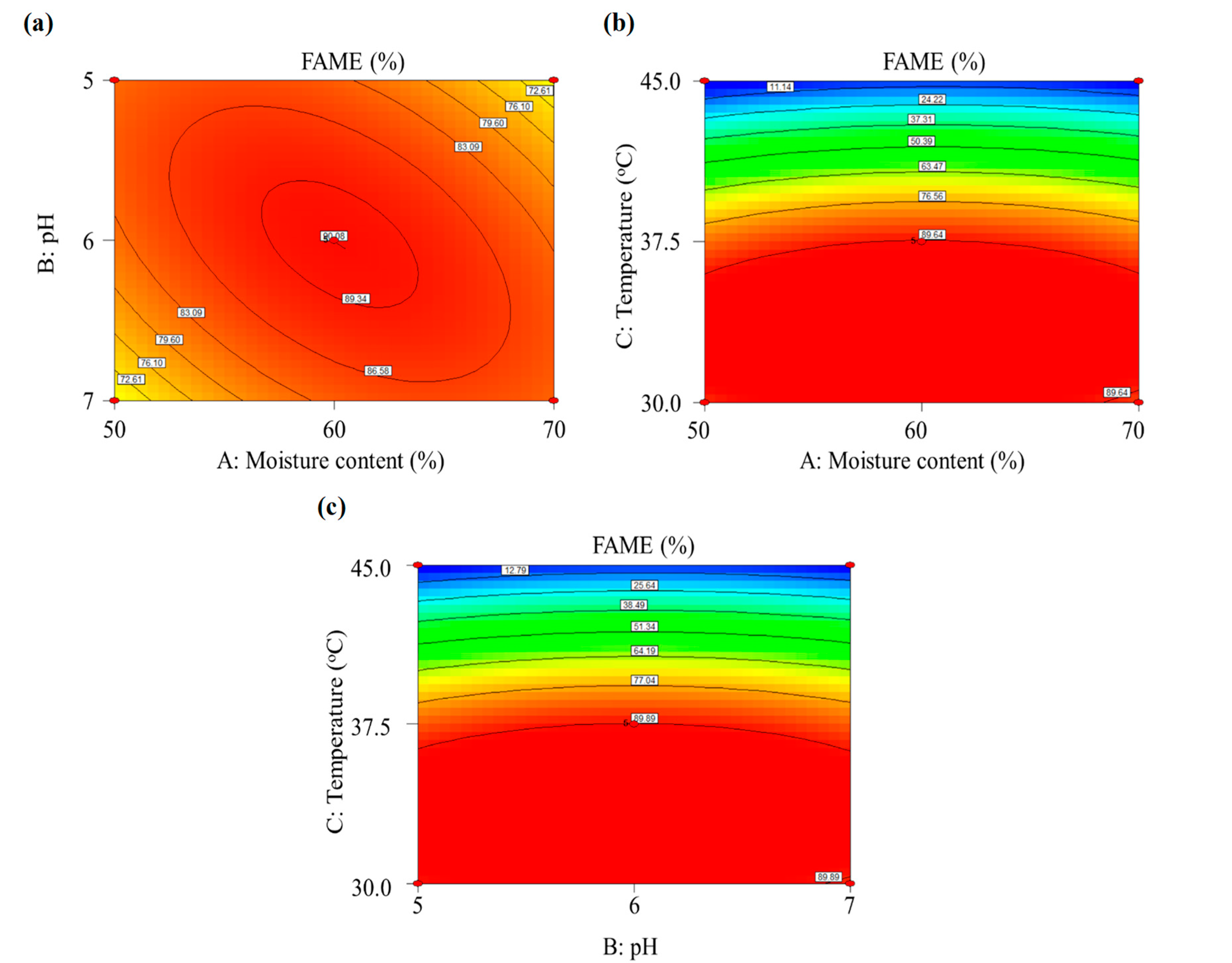
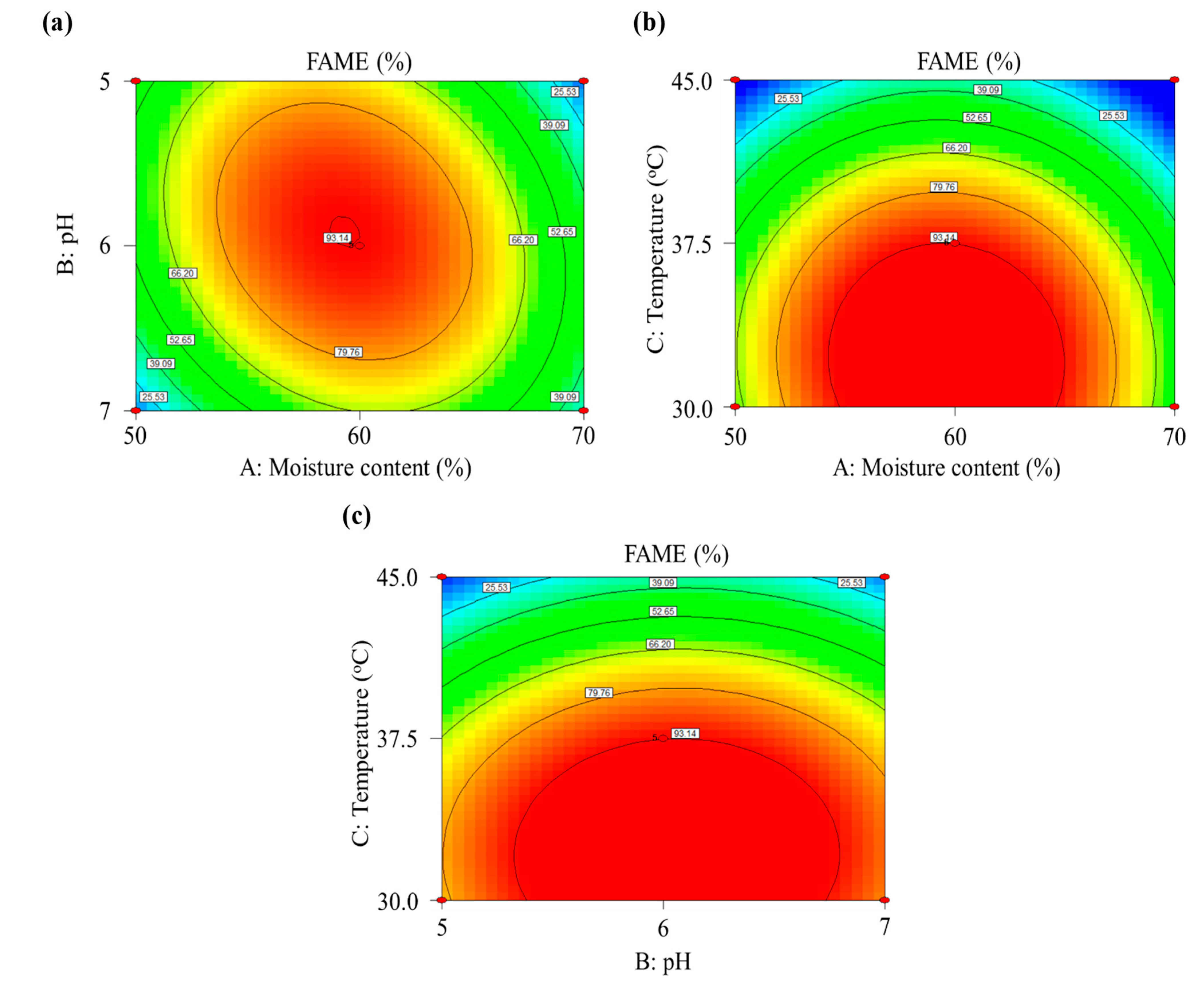
3.2.3. Optimization of SSF through Response Surface Methodology
3.3. Optimization of Biodiesel Production Using Whole-Cell Lipase
3.3.1. Effect of Reaction Temperature
3.3.2. Optimization of Water Content and Methanol Molar Ratio in Esterification Reaction
3.3.3. Optimization of Water Content and Methanol Molar Ratio in Transesterification Reaction
3.4. Selection of Whole-Cell Lipase Producing Fungi
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rakchai, N.; H-Kittikun, A.; Zimmermann, W. The production of immobilized whole-cell lipase from Aspergillus nomius ST57 and the enhancement of the synthesis of fatty acid methyl esters using a two-step reaction. J. Mol. Catal. B Enzym. 2016, 133, 128–136. [Google Scholar] [CrossRef]
- Rakchai, N.; H-Kittikun, A.; Zimmermann, W. Production of whole-cell lipase from Aspergillus nomius ST57 and optimization of methyl esters synthesis from palm oil in one step. Chiang Mai J. Sci. 2018, 45, 746–761. [Google Scholar]
- Jegannathan, K.R.; Abang, S.; Poncelet, D.; Chan, E.S.; Ravindra, P. Production of biodiesel using immobilized lipase–A critical review. Crit. Rev. Biotechnol. 2008, 28, 253–264. [Google Scholar] [CrossRef]
- Stergiou, P.Y.; Foukis, A.; Filippou, M.; Koukouritaki, M.; Parapouli, M.; Theodorou, L.G.; Hatziloukas, E.; Afendra, A.M.; Pandey, A.; Papamichael, E. Advances in lipase-catalyzed esterification reactions. Biotechnol. Adv. 2013, 31, 1846–1859. [Google Scholar] [CrossRef] [PubMed]
- Ungcharoenwiwat, P.; H-Kittikun, A. Purification and characterization of lipase from Burkholderia sp. EQ3 isolated from wastewater from a canned fish factory and its application for the synthesis of wax esters. J. Mol. Catal. B Enzym. 2015, 115, 96–104. [Google Scholar] [CrossRef]
- Katayama, M.; Kuroiwa, T.; Suzuno, K.; Igusa, A.; Matsui, T.; Kanazawa, A. Hydration-aggregation pretreatment for drastically improving esterification activity of commercial lipases in non-aqueous media. Enzyme Microb. Technol. 2017, 105, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Koutinas, M.; Yiangou, C.; Osório, N.M.; Ioannou, K.; Canet, A.; Valero, F.; Ferreira Dias, S. Application of commercial and non-commercial immobilized lipases for biocatalytic production of ethyl lactate in organic solvents. Bioresour. Technol. 2018, 247, 496–503. [Google Scholar] [CrossRef]
- Kajiwara, S.; Komatsu, K.; Yamada, R.; Matsumoto, T.; Yasuda, M.; Ogino, H. Improvement of the organic solvent stability of a commercial lipase by chemical modification with dextran. Biochem. Eng. J. 2019, 142, 1–6. [Google Scholar] [CrossRef]
- Regner, E.L.; Salvatierra, H.N.; Baigorí, M.D.; Pera, L.M. Biomass-bound biocatalysts for biodiesel production: Tuning a lipolytic activity from Aspergillus niger MYA 135 by submerged fermentation using agro-industrial raw materials and waste products. Biomass Bioenergy 2019, 120, 59–67. [Google Scholar] [CrossRef]
- Razak, M.N.A.; Ibrahim, M.F.; Yee, P.L.; Hassan, M.A.; Abd-Aziz, S. Utilization of oil palm decanter cake for cellulase and polyoses production. Biotechnol. Bioprocess Eng. 2012, 17, 547–555. [Google Scholar] [CrossRef]
- Thamvithayakorn, P.; Phosri, C.; Pisutpaisal, N.; Krajangsang, S.; Whalley, A.J.; Suwannasai, N. Utilization of oil palm decanter cake for valuable laccase and manganese peroxidase enzyme production from a novel white-rot fungus, Pseudolagarobasidium sp. 3 Biotech 2019, 9, 17–33. [Google Scholar] [CrossRef]
- Oliveira, A.C.; Amorim, G.M.; Azevêdo, J.A.G.; Godoy, M.G.; Freire, D.M.G. Solid-state fermentation of co-products from palm oil processing: Production of lipase and xylanase and effects on chemical composition. Biocatal. Biotrans. 2018, 36, 381–388. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 16th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1999. [Google Scholar]
- Mazaheri, H.; Lee, K.T.; Bhatia, S.; Mohamed, A.R. Subcritical water liquefaction of oil palm fruit press fiber in the presence of sodium hydroxide: An optimisation study using response surface methodology. Bioresour. Technol. 2010, 101, 9335–9341. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Rhee, J.S. Production and partial purification of a lipase from Pseudomonas putida 3SK. Enzyme Microb. Technol. 1993, 15, 617–623. [Google Scholar] [CrossRef]
- Hama, S.; Tamalampudi, S.; Fukumizu, T.; Miura, K.; Yamaji, H.; Kondo, A.; Fukuda, H. Lipase localization in Rhizopus oryzae cells immobilized within biomass support particles for use as whole-cell biocatalysts in biodiesel-fuel production. J. Biosci. Bioeng. 2006, 101, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Fedosov, S.N.; Brask, J.; Xu, X. Analysis of biodiesel conversion using thin layer chromatography and nonlinear calibration curves. J. Chromatogr. A 2011, 1218, 2785–2792. [Google Scholar] [CrossRef] [PubMed]
- Lima, V.M.G.; Krieger, N.; Sarquis, M.I.M.; Mitchell, D.A.; Ramos, L.P.; Fontana, J.D. Effect of nitrogen and carbon sources on lipase production by Penicillium aurantiogriseum. Food Technol. Biotechnol. 2003, 41, 105–110. [Google Scholar]
- Tanyol, M.; Uslu, G.; Yönten, V. Optimization of lipase production on agro-industrial residue medium by Pseudomonas fluorescens (NRLL B-2641) using response surface methodology. Biotechnol. Biotechnol. Equip. 2015, 29, 64–71. [Google Scholar] [CrossRef]
- Helal, S.E.; Abdelhady, H.M.; Abou-Taleb, K.A.; Hassan, M.G.; Amer, M.M. Evaluation of factors affecting the fungal lipase production using one factor at a time approach and response surface methodology. Egypt. J. Microbiol. 2017, 52, 1–16. [Google Scholar] [CrossRef]
- Oliveira, F.; Salgado, J.M.; Abrunhosa, L.; Perez-Rodrigues, N.; Dominguez, J.M.; Venancio, A.; Belo, I. Optimization of lipase production by solid-state fermentation of olive pomace: From flask to laboratory-scale packed-bed bioreactor. Bioprocess Biosyst. Eng. 2017, 40, 1123–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geoffry, K.; Achur, R.N. Screening and production of lipase from fungal organisms. Biocatal. Agric. Biotechnol. 2018, 14, 241–253. [Google Scholar] [CrossRef]
- Kitcha, S.; Cheirsilp, B. Bioconversion of lignocellulosic palm byproducts into enzymes and lipid by newly isolated oleaginous fungi. Biochem. Eng. J. 2014, 88, 95–100. [Google Scholar] [CrossRef]
- Gutarra, M.L.E.; Godoy, M.G.; Maugeri, F.; Rodrigues, M.I.; Freire, D.M.G.; Castilho, L.R. Production of an acidic and thermostable lipase of the mesophilic fungus Penicillium simplicissimum by solid-state fermentation. Bioresour. Technol. 2009, 100, 5249–5254. [Google Scholar] [CrossRef]
- Colla, L.M.; Primaz, A.L.; Benedetti, S.; Loss, R.A.; de Lima, M.; Reinehr, C.O.; Bertolin, T.E.; Costa, J.A. Surface response methodology for the optimization of lipase production under submerged fermentation by filamentous fungi. Braz. J. Microbiol. 2016, 47, 461–467. [Google Scholar] [CrossRef] [Green Version]
- Rakkan, T.; Suwanno, S.; Paichid, N.; Yunu, T.; Klomklao, S.; Sangkharak, K. Optimized synthesis method for transesterification of residual oil from palm oil mill effluent and lipase from Pacific white shrimp (Litopenaeus vannamei) hepatopancreas to environmentally friendly biodiesel. Fuel 2017, 209, 309–314. [Google Scholar] [CrossRef]
- Kuepethkaew, S.; Sangkharak, K.; Benjakul, S.; Klomklao, S. Optimized synthesis of biodiesel using lipase from Pacific white shrimp (Litopenaeus vannamei) hepatopancreas. Renew. Energy 2017, 104, 139–147. [Google Scholar] [CrossRef]
- Barahona, D.; Pfromm, P.H.; Rezac, M.E. Effect of water activity on the lipase catalyzed esterification of geraniol in ionic liquid [bmim]PF6. Biotechnol. Bioeng. 2006, 93, 318–324. [Google Scholar] [CrossRef]
- Márkus, Z.; Bélafi-bakó, K.; Tóth, G.; Nemestóthy, N.; Gubicza, L. Effect of chain length and order of the alcohol on enzyme activity during enzymatic esterification in organic media. Hung. J. Ind. Chem. 2017, 45, 35–39. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.C.; Liang, S.-H.; Chen, S.-S.; Su, C.-H.; Lin, J.-H.; Chien, C.-C. Enzymatic production of biodiesel from insect fat using methyl acetate as an acyl acceptor: Optimization by using response surface methodology. Energy Convers. Manag. 2018, 158, 168–175. [Google Scholar] [CrossRef]
- Matsumoto, M.; Odachi, D.; Kondo, K. Effect of water activity on rate of esterification by lipase in organic media. J. Chem. Eng. Jpn. 2001, 34, 437–440. [Google Scholar] [CrossRef]
- Noureddini, H.; Gao, X.; Philkana, R.S. Immobilized Pseudomonas cepacia lipase for biodiesel fuel production from soybean oil. Bioresour. Technol. 2005, 96, 769–777. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Sun, S.; Xin, Z.; Sheng, B.; Liu, Q. Synthesis and component confirmation of biodiesel from palm oil and dimethyl carbonate catalyzed by immobilized-lipase in solvent-free system. Fuel 2010, 89, 3960–3965. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Run | X1: Moisture Content (%) | X2: pH | X3: Temperature (°C) | Methyl Ester (%) | |||
|---|---|---|---|---|---|---|---|
| Esterification | Transesterification | ||||||
| Predicted | Actual | Predicted | Actual | ||||
| 1 | 50 | 5 | 37.5 | 69.11 | 67.12 | 15.31 | 18.42 |
| 2 | 70 | 5 | 37.5 | 83.36 | 83.84 | 30.96 | 20.13 |
| 3 | 50 | 7 | 37.5 | 82.68 | 82.21 | 47.26 | 58.06 |
| 4 | 70 | 7 | 37.5 | 69.39 | 71.39 | 11.99 | 8.85 |
| 5 | 50 | 6 | 30 | 88.70 | 89.24 | 63.09 | 46.01 |
| 6 | 70 | 6 | 30 | 87.17 | 85.23 | 57.39 | 54.24 |
| 7 | 50 | 6 | 45 | 0 | 0 | 0 | 0 |
| 8 | 70 | 6 | 45 | 0 | 0 | 0 | 0 |
| 9 | 60 | 5 | 30 | 90.30 | 91.77 | 77.78 | 91.71 |
| 10 | 60 | 7 | 30 | 88.39 | 88.34 | 83.09 | 89.33 |
| 11 | 60 | 5 | 45 | 0 | 0 | 6.27 | 0 |
| 12 | 60 | 7 | 45 | 1.46 | 0 | 13.95 | 0 |
| 13 | 60 | 6 | 37.5 | 90.07 | 90.32 | 93.04 | 92.47 |
| 14 | 60 | 6 | 37.5 | 90.07 | 90.69 | 93.04 | 92.79 |
| 15 | 60 | 6 | 37.5 | 90.07 | 91.03 | 93.04 | 93.95 |
| 16 | 60 | 6 | 37.5 | 90.07 | 91.46 | 93.04 | 92.59 |
| 17 | 60 | 6 | 37.5 | 90.07 | 86.89 | 93.04 | 93.33 |
| Factors | Methyl Ester (%) |
|---|---|
| Optimization of esterification reaction | |
| Effect of temperature (°C) | |
| 30 | 90.08 ± 1.83 |
| 37 | 90.72 ± 1.84 |
| 45 | 83.53 ± 2.05 |
| Effect of water content (aw) | |
| 0.71 | 90.72 ± 1.84 |
| 0.79 | 82.13 ± 5.19 |
| 0.83 | 83.82 ± 2.35 |
| 0.85 | 83.01 ± 1.87 |
| 0.87 | 76.38 ± 2.33 |
| 0.90 | 72.65 ± 3.60 |
| Effect of molar ratio of methanol and oleic acid | |
| 1:1 | 90.72 ± 1.84 |
| 2:1 | 96.25 ± 0.67 |
| 3:1 | 81.30 ± 0.37 |
| Optimization of transesterification reaction | |
| Effect of temperature (°C) | |
| 30 | 93.03 ± 0.61 |
| 37 | 94.70 ± 0.60 |
| 45 | 72.16 ± 4.35 |
| Effect of water content (aw) | |
| 0.73 | 94.70 ± 0.60 |
| 0.81 | 95.74 ± 0.28 |
| 0.85 | 90.16 ± 2.09 |
| 0.87 | 87.11 ± 1.98 |
| 0.89 | 83.38 ± 4.67 |
| 0.91 | 31.12 ± 3.25 |
| Effect of molar ratio of methanol to oil | |
| 3:1 | 95.74 ± 0.28 |
| 3.5:1 | 87.31 ± 1.27 |
| 4:1 | 27.89 ± 6.06 |
| 5:1 | 7.60 ± 3.57 |
| 6:1 | 3.37 ± 0.97 |
| Properties | Diesel ASTM D975 * | ASTM D6751 ** | EN 14214 *** | Produced Biodiesel |
|---|---|---|---|---|
| Methyl ester (%wt) | N.A. | N.A. | ≥96.5 | 94–97 |
| Viscosity at 40 °C (mm2/s) | 1.3 to 4.1 | 1.9 to 6.0 | 3.5 to 5.0 | 5.99 |
| Flash point (°C) | >52 | ≥130 | ≥120 | 175 |
| Copper strip corrosion | N.A. | ≤No. 3 | ≤No. 1 | No.1a |
| Cloud point (°C) | −15 to 5 | N.A. | N.A. | 11 |
| Pour point (°C) | −35 to 15 | N.A. | N.A. | 8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
H-Kittikun, A.; Cheirsilp, B.; Sohsomboon, N.; Binmarn, D.; Pathom-aree, W.; Srinuanpan, S. Palm Oil Decanter Cake Wastes as Alternative Nutrient Sources and Biomass Support Particles for Production of Fungal Whole-Cell Lipase and Application as Low-Cost Biocatalyst for Biodiesel Production. Processes 2021, 9, 1365. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9081365
H-Kittikun A, Cheirsilp B, Sohsomboon N, Binmarn D, Pathom-aree W, Srinuanpan S. Palm Oil Decanter Cake Wastes as Alternative Nutrient Sources and Biomass Support Particles for Production of Fungal Whole-Cell Lipase and Application as Low-Cost Biocatalyst for Biodiesel Production. Processes. 2021; 9(8):1365. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9081365
Chicago/Turabian StyleH-Kittikun, Aran, Benjamas Cheirsilp, Natthapat Sohsomboon, Darun Binmarn, Wasu Pathom-aree, and Sirasit Srinuanpan. 2021. "Palm Oil Decanter Cake Wastes as Alternative Nutrient Sources and Biomass Support Particles for Production of Fungal Whole-Cell Lipase and Application as Low-Cost Biocatalyst for Biodiesel Production" Processes 9, no. 8: 1365. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9081365