3.1. Color Parameters
Our data show how cauliflower floret color was affected by the heat and arginine treatments. The effect of heat on the color parameters can be observed in
Table 1. When arginine (Arg) was not applied, lightness (L*) by 17% in high (T
H) or extreme (T
E) temperatures, and chroma (C°) in T
E by 27%
. However, a* and hue (H°) decreased in T
H, although these recovered in T
E.
These results are similar to those from Hodges et al. [
14], who reported that the curd color of the Indian cauliflower cv. Fremont was yellowish to creamy white. Additionally, Singh et al. [
15] indicated that cauliflower showed a wide adaptability to temperature and humidity. Regarding the application of Arg, the data show that the foliar application did not produce significant differences in any parameter, except for L*. These results are similar to the results found by other authors. Taking into account that Singh et al. [
15] reported that mature cauliflowers were yellowish and had a greater luminosity, our results indicate that preharvest spraying of arginine may retard color changes. White-colored cauliflowers are favored by customers, and also, for a cauliflower to be well considered by consumers, it must have a compact, white-colored, and medium-sized curd and be free from any disease or disorder [
16]. Moreover, although the white cauliflower has low levels of colored pigments (e.g., carotenoids, chlorophylls), it is widely consumed due to its content of certain flavonoids and glucosinolates, making it a vegetable that is very rich in antioxidant activity [
17].
3.2. Mineral Content
In this study, 10 cations belonging to two classes—macro- and micronutrients—and 5 anions were studied (
Table 2,
Table 3 and
Table 4). The total macroelement cations varied from 46.01 to 54.65 g kg
−1 DW, and the individual macroelement content ranged between 1.71 and 41.70 g kg
−1 DW. The most abundant cation was potassium (K) (
Table 2). Calcium (Ca) was the only macroelement that was affected by heat stress. Thus, Ca content significantly increased as the temperature increased from T
A to T
H or T
E (
Table 2). The preharvest spraying of arginine produced a decrease in sodium (Na) content, with the lowest value of Na obtained when arginine was applied at 4 mM (
Table 2).
Table 3 shows the most abundant anions present in cauliflower florets. The total anion concentration varied from 9.0 to 30.1 g kg
−1 DW, varying individually from 0.3 to 20.1 g kg
−1 DW. The least abundant and predominant anions were nitrite and phosphates, respectively. In the absence of foliar arginine application, the florets from plants exposed to heat stress showed a significant reduction in the sulfates and phosphates and a sharp increase in chloride and nitrate (
Table 3). With respect to the effect of the foliar spraying,
Table 3 shows that the plants sprayed with arginine had a higher content of chloride, regardless of the temperature, and a high content of nitrate and phosphate at T
E. Nitrite was only found in plants sprayed with 1 mM arginine, and at T
H and T
E (
Table 3).
The total microelements varied in concentration from 83.85 to 115.70 mg kg
−1 DW, with individual microelements oscillating from 0.50 to 35.83 mg kg
−1 DW (
Table 4). The most abundant were iron (Fe) and zinc (Zn), and the least abundant was copper. Heat stress affected all the microelements except for Fe. In this sense, boron (B), Zn, and copper (Cu) were reduced at T
H and T
E, while manganese (Mn) decreased only at T
E (
Table 4). No significant changes were observed in the total microelements as a result of foliar arginine application. Similar results were obtained for Mn and B, but other microelements were affected. In this sense, the concentration of Zn increased regardless of the concentration of arginine at T
H and T
E, whereas the Cu and Fe content was reduced after arginine application (
Table 4).
The contents of macro- and microelements obtained in the cauliflower cv. Moonshine were higher than those of the Australian cauliflowers studied by Cunningham et al. [
18] and those reported by the USDA for raw cauliflower [
19]. Nevertheless, our results are consistent with those previously reported for white cauliflower [
20]. There is a small number of studies aimed at investigating the effect of heat stress on the mineral concentration of plants. Our results obtained due to heat stress application are in agreement with previous reports which indicate that heat stress alters the uptake of nutrients by plants. Among the nutrients whose accumulation was affected by heat stress, with an important decrease in their contents in plants, were potassium, sodium, and phosphorus. The uptake of nutrients could be influenced by several factors, which ultimately results in a decrease in the content of the nutrients in the plant as compared with a control treatment [
21].
Regarding the results after spraying with arginine on the accumulation of nutrients in cauliflower florets, calcium, magnesium, phosphorous, and potassium tended to increase, although a significant reduction in sodium accumulation was also observed. These results matched previous studies carried out on
Rosa hybrid, which indicate that heat-sensitive plants apparently have a reduced capacity of using these nutrients for physiological processes under heat stress [
22]. The heat tolerance of these sensitive plants can be improved by the exogenous application of osmoprotectors such as arginine or polyamines [
7]. A reduction in the Na/K ratio could also be observed in our results. This is supported by a study carried out on pistachio treated exogenously with free polyamines: Kamiab et al. [
23] reported that this not only decreased Na
+ accumulation but also Cl
− accumulation and the Na
+/K
+ ratio. Our data also show an increase in the accumulation of Cl
− and nitrate under moderate heat stress in plants sprayed with 1mM of arginine. A point worth mentioning is that arginine is not only an amino acid but also a precursor of polyamines and the cell signaling molecules glutamine and nitric oxide (NO) [
5]. In accordance with this, in a study performed on mung bean under water stress, it was shown that both nitrogen uptake and nitrogen content improved in plants treated with polyamines [
24].
From a health perspective, the nutrients that should be highlighted are iron, magnesium, and zinc. These cations are crucial for cognitive, behavioral, and motor development. In this sense, iron-deficit anemia and low zinc and magnesium contents are more common in children with Autism Spectrum Disorder [
25,
26]. Our results show an increase in the zinc content of cauliflower florets after the application of foliar arginine and an increase in magnesium after the application of both treatments: heat stress and arginine spraying.
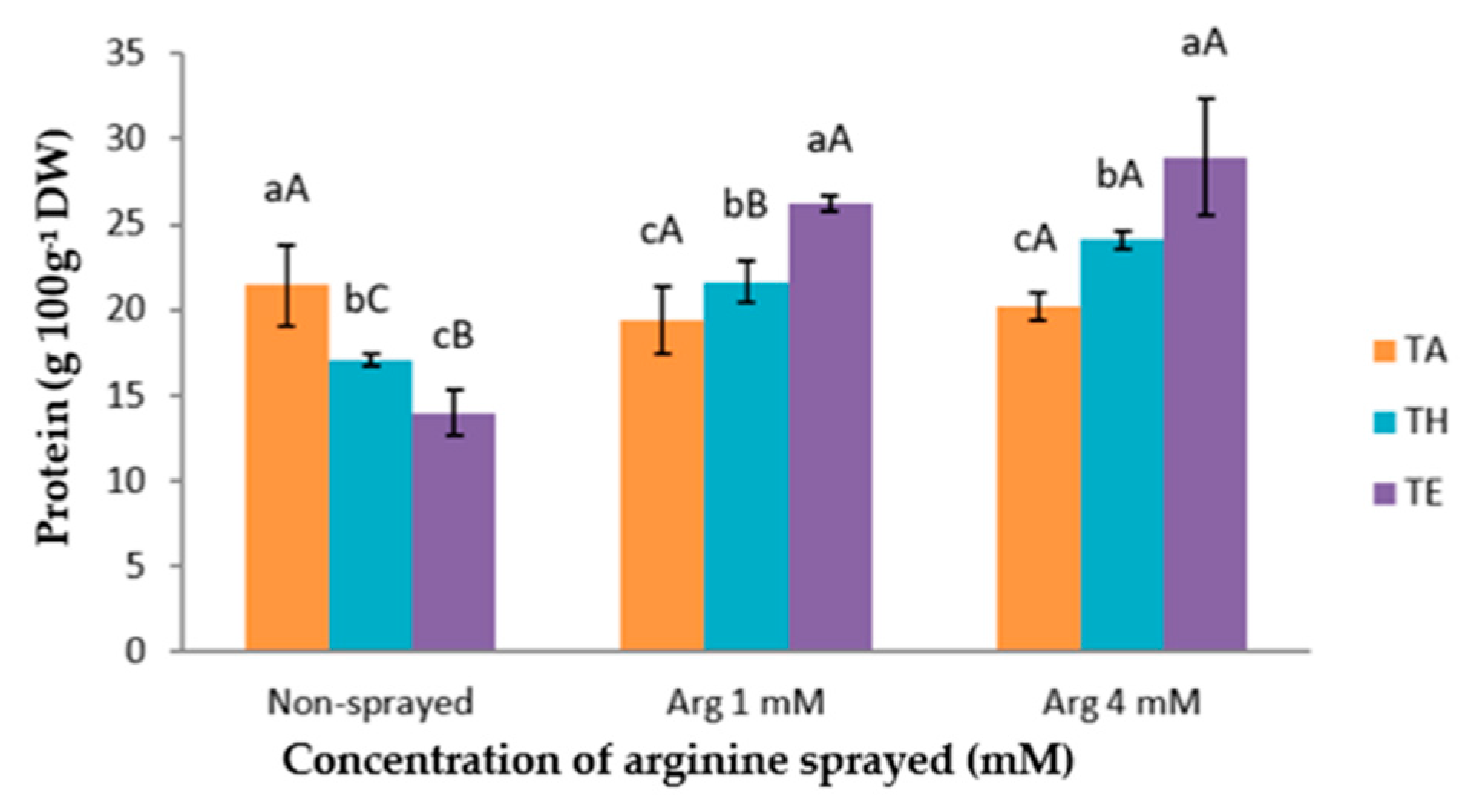
3.3. Total Protein Content
The protein concentration ranged from 14.00 to 28.91 g 100 g
−1 DW (
Figure 1). The heat stress and both foliar arginine treatments affected N uptake and, consequently, the protein content in the cauliflower florets. In this sense, when only the heat stress was applied, in the absence of arginine, a decrease of the protein content was observed. This finding is in agreement with the results reported by several authors, who indicated that heat stress in plants leads to a significant decrease in the nitrogen content [
27]. Additionally, the lower content of proteins in the plants subjected to heat stress could be mainly attributed to the fact that at high temperatures some proteins are not only not synthesized but also denatured [
28]. This decrease in the protein content in plants will depend not only on the characteristics of the stress (intensity and duration) but also on the plant species, and mainly on the phenological period in which the heat stress was applied [
6]. Moreover, our data surprisingly show the highest concentration of proteins after the exogenous application of 4 mM arginine in the cauliflower cv. Moonshine. This is consistent with the results obtained in previous works, where it was found that the protein content increased in wheat plants and mung beans after plants were treated with polyamines, improving their thermotolerance [
24,
29]. In a study carried out on sugarcane (
Saccharum sp.) and alfalfa, it was observed that arginine may have played a positive role in the somatic embryogenesis of sugarcane [
30]. At first, this finding was only expected for the 1 mM arginine treatment and not for the 4 mM one, because all the previous authors had used polyamines or arginine in a range between 1 and 2.5 mM, and 4 mM arginine seemed to be too high for obtaining good results.
3.4. Qualitative and Quantitative Profiles of Free Soluble Sugars
Four free sugars—inositol, glucose, fructose, and sucrose—were evaluated in this study, and glucose was the predominant one in cauliflower cv. Moonshine (
Table 5). The total free sugar content varied within a range from 163.59 to 240.18 g kg
−1 DW in the florets, showing an increase in those subjected to the T
E and foliar arginine at 4 mM (
Table 5).
Hodges et al. [
14] and Bhandari and Kwak [
17] reported that lyophilized cauliflower florets contain over 300 mg g
−1 of total free sugars, suggesting that free sugar levels in cauliflower are genotype dependent. Our data show a reduction in free total soluble sugars as a result of the application of the heat stress treatments. Several authors have reported that the production of osmolytes, including free sugars, in plants under heat stress could be related to the stability of invertase activity, as it stabilizes the structure of the membrane bilayer, exerting protective effects [
31]. However, it is necessary to know that the regulation of starch metabolism induced by abiotic stress can favor starch biosynthesis. [
32]. The fact that the free sugars increased by 25% or 47% depending if 1 or 4 mM of arginine was applied before the short-term heat stress can be attributed to the role of arginine as an osmoregulator [
32,
33]. Our results are similar to those found in a study carried out in wheat plants, where increases in sugars were obtained as a result of spraying different concentrations of arginine [
29]. Although there are some works about exogenous application of polyamines and, specifically, arginine, the mechanism of action of polyamines is still unknown [
32].
3.5. Total Phenolic Compounds and Antioxidant Activity (ABTS+)
It is known that
Brassica crops are a very good source of bioactive substances [
34,
35,
36]. The data on total phenolic compounds (TPC) and antioxidant activity of cauliflower florets ranged between 77.53 and 132.36 mg GAE g
−1 FW, and between 38.17 and 138.73 µM g
−1 DW, respectively (
Table 6). However, as our work was designed and carried out to avoid any effects produced by drought, both the antioxidant activity and the TPC content were enhanced by heat stress (
Table 6). Similar results were previously found for white cauliflower florets [
17]. It has been reported that heat stress generates a significant increase in antioxidant enzyme activities, confirming that plants subjected to stressful temperatures suffer oxidative stress [
37]. As shown in
Table 6, plants treated with foliar arginine showed an increase in both the antioxidant activity and the TPC content in a concentration-dependent manner. Thus, a higher increase in antioxidant activity and the TPC content when 4 mM arginine was sprayed can be observed. These findings match those found by other authors with other plant materials (hot pepper fruits, rice (
Oryza sativa L.), and Indian mustard (
Brassica juncea L.)), who reported that spraying arginine or putrescine increased the TPC of plants under abiotic stress [
6,
38,
39]. It is worth stating that putrescine can be synthesized from arginine via agmatine, through the action of arginine decarboxylase [
40].
The importance of phenols lies in their cell protection role against the oxidative damage suffered by plants due to stress. Furthermore, these compounds are also capable of increasing the stability of the cell membrane [
39]. These results suggest that moderate or extreme heat stress combined with the foliar application of 1 or 4 mM arginine can improve the nutritional value and the content of beneficial substances of cauliflower cv. Moonshine.
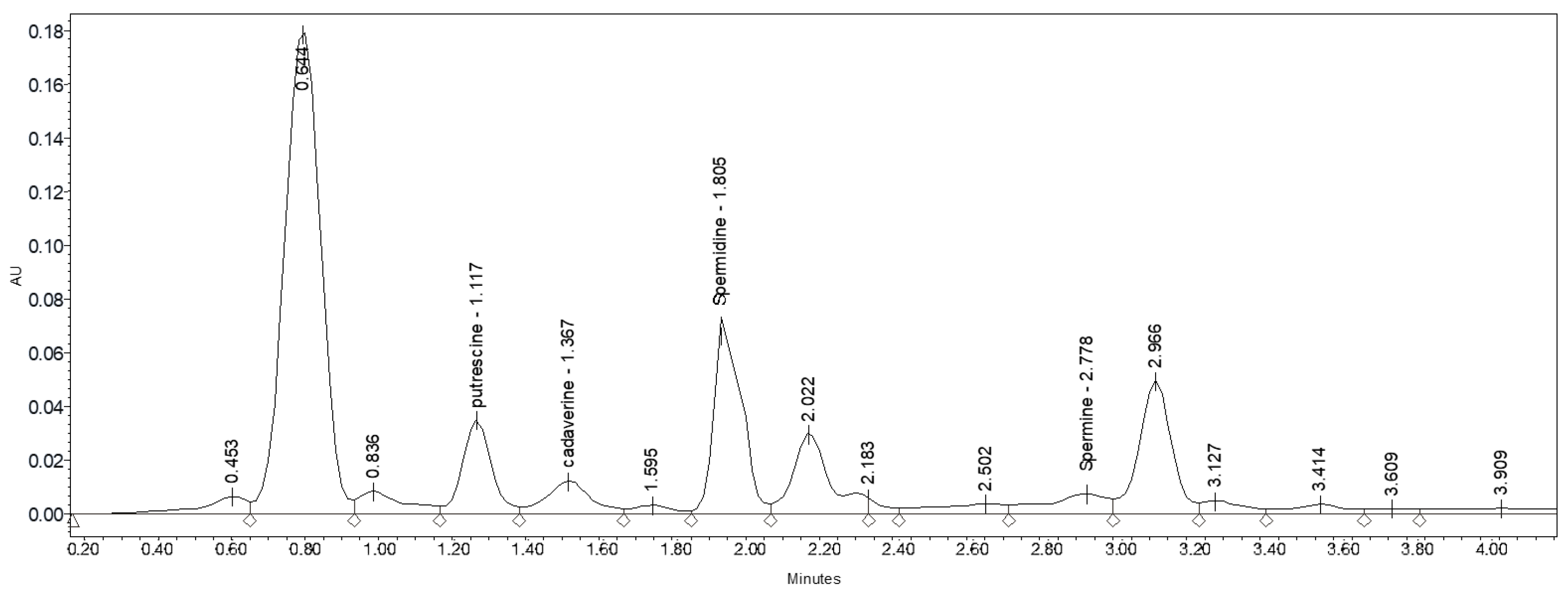
3.6. Determination of Polyamines
Table 7 and
Figure 2 show the chromatographic profile of the polyamines present in cauliflower florets. The total polyamine concentration in the cauliflower florets oscillated between 21.14 and 259.05 nmol g
−1 FW, with the individual contents ranging from 1.88 to 124.69 nmol g
−1 FW. Putrescine was the most abundant polyamine, except at ambient temperature without a foliar treatment or when plants were sprayed with arginine at 1 mM. In these cases, cadaverine was the most abundant polyamine (
Table 7). Specifically, the concentrations of putrescine, cadaverine, spermidine, and spermine varied from 3.19 to 124.69 nmol g
−1 FW, 11.19 to 26.72 nmol g
−1 FW, 1.88 to 80.89 nmol g
−1 FW, and 4.88 to 26.75 nmol g
−1 FW, respectively (
Table 7). The polyamine content in cauliflower plants under heat stress was higher as the temperature increased. These results are in accordance with those obtained in several studies carried out with plant materials such as
Brassica alboglabra Bailey leaves,
O. sativa,
B. juncea,
Vitis vinifera L.,
Valeriana officinalis L.,
Matricaria chamomilla L., and
Origanum majorana L., which reported that the concentrations of polyamines were usually higher under biotic stresses, including heat stress [
41,
42]. Additionally, our results also show that as a consequence of the foliar arginine treatments, the accumulation of endogenous polyamines increased, showing a higher accumulation when 4 mM arginine was applied (
Table 7). The findings of this study are similar to those of other works performed on stressed plants (rice, chickpea, alfalfa,
Nymphoides pelsatum, wheat, tomato, citrus, welsh onion, eggplant, fenugreek sprouts, and bean), where it was shown that the exogenous application of polyamines is an effective approach for endowing plants with tolerance against abiotic stresses such as salinity [
33], cold [
43], drought [
44], heavy metals [
45], osmotic stress [
46], high temperatures [
29], waterlogging [
47], and flooding [
48]. However, until now, there have been no reports focused on the exogenous preharvest application of arginine in cauliflower to reduce heat damage to the florets. Several authors have suggested that the increase in endogenous polyamines as a result of the foliar application of arginine in wheat plants could be due to a decline in ethylene biosynthesis and a synergistic interaction between ethylene and polyamines [
29].
Our data have shown that cauliflower cv. Moonshine under short-term heat stress and sprayed with 4 mM arginine contained 339.60 nmoles g
−1 of polyamines. This value is higher than those observed by Eliassen et al. [
49]. These authors reported that fresh cauliflower contained around 290 nmoles g
−1 of polyamines, indicating that broccoli and cauliflower showed a content of polyamines that was even higher than that obtained for meat or fish. There is emerging evidence that polyamines can be utilized as a therapeutic option for humans. In fact, an earlier work associated the intake of polyamines with a low incidence of cardiovascular events and reduced mortality [
50]. Therefore, it has been estimated that a standard human diet should provide a daily dose of polyamines [
51].


 and
and 


{kind=link}
{kind=link}