
Food-Derived Bioactive Peptides with Antioxidative Capacity, Xanthine Oxidase and Tyrosinase Inhibitory Activity


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. DPPH Assay
2.3. ABTS Assay
2.4. Reducing Power Assay
2.5. In Silico Analysis of 11 Bioactive Peptides (BPs)
2.6. XOD Inhibition Assay
2.7. Tyrosinase Inhibition Assay
2.8. Statistical Analysis
3. Results and Discussion
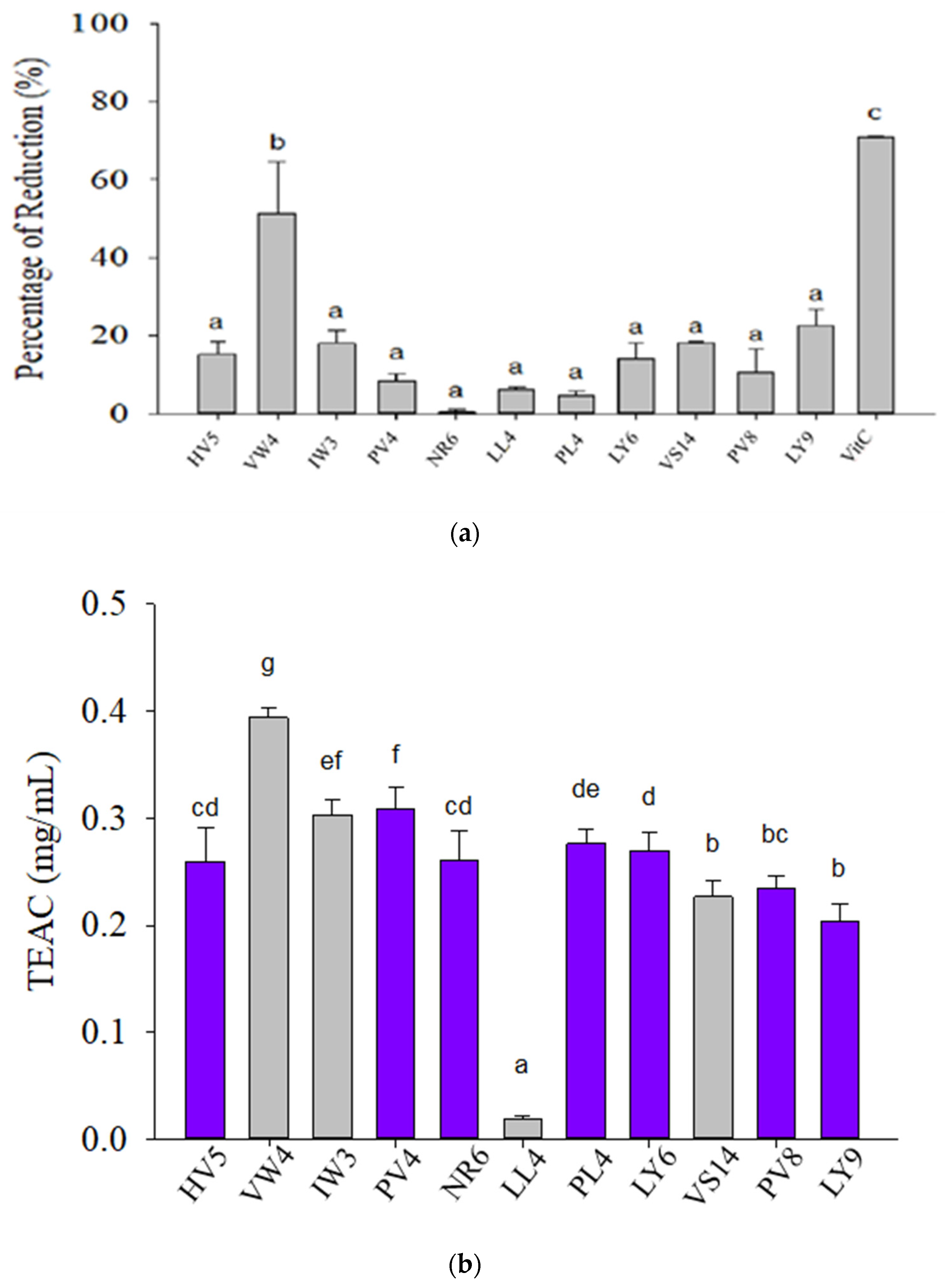
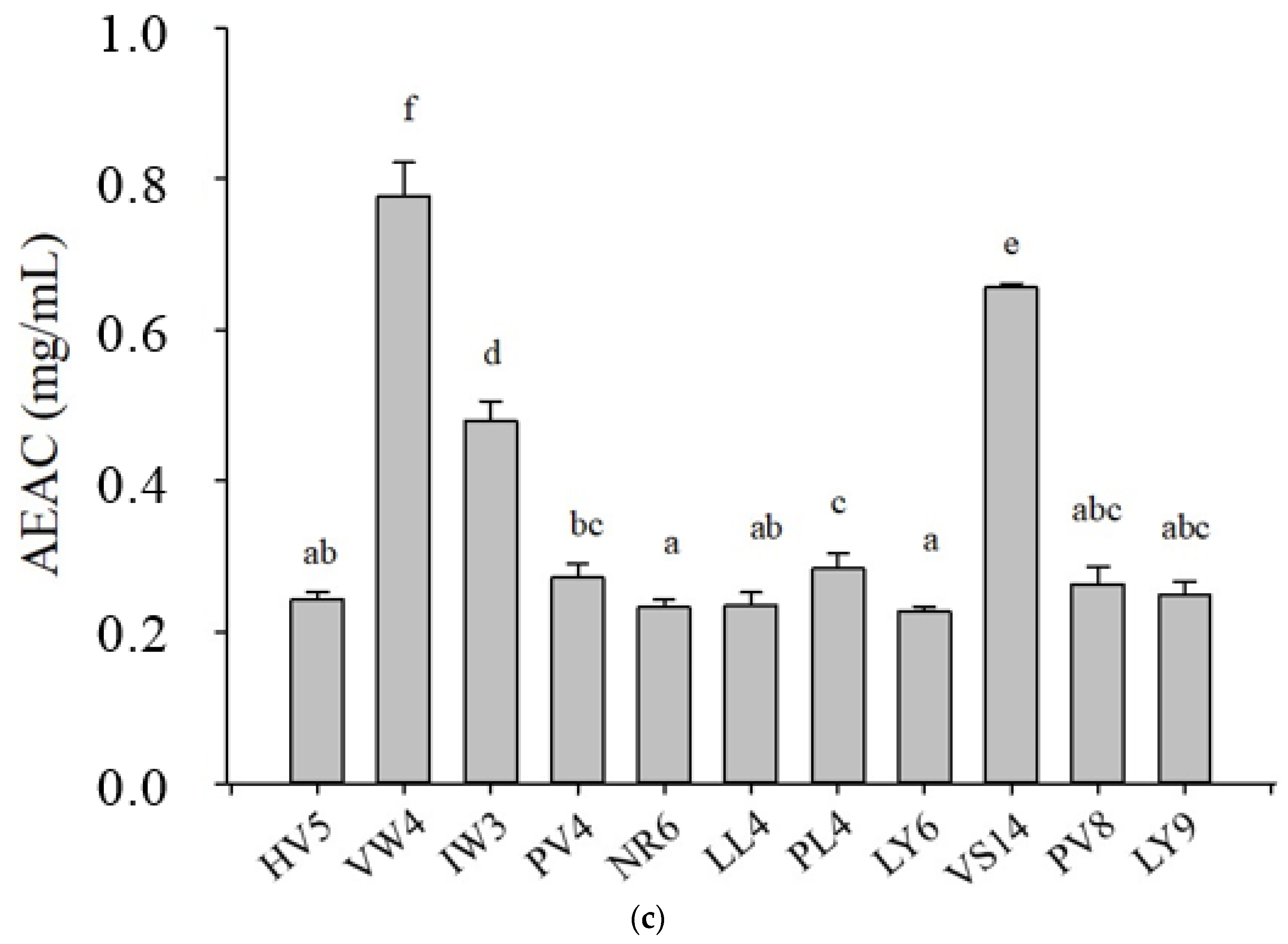
3.1. Antioxidant Activities of 11 Selected BPs
3.2. In Silico Analysis of Biological Activities of 11 Selected BPs
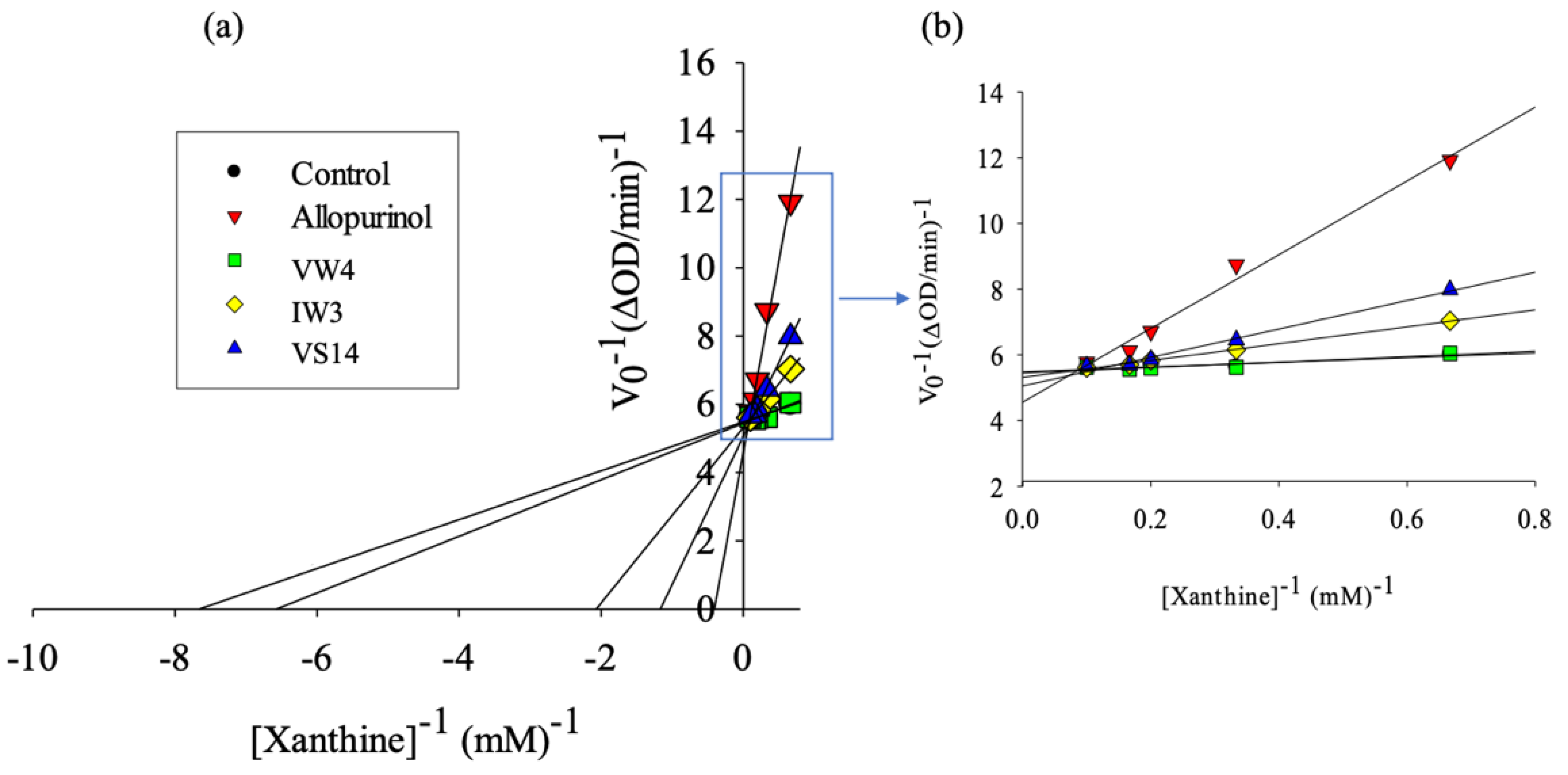
3.3. XOD Inhibitory Activity of Selected BPs
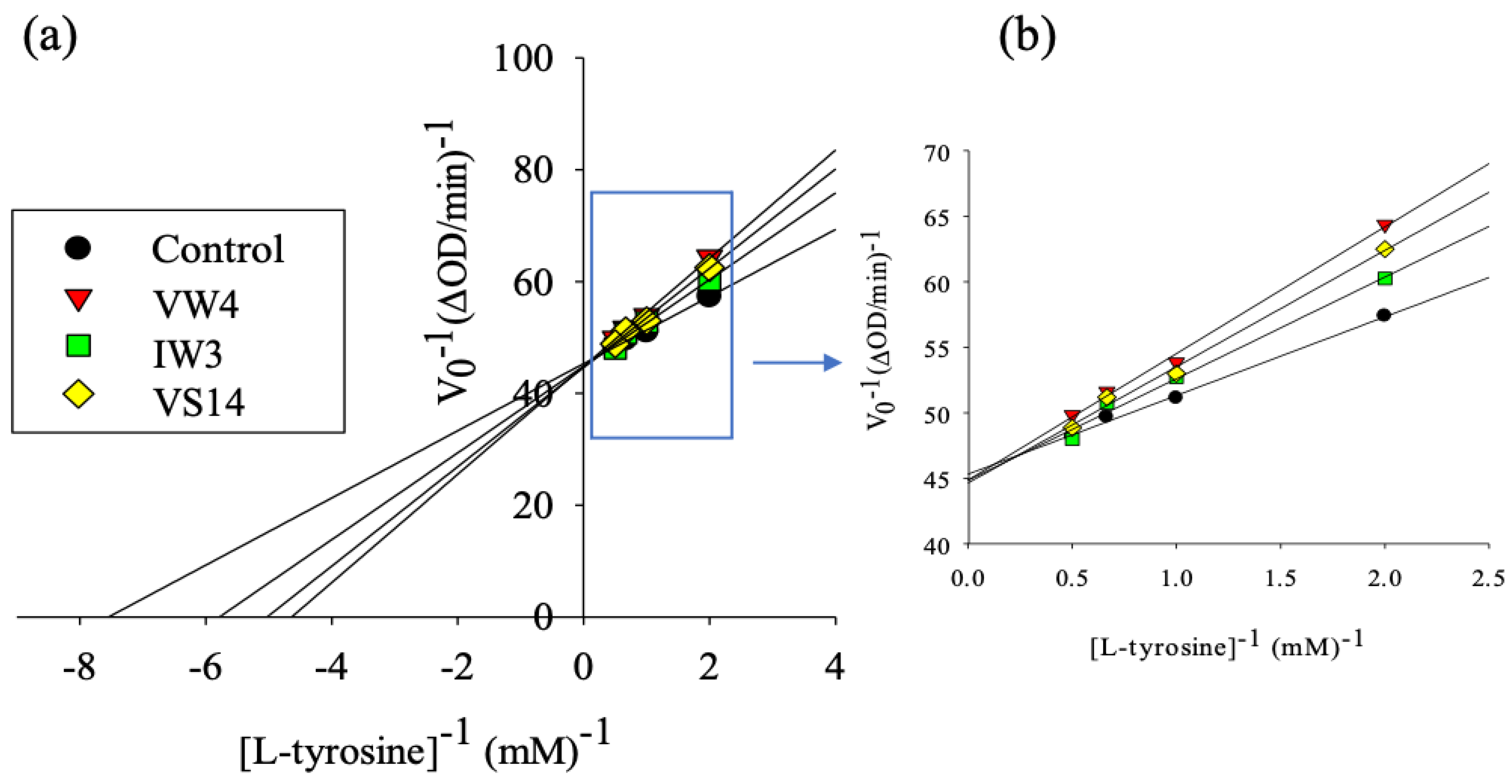
3.4. Tyrosinase Inhibitory Activity of Selected BPs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chakrabarti, S.; Jahandideh, F.; Wu, J. Food-Derived Bioactive Peptides on Inflammation and Oxidative Stress. BioMed Res. Int. 2014, 2014, 608979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halim, N.; Yusof, H.; Sarbon, N. Functional and bioactive properties of fish protein hydolysates and peptides: A comprehensive review. Trends Food Sci. Technol. 2016, 51, 24–33. [Google Scholar] [CrossRef]
- Chalamaiah, M.; Kumar, B.D.; Hemalatha, R.; Jyothirmayi, T. Fish protein hydrolysates: Proximate composition, amino acid composition, antioxidant activities and applications: A review. Food Chem. 2012, 135, 3020–3038. [Google Scholar] [CrossRef]
- Kitts, D.; Weiler, D.D.K.A.K. Bioactive Proteins and Peptides from Food Sources. Applications of Bioprocesses used in Isolation and Recovery. Curr. Pharm. Des. 2003, 9, 1309–1323. [Google Scholar] [CrossRef]
- Daliri, E.B.-M.; Lee, B.H.; Oh, D.H. Current trends and perspectives of bioactive peptides. Crit. Rev. Food Sci. Nutr. 2017, 58, 2273–2284. [Google Scholar] [CrossRef]
- Li-Chan, E.C. Bioactive peptides and protein hydrolysates: Research trends and challenges for application as nutraceuticals and functional food ingredients. Curr. Opin. Food Sci. 2015, 1, 28–37. [Google Scholar] [CrossRef] [Green Version]
- Landreh, M.; Jörnvall, H. Biological activity versus physiological function of proinsulin C-peptide. Cell. Mol. Life Sci. 2021, 78, 1131–1138. [Google Scholar] [CrossRef]
- Jakubczyk, A.; Karaś, M.; Rybczyńska-Tkaczyk, K.; Zielińska, E.; Zieliński, D. Current Trends of Bioactive Peptides—New Sources and Therapeutic Effect. Foods 2020, 9, 846. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Sillero, J.; Gharsallaoui, A.; Prentice, C. Peptides from Fish By-product Protein Hydrolysates and Its Functional Properties: An Overview. Mar. Biotechnol. 2018, 20, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Jafarpour, A.; Gomes, R.M.; Gregersen, S.; Sloth, J.J.; Jacobson, C.; Sørensen, A.-D.M. Characterization of cod (Gadus morhua) frame composition and its valorization by enzymatic hydrolysis. J. Food Compos. Anal. 2020, 89, 103469. [Google Scholar] [CrossRef]
- Sila, A.; Bougatef, A. Antioxidant peptides from marine by-products: Isolation, identification and application in food systems. A review. J. Funct. Foods 2016, 21, 10–26. [Google Scholar] [CrossRef]
- Hajfathalian, M.; Ghelichi, S.; Moreno, P.J.G.; Sørensen, A.-D.M.; Jacobsen, C. Peptides: Production, bioactivity, functionality, and applications. Crit. Rev. Food Sci. Nutr. 2017, 58, 3097–3129. [Google Scholar] [CrossRef] [Green Version]
- Tonolo, F.; Moretto, L.; Grinzato, A.; Fiorese, F.; Folda, A.; Scalcon, V.; Ferro, S.; Arrigoni, G.; Bellamio, M.; Feller, E.; et al. Fermented Soy-Derived Bioactive Peptides Selected by a Molecular Docking Approach Show Antioxidant Properties Involving the Keap1/Nrf2 Pathway. Antioxidants 2020, 9, 1306. [Google Scholar] [CrossRef]
- Tadesse, S.A.; Emire, S.A. Production and processing of antioxidant bioactive peptides: A driving force for the functional food market. Heliyon 2020, 6, e04765. [Google Scholar] [CrossRef] [PubMed]
- Jafarpour, A.; Gregersen, S.; Gomes, R.M.; Marcatili, P.; Olsen, T.H.; Jacobsen, C.; Overgaard, M.T.; Sørensen, A.-D.M. Biofunctionality of Enzymatically Derived Peptides from Codfish (Gadus morhua) Frame: Bulk In Vitro Properties, Quantitative Proteomics, and Bioinformatic Prediction. Mar. Drugs 2020, 18, 599. [Google Scholar] [CrossRef] [PubMed]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM Database of Bioactive Peptides: Current Opportunities. Int. J. Mol. Sci. 2019, 20, 5978. [Google Scholar] [CrossRef] [Green Version]
- Olsen, T.H.; Yesiltas, B.; Marin, F.I.; Pertseva, M.; García-Moreno, P.J.; Gregersen, S.; Overgaard, M.T.; Jacobsen, C.; Lund, O.; Hansen, E.B.; et al. AnOxPePred: Using deep learning for the prediction of antioxidative properties of peptides. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Singh, H. The milk fat globule membrane—A biophysical system for food applications. Curr. Opin. Colloid Interface Sci. 2006, 11, 154–163. [Google Scholar] [CrossRef]
- Kathriarachchi, K.; Leus, M.; Everett, D.W. Oxidation of aldehydes by xanthine oxidase located on the surface of emulsions and washed milk fat globules. Int. Dairy J. 2014, 37, 117–126. [Google Scholar] [CrossRef]
- Battelli, M.G.; Polito, L.; Bolognesi, A. Xanthine oxidoreductase in atherosclerosis pathogenesis: Not only oxidative stress. Atherosclerosis 2014, 237, 562–567. [Google Scholar] [CrossRef] [Green Version]
- Khanna, D.; Fitzgerald, J.D.; Khanna, P.P.; Bae, S.; Singh, M.K.; Neogi, T.; Pillinger, M.H.; Merill, J.; Lee, S.; Prakash, S.; et al. 2012 American College of Rheumatology guidelines for management of gout. Part 1: Systematic nonpharmacologic and pharmacologic therapeutic approaches to hyperuricemia. Arthritis Rheum. 2012, 64, 1431–1446. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Tang, Y.; Liu, C.; Li, J.; Guo, L.; Zhang, Y. Development of a method to screen and isolate potential xanthine oxidase inhibitors from Panax japlcus var via ultrafiltration liquid chromatography combined with counter-current chromatography. Talanta 2015, 134, 665–673. [Google Scholar] [CrossRef]
- Mayer, A.M. Polyphenol oxidases in plants and fungi: Going places? A review. Phytochemistry 2006, 67, 2318–2331. [Google Scholar] [CrossRef]
- Nirmal, N.P.; Benjakul, S. Use of tea extracts for inhibition of polyphenoloxidase and retardation of quality loss of Pacific white shrimp during iced storage. LWT 2011, 44, 924–932. [Google Scholar] [CrossRef]
- Loizzo, M.R.; Tundis, R.; Menichini, F. Natural and Synthetic Tyrosinase Inhibitors as Antibrowning Agents: An Update. Compr. Rev. Food Sci. Food Saf. 2012, 11, 378–398. [Google Scholar] [CrossRef]
- Dai, X.-Y.; Zhang, M.-X.; Wei, X.-Y.; Hider, R.C.; Zhou, T. Novel Multifunctional Hydroxypyridinone Derivatives as Potential Shrimp Preservatives. Food Bioprocess Technol. 2016, 9, 1079–1088. [Google Scholar] [CrossRef]
- Chen, K.; Zhao, D.-Y.; Chen, Y.-L.; Wei, X.-Y.; Li, Y.-T.; Kong, L.-M.; Hider, R.C.; Zhou, T. A Novel Inhibitor Against Mushroom Tyrosinase with a Double Action Mode and Its Application in Controlling the Browning of Potato. Food Bioprocess Technol. 2017, 10, 2146–2155. [Google Scholar] [CrossRef] [Green Version]
- Demirkiran, O.; Sabudak, T.; Ozturk, M.; Topcu, G. Antioxidant and Tyrosinase Inhibitory Activities of Flavonoids from Trifolium nigrescens Subsp. petrisavi. J. Agric. Food Chem. 2013, 61, 12598–12603. [Google Scholar] [CrossRef] [PubMed]
- Perluigi, M.; De Marco, F.; Foppoli, C.; Coccia, R.; Blarzino, C.; Marcante, M.L.; Cini, C. Tyrosinase protects human melanocytes from ROS-generating compounds. Biochem. Biophys. Res. Commun. 2003, 305, 250–256. [Google Scholar] [CrossRef]
- Dalle-Donne, I.; Rossi, R.; Colombo, R.; Giustarini, D.; Milzani, A. Biomarkers of Oxidative Damage in Human Disease. Clin. Chem. 2006, 52, 601–623. [Google Scholar] [CrossRef]
- Wu, H.C. Studies on bioactive peptides as related to antioxidant activities of meat essences and enzyme hydrolysates of mackerel meat. Ph.D. Thesis, National Taiwan Ocean University, Keelung, Taiwan, 2003. [Google Scholar]
- Chou, C.-T.; Kong, Z.-L.; Shiau, C.-Y. Assessment of anti-inflammation function of Val-Trp-Trp-Trp from mackerel hydrolysates using cultured cell model. J. Biosci. Bioeng. 2009, 108, S143. [Google Scholar] [CrossRef]
- You, L.; Zhao, M.; Regenstein, J.M.; Ren, J. Purification and identification of antioxidative peptides from loach (Misgurnus anguillicaudatus) protein hydrolysate by consecutive chromatography and electrospray ionization-mass spectrometry. Food Res. Int. 2010, 43, 1167–1173. [Google Scholar] [CrossRef]
- Kumar, N.S.S.; Nazeer, R.A.; Jaiganesh, R. Purification and identification of antioxidant peptides from the skin protein hydrolysate of two marine fishes, horse mackerel (Magalaspis cordyla) and croaker (Otolithes ruber). Amino Acids 2011, 42, 1641–1649. [Google Scholar] [CrossRef] [PubMed]
- Bougatef, A.; Nedjar-Arroume, N.; Manni, L.; Ravallec, R.; Barkia, A.; Guillochon, D.; Nasri, M. Purification and identification of novel antioxidant peptides from enzymatic hydrolysates of sardinelle (Sardinella aurita) by-products proteins. Food Chem. 2010, 118, 559–565. [Google Scholar] [CrossRef]
- Je, J.-Y.; Park, P.-J.; Kim, S.-K. Antioxidant activity of a peptide isolated from Alaska pollack (Theragra chalcogramma) frame protein hydrolysate. Food Res. Int. 2005, 38, 45–50. [Google Scholar] [CrossRef]
- Je, J.-Y.; Qian, Z.-J.; Byun, H.-G.; Kim, S.-K. Purification and characterization of an antioxidant peptide obtained from tuna backbone protein by enzymatic hydrolysis. Process. Biochem. 2007, 42, 840–846. [Google Scholar] [CrossRef]
- Ren, J.; Zhao, M.; Shi, J.; Wang, J.; Jiang, Y.; Cui, C.; Kakuda, Y.; Xue, S.J. Purification and identification of antioxidant peptides from grass carp muscle hydrolysates by consecutive chromatography and electrospray ionization-mass spectrometry. Food Chem. 2008, 108, 727–736. [Google Scholar] [CrossRef]
- Hsu, K.-C. Purification of antioxidative peptides prepared from enzymatic hydrolysates of tuna dark muscle by-product. Food Chem. 2010, 122, 42–48. [Google Scholar] [CrossRef]
- Majumder, K.; Wu, J. A new approach for identification of novel antihypertensive peptides from egg proteins by QSAR and bioinformatics. Food Res. Int. 2010, 43, 1371–1378. [Google Scholar] [CrossRef]
- Huang, W.; Chakrabarti, S.; Majumder, K.; Jiang, Y.; Davidge, S.T.; Wu, J. Egg-Derived Peptide IRW Inhibits TNF-α-Induced Inflammatory Response and Oxidative Stress in Endothelial Cells. J. Agric. Food Chem. 2010, 58, 10840–10846. [Google Scholar] [CrossRef]
- Liao, W.; Chakrabarti, S.; Davidge, S.T.; Wu, J. Modulatory Effects of Egg White Ovotransferrin-Derived Tripeptide IRW (Ile-Arg-Trp) on Vascular Smooth Muscle Cells against Angiotensin II Stimulation. J. Agric. Food Chem. 2016, 64, 7342–7347. [Google Scholar] [CrossRef] [PubMed]
- Son, M.; Wu, J. Egg white hydrolysate and peptide reverse insulin resistance associated with tumor necrosis factor-α (TNF-α) stimulated mitogen-activated protein kinase (MAPK) pathway in skeletal muscle cells. Eur. J. Nutr. 2019, 58, 1961–1969. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Yan, W.; Ding, S.; Jiang, H.; Ma, Y.; Wang, H.; Fang, J. Effects of IRW and IQW on Oxidative Stress and Gut Microbiota in Dextran Sodium Sulfate-Induced Colitis. Cell. Physiol. Biochem. 2018, 51, 441–451. [Google Scholar] [CrossRef]
- Ma, Y.; Ding, S.; Liu, G.; Fang, J.; Yan, W.; Duraipandiyan, V.; Al-Dhabi, N.A.; Esmail, G.A.; Jiang, H. Egg Protein Transferrin-Derived Peptides IRW and IQW Regulate Citrobacter rodentium-Induced, Inflammation-Related Microbial and Metabolomic Profiles. Front. Microbiol. 2019, 10, 643. [Google Scholar] [CrossRef] [Green Version]
- Liao, W.; Fan, H.; Davidge, S.T.; Wu, J. Egg White–Derived Antihypertensive Peptide IRW (Ile-Arg-Trp) Reduces Blood Pressure in Spontaneously Hypertensive Rats via the ACE2/Ang (1-7)/Mas Receptor Axis. Mol. Nutr. Food Res. 2019, 63, 1900063. [Google Scholar] [CrossRef] [Green Version]
- Alam, N.; Bristi, N.J. Rafiquzzaman Review on in vivo and in vitro methods evaluation of antioxidant activity. Saudi Pharm. J. 2013, 21, 143–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheih, I.-C.; Wu, T.-K.; Fang, T.J. Antioxidant properties of a new antioxidative peptide from algae protein waste hydrolysate in different oxidation systems. Bioresour. Technol. 2009, 100, 3419–3425. [Google Scholar] [CrossRef]
- Cian, R.E.; Martínez-Augustin, O.; Drago, S.R. Bioactive properties of peptides obtained by enzymatic hydrolysis from protein byproducts of Porphyra columbina. Food Res. Int. 2012, 49, 364–372. [Google Scholar] [CrossRef]
- Noro, T.; Oda, Y.; Miyase, T.; Ueno, A.; Fukushima, S. Inhibitors of xanthine oxidase from the flowers and buds of Daphne genkwa. Chem. Pharm. Bull. 1983, 31, 3984–3987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takagi, K.; Mitsunaga, T. Tyrosinase inhibitory activity of proanthocyanidins from woody plants. J. Wood Sci. 2003, 49, 461–465. [Google Scholar] [CrossRef]
- Sae-Leaw, T.; Karnjanapratum, S.; O’Callaghan, Y.C.; O’Keeffe, M.B.; Fitzgerald, R.J.; O’Brien, N.M.; Benjakul, S. Purification and identification of antioxidant peptides from gelatin hydrolysate of seabass skin. J. Food Biochem. 2016, 41, e12350. [Google Scholar] [CrossRef]
- Elias, R.J.; McClements, D.J.; Decker, E.A. Antioxidant Activity of Cysteine, Tryptophan, and Methionine Residues in Continuous Phase β-Lactoglobulin in Oil-in-Water Emulsions. J. Agric. Food Chem. 2005, 53, 10248–10253. [Google Scholar] [CrossRef]
- Ashaolu, T.J. Antioxidative peptides derived from plants for human nutrition: Their production, mechanisms and applications. Eur. Food Res. Technol. 2020, 246, 853–865. [Google Scholar] [CrossRef]
- Chen, H.-M.; Muramoto, K.; Yamauchi, F.; Fujimoto, K.; Nokihara, K. Antioxidative Properties of Histidine-Containing Peptides Designed from Peptide Fragments Found in the Digests of a Soybean Protein. J. Agric. Food Chem. 1998, 46, 49–53. [Google Scholar] [CrossRef]
- Yang, Q.; Cai, X.; Yan, A.; Tian, Y.; Du, M.; Wang, S. A specific antioxidant peptide: Its properties in controlling oxidation and possible action mechanism. Food Chem. 2020, 327, 126984. [Google Scholar] [CrossRef] [PubMed]
- Ajibola, C.F.; Fashakin, J.B.; Fagbemi, T.N.; Aluko, R.E. Effect of Peptide Size on Antioxidant Properties of African Yam Bean Seed (Sphenostylis stenocarpa) Protein Hydrolysate Fractions. Int. J. Mol. Sci. 2011, 12, 6685–6702. [Google Scholar] [CrossRef] [Green Version]
- Qian, Z.-J.; Jung, W.-K.; Kim, S.-K. Free radical scavenging activity of a novel antioxidative peptide purified from hydrolysate of bullfrog skin, Rana catesbeiana Shaw. Bioresour. Technol. 2008, 99, 1690–1698. [Google Scholar] [CrossRef]
- You, L.; Zhao, M.; Cui, C.; Zhao, H.; Yang, B. Effect of degree of hydrolysis on the antioxidant activity of loach (Misgurnus anguillicaudatus) protein hydrolysates. Innov. Food Sci. Emerg. Technol. 2009, 10, 235–240. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; Fitzgerald, R.J. Tryptophan-containing milk protein-derived dipeptides inhibit xanthine oxidase. Peptides 2012, 37, 263–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Kang, X.; Shi, C.; Li, Y.; Majumder, K.; Ning, Z.; Ren, J. Moderation of hyperuricemia in rats via consuming walnut protein hydrolysate diet and identification of new antihyperuricemic peptides. Food Funct. 2017, 9, 107–116. [Google Scholar] [CrossRef]
- Li, Q.; Shi, C.; Wang, M.; Zhou, M.; Liang, M.; Zhang, T.; Yuan, E.; Wang, Z.; Yao, M.; Ren, J. Tryptophan residue enhances in vitro walnut protein-derived peptides exerting xanthine oxidase inhibition and antioxidant activities. J. Funct. Foods 2019, 53, 276–285. [Google Scholar] [CrossRef]
- Rashidinejad, A.; Birch, J. Xanthine Oxidase in Dairy Foods. Encycl. Food Chem. 2019, 374–380. [Google Scholar] [CrossRef]
- Sha, W.; Hou, C.; Yuan, E.; Li, Y.; Ren, J. Different processed milk with residual xanthine oxidase activity and risk of increasing serum uric acid level. Food Biosci. 2021, 40, 100892. [Google Scholar] [CrossRef]
- Masuda, T.; Fukuyama, Y.; Doi, S.; Masuda, A.; Kurosawa, S.; Fujii, S.; Kurosawa, S. Effects of Temperature on the Composition and Xanthine Oxidase Inhibitory Activities of Caffeic Acid Roasting Products. J. Agric. Food Chem. 2019, 67, 8977–8985. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Abdullah; Zhang, Y.; Deng, L.; Zhao, M.; Tang, J.; Zhang, H.; Feng, F.; Wang, J. Exploring the Potential of Novel Xanthine Oxidase Inhibitory Peptide (ACECD) Derived from Skipjack Tuna Hydrolysates Using Affinity-Ultrafiltration Coupled with HPLC–MALDI-TOF/TOF-MS. Food Chem. 2021, 347, 129068. [Google Scholar] [CrossRef] [PubMed]
- Bu, Y.; Wang, F.; Zhu, W.; Li, X. Combining bioinformatic prediction and assay experiment to identify novel xanthine oxidase inhibitory peptides from Pacific bluefin tuna (Thunnus orientalis). E3S Web Conf. 2020, 185, 04062. [Google Scholar] [CrossRef]
- Chang, T.-S. An Updated Review of Tyrosinase Inhibitors. Int. J. Mol. Sci. 2009, 10, 2440–2475. [Google Scholar] [CrossRef] [Green Version]
- Zolghadri, S.; Bahrami, A.; Khan, M.T.H.; Munoz-Munoz, J.; Garcia-Molina, F.; Garcia-Canovas, F.; Saboury, A.A. A comprehensive review on tyrosinase inhibitors. J. Enzym. Inhib. Med. Chem. 2019, 34, 279–309. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide Sequence | Source (By-Product) | Enzyme | Antioxidant Activity | Reference |
|---|---|---|---|---|
| HV5 (HGAYV, 380 Da) | Mackerel meat (Scomber austriasicus) | Papain | LPIA | [31] |
| VW4 (VWWW, 680 Da) | Mackerel meat (Scomber austriasicus) | Papain | LPIA, cell model | [31,32] |
| IW3 (IRW, 340 Da) | Egg white | Thermolysin, pepsin | Cell model | [40] |
| PV4 (PSYV, 464.2 Da) | Loach meat (Misgurnus anguillicaudatus) | Papain | HRSA, DPPH, Cu2+ ion, LPIA | [33] |
| NR6 (NHRYDR, 856 Da) | Horse mackerel (skin) (Magalaspis cordyla) | In vitro-GI tract digestion | RP, Chelating activity, DPPH, HRSA | [34] |
| LL4 (LARL, 471.3 Da) | Sardinelle (head and viscera) (Sardinella aurita) | Crude enzymes from viscera of sardine | DPPH, LPIA, RP | [35] |
| PL4 (PHYL, 528.2 Da) | Sardinelle (head and viscera) (Sardinella aurita) | Crude enzymes from viscera of sardine | DPPH, LPIA, RP | [35] |
| LY6 (LPHSGY, 672 Da) | Alaska pollack (frame protein) (Theragra chalcogramma) | Mackerel intestine crude enzyme | LPIA, HRSA | [36] |
| VS14 (VKAGFAWTANQQLS, 1519 Da) | Tuna (backbone protein) | Pepsin | DPPH, HRSA, superoxide, LPIA | [37] |
| PV8 (PSKYEPFV, 966.3 Da) | Grass carp meat (Ctenopharyngodon idellus) | Alcalase | HRSA, LPIA | [38] |
| LY9 (LPTSEAAKY, 978 Da) | Tuna (dark meat) (Thunnus tonggol) | Orientase | DPPH, LPIA | [39] |
| Sample | ACE Inhibitory | DPP IV Inhibitor | Antioxidant | Others | FRS Score | MCA Score |
|---|---|---|---|---|---|---|
| HV5 | AY, GA, HG, YV | AY, GA, YV | AY | - | 0.52 | 0.24 |
| VW4 | VW | WWW, WW, VW | VW | VW (Alpha-glucosidase inhibitor) | 0.62 | 0.20 |
| IW3 | IRW, RW, IR | RW, IR | IRW, RW, IR | IRW (hypotensive, anti-inflammatory), RW (DPP-III inhibitor), IR (Renin inhibitor, CaMPDE inhibitor) | 0.43 | 0.22 |
| PV4 | PSY, SY, YV | PS, SY, YV | - | - | 0.45 | 0.23 |
| NR6 | RY | DR, HR, NH, YD | - | - | 0.48 | 0.19 |
| LL4 | RL, LA, AR | LA, RL | LARL | LA (DPP-III inhibitor, activating ubiquitin-mediated proteolysis) | 0.36 | 0.24 |
| PL4 | HY, PH | HY, PH, YL | PHY, PHYL | YL (neuropeptide), HY (anti- inflammatory), YL(DPP-III inhibitor) | 0.57 | 0.28 |
| LY6 | GY, SG, PH | LP, GY, HS | LPHSGY, PHS | PH (DPP-III inhibitor) | 0.52 | 0.29 |
| VS14 | AW, VK, KA | KA, FA, TA, WT, AW, NQ, QL, QQ, VK | AW | KA, FA (DPP-III inhibitor) | 0.51 | 0.24 |
| PV8 | KY, YE | EP, KY, PF, PS, SK, YE | PSKYEPFV | PF (DPP-III inhibitor) | 0.51 | 0.19 |
| LY9 | AA, EA, KY, PT | LP, AA, KY, PT, TS | LPTSEAAKY | SE (stimulating), LPTSEAAKY (anticancer), EA (alpha-glucosidase inhibitor) | 0.35 | 0.24 |
| Sample | IC50 (mg/mL) * | IC50 (mM) | Vmax (ΔOD/min) | Km (mM) |
|---|---|---|---|---|
| Control | 0.182 | 0.127 | ||
| Allopurinol | 0.171 ± 0.001 a,+ | 1.253 × 10−3 | 0.223 | 2.544 |
| VW4 | 3.611 ± 0.105 d | 5.310 | 0.183 | 0.147 |
| IW3 | 1.338 ± 0.015 b | 3.935 | 0.188 | 0.478 |
| VS14 | 2.740 ± 0.022 c | 1.804 | 0.196 | 0.816 |
| Sample | IC50 (mg/mL) * | IC50 (mM) | Vmax (ΔOD/min) | Km (mM) |
|---|---|---|---|---|
| Control | 0.022 | 0.132 | ||
| VW4 | 0.853 ± 0.027 a | 1.254 | 0.022 | 0.212 |
| IW3 | 0.984 ± 0.009 c | 2.895 | 0.022 | 0.176 |
| VS14 | 0.903 ± 0.009 b | 0.595 | 0.022 | 0.198 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thaha, A.; Wang, B.-S.; Chang, Y.-W.; Hsia, S.-M.; Huang, T.-C.; Shiau, C.-Y.; Hwang, D.-F.; Chen, T.-Y. Food-Derived Bioactive Peptides with Antioxidative Capacity, Xanthine Oxidase and Tyrosinase Inhibitory Activity. Processes 2021, 9, 747. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9050747
Thaha A, Wang B-S, Chang Y-W, Hsia S-M, Huang T-C, Shiau C-Y, Hwang D-F, Chen T-Y. Food-Derived Bioactive Peptides with Antioxidative Capacity, Xanthine Oxidase and Tyrosinase Inhibitory Activity. Processes. 2021; 9(5):747. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9050747
Chicago/Turabian StyleThaha, Anthony, Bor-Sen Wang, Yu-Wei Chang, Shih-Min Hsia, Tsui-Chin Huang, Chyuan-Yuan Shiau, Deng-Fwu Hwang, and Tai-Yuan Chen. 2021. "Food-Derived Bioactive Peptides with Antioxidative Capacity, Xanthine Oxidase and Tyrosinase Inhibitory Activity" Processes 9, no. 5: 747. https://0-doi-org.brum.beds.ac.uk/10.3390/pr9050747