
Yield, Grain Quality, and Starch Physicochemical Properties of 2 Elite Thai Rice Cultivars Grown under Varying Production Systems and Soil Characteristics


Abstract
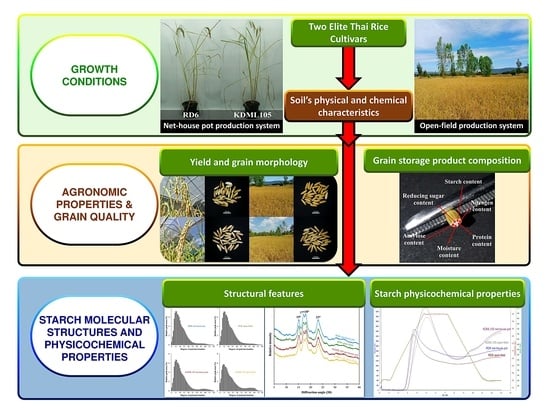
:
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Plant Productivity and Grain Morphology
2.3. Storage Product Composition
2.4. Starch Isolation
2.5. Starch Molecular Structure and Physicochemical Property
2.5.1. Amylopectin Branch Chain Length Distribution
2.5.2. Starch Granule Size Distribution
2.5.3. X-ray Diffraction and Relative Crystallinity
2.5.4. Swelling Power and Water Solubility
2.5.5. Starch Gelatinization and Retrogradation Properties
2.5.6. Starch Pasting Properties
2.6. Statistical Analysis
3. Results
3.1. Rice Productivity and Grain Morphology
3.2. Grain Storage Product Compositions
3.3. Starch Molecular Structure
3.3.1. Chain-Length Distribution of Amylopectin Branches
3.3.2. Starch Particle Size and Granule Size Distribution
3.3.3. Crystalline Structure and Relative Crystallinity
3.4. Starch Physicochemical Properties
3.4.1. Gelatinization and Retrogradation Properties
3.4.2. Starch Swelling Power and Water Solubility
3.4.3. Pasting Properties
4. Discussion
4.1. Effect of Production System and Soil Characteristic on Yield and Grain Appearance Quality of Rice
4.2. Effect of Production System and Soil Characteristic on Grain Storage Product Compositions
4.3. Soil Characteristic Effect on Amylopectin Fine Structure
4.4. Compositional Influences of Starch on Its Median Particle Size, Granule Size Distribution, and Relative Crystallinity under Different Production Systems and Soil Qualities
4.5. Relationship between the Compositional and Structural Features and Physicochemical Properties of Starches from the Two Rice Cultivars Grown under Varying Production Systems and Soil Characteristics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chung, H.-J.; Liu, Q.; Lee, L.; Wei, D. Relationship between the structure, physicochemical properties and in vitro digestibility of rice starches with different amylose contents. Food Hydrocoll. 2011, 25, 968–975. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Jabran, K.; Mahajan, G. Rice Production Worldwide, 1st ed.; Chauhan, B.S., Jabran, K., Mahajan, G., Eds.; Springer: Chem, Switzerland, 2017; Volume 8, p. 563. [Google Scholar]
- Dunwell, J.M. Transgenic cereals: Current status and future prospects. J. Cereal Sci. 2014, 59, 419–434. [Google Scholar] [CrossRef]
- US Department of Agriculture (USDA). Grain: World Markets and Trade. Available online: https://apps.fas.usda.gov/psdonline/circulars/grain.pdf (accessed on 22 April 2021).
- Seck, P.A.; Diagne, A.; Mohanty, S.; Wopereis, M.C. Crops that feed the world 7: Rice. Food Secur. 2012, 4, 7–24. [Google Scholar] [CrossRef]
- Juliano, B. Rice: Overview. In Encyclopedia of Food Grains; Wrigley, C., Corke, H., Seetharaman, K., Faubion, J., Eds.; Academic Press: Oxford, UK, 2016; Volume 1, pp. 125–129. [Google Scholar]
- Tanakasempipat, P.; Thepgumpanat, P. Thailand’s 2020 Rice Exports Forecast to Fall to Lowest in Seven Years. Available online: https://www.reuters.com/article/thailand-rice/thailands-2020-rice-exports-forecast-to-fall-to-lowest-in-seven-years-idUSL4N29L18V (accessed on 29 April 2021).
- Rerkasem, B. The rice value chain: A case study of Thai rice. ASR Chiang Mai Univ. J. Soc. Sci. Hum. 2017, 4, 1–26. [Google Scholar] [CrossRef]
- Thitisaksakul, M.; Sangwongchai, W.; Mungmonsin, U.; Promrit, P.; Krusong, K.; Wanichthanarak, K.; Tananuwong, K. Granule morphological and structural variability of Thai certified glutinous rice starches in relation to thermal, pasting, and digestible properties. Cereal Chem. 2020, 98, 492–506. [Google Scholar] [CrossRef]
- Wanichthanarak, K.; Thitisaksakul, M. ThRSDB: A database of Thai rice starch composition, molecular structure and functionality. Database 2020, 2020, 1–9. [Google Scholar] [CrossRef]
- Asante, M.D. Breeding rice for improved grain quality. In Advances in International Rice Research; Li, J.Q., Ed.; IntechOpen: London, UK, 2017; pp. 69–89. [Google Scholar]
- Suwannaporn, P.; Linnemann, A. Rice-eating quality among consumers in different rice grain preference countries. J. Sens. Stud. 2008, 23, 1–13. [Google Scholar] [CrossRef]
- Nishimura, T.; Cha-Um, S.; Takagaki, M.; Ohyama, K.; Kirdmanee, C. Survival percentage, photosynthetic abilities and growth characters of two indica rice (Oryza sativa L. spp. indica) cultivars in response to iso-osmotic stress. Span. J. Agric. Res. 2011, 9, 262–270. [Google Scholar] [CrossRef]
- Srisompun, O. Rice Economy in Northeast Thailand: Current Status and Challenges. Available online: https://ap.fftc.org.tw/article/1863 (accessed on 5 May 2021).
- Department of International Trade Promotion (DITP). Thai Rice: Quality That Comes with Variety. Available online: https://www.thaitradeusa.com/home/?p=25485&fbclid=IwAR0dIMrB_vTSm8bnolDggaDXsNYwklYDxj-e96yzW93xEFYF1fh9-t9xcro (accessed on 30 April 2021).
- Jiamyangyuen, S.; Wichaphon, J.; Boonmeejoy, J. Classification of rice cultivars by using chemical, physicochemical, thermal, hydration properties, and cooking quality. Food Appl. Biosci. J. 2019, 7, 42–62. [Google Scholar]
- Oko, A.; Ubi, B.; Dambaba, N. Rice cooking quality and physico-chemical characteristics: A comparative analysis of selected local and newly introduced rice varieties in Ebonyi State, Nigeria. Public Health Nutr. 2012, 2, 43–49. [Google Scholar]
- Nakamura, S.; Satoh, H.; Ohtsubo, K.I. Development of formulae for estimating amylose content, amylopectin chain length distribution, and resistant starch content based on the iodine absorption curve of rice starch. Biosci. Biotechnol. Biochem. 2015, 79, 443–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tangsrianugul, N.; Wongsagonsup, R.; Suphantharika, M. Physicochemical and rheological properties of flour and starch from Thai pigmented rice cultivars. Int. J. Biol. Macromol. 2019, 137, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Sirivongpaisal, P.; Hill, S.E.; Pradipasena, P.; Mitchell, J.R. Amylose Content and Amylopectin Fine Structure of Thai Rice Starches. In Proceedings of the the 3rd Conference on Starch Technology, National Center for Genetic Engineering and Biotechnology (BIOTEC), Bangkok, Thailand, 4 November 2005; pp. 221–226. [Google Scholar]
- Office of Agricultural Economics (OAE). At Air Humidity of 15%, in-Season Rice is Divided into Four Cultivars by Province. Available online: https://www.oae.go.th/assets/portals/1/fileups/prcaidata/files/rice%204%20type.pdf (accessed on 16 October 2021).
- Pitiphunpong, S.; Suwannaporn, P. Physicochemical properties of KDML 105 rice cultivar from different cultivated locations in Thailand. J. Sci. Food Agric. 2009, 89, 2186–2190. [Google Scholar] [CrossRef]
- Gajda, A.M.; Czyż, E.A.; Ukalska-Jaruga, A. Comparison of the Effects of Different Crop Production Systems on Soil Physico-Chemical Properties and Microbial Activity under Winter Wheat. Agronomy 2020, 10, 1130. [Google Scholar] [CrossRef]
- Ishfaq, M.; Akbar, N.; Zulfiqar, U.; Hussain, S.; Murtza, K.; Batool, Z.; Ashraf, U.; Alyemeni, M.N.; Ahmad, P. Influence of Nitrogen Management Regimes on Milling Recovery and Grain Quality of Aromatic Rice in Different Rice Production Systems. Agronomy 2020, 10, 1841. [Google Scholar] [CrossRef]
- Suebpongsang, P.; Ekasingh, B.; Cramb, R. Commercialisation of Rice Farming in Northeast Thailand, 1st ed.; Cramb, R., Ed.; Palgrave Macmillan: Singapore, 2020; p. 456. [Google Scholar]
- Zhao, C.; Chen, M.; Li, X.; Dai, Q.; Xu, K.; Guo, B.; Hu, Y.; Wang, W.; Huo, Z. Effects of Soil Types and Irrigation Modes on Rice Root Morphophysiological Traits and Grain Quality. Agronomy 2021, 11, 120. [Google Scholar] [CrossRef]
- Anderson, S.H. Cropping Systems, Effect on Soil Physical Properties; Gliński, J.H.J., Lipiec, J., Eds.; Springer: Dordrecht, The Netherlands, 2011; Volume 180. [Google Scholar]
- Alhaj Hamoud, Y.; Wang, Z.; Guo, X.; Shaghaleh, H.; Sheteiwy, M.; Chen, S.; Qiu, R.; Elbashier, M. Effect of irrigation regimes and soil texture on the potassium utilization efficiency of rice. Agronomy 2019, 9, 100. [Google Scholar] [CrossRef] [Green Version]
- Dou, F.; Soriano, J.; Tabien, R.E.; Chen, K. Soil texture and cultivar effects on rice (Oryza sativa L.) grain yield, yield components and water productivity in three water regimes. PLoS ONE 2016, 11, e0150549. [Google Scholar] [CrossRef] [PubMed]
- Suriya-Arunroj, D.; Chaiyawat, P.; Fukai, S.; Blamey, P. Identification of nutrients limiting rice growth in soils of Northeast Thailand under water-limiting and non-limiting conditions. Plant Prod. Sci. 2000, 3, 417–421. [Google Scholar] [CrossRef]
- Atapattu, A.J.; Prasantha, B.R.; Amaratunga, K.; Marambe, B. Increased rate of potassium fertilizer at the time of heading enhances the quality of direct seeded rice. Chem. Biol. Technol. 2018, 5, 1–9. [Google Scholar] [CrossRef]
- Das, S.K. Role of micronutrient in rice cultivation and management strategy in organic agriculture—A reappraisal. Agric. Sci. 2014, 5, 765–769. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, J.; Kandel, M.; Subedi, S.; Shah, K.K. Role of nutrients in rice (Oryza sativa L.): A review. Agrica 2020, 9, 53–62. [Google Scholar] [CrossRef]
- Panda, B.B.; Sharma, S.; Mohapatra, P.K.; Das, A. Application of excess nitrogen, phosphorus, and potassium fertilizers leads to lowering of grain iron content in high-yielding tropical rice. Commun. Soil Sci. Plant Anal. 2012, 43, 2590–2602. [Google Scholar] [CrossRef]
- Val-Torregrosa, B.; Bundó, M.; San Segundo, B. Crosstalk between Nutrient Signalling Pathways and Immune Responses in Rice. Agriculture 2021, 11, 747. [Google Scholar] [CrossRef]
- Kamal, M.A.; Rasul, F.; Zohaib, A.; Ahmad, K.; Abbas, T.; Rasool, T.; Nawaz, M. Effect of NPK application at various levels on yield and quality of two rice hybrids. Sci. J. Seoul Sci. 2016, 4, 14–19. [Google Scholar]
- Zhang, J.; Tong, T.; Potcho, M.; Huang, S.; Ma, L.; Tang, X. Nitrogen Effects on Yield, Quality and Physiological Characteristics of Giant Rice. Agronomy 2020, 10, 1816. [Google Scholar] [CrossRef]
- Shankar, T.; Malik, G.C.; Banerjee, M.; Dutta, S.; Maitra, S.; Praharaj, S.; Sairam, M.; Kumar, D.S.; Dessoky, E.S.; Hassan, M.M. Productivity and Nutrient Balance of an Intensive Rice–Rice Cropping System Are Influenced by Different Nutrient Management in the Red and Lateritic Belt of West Bengal, India. Plants 2021, 10, 1622. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Hua, J.; Li, Y.; Chen, Y.; Yang, J. Effects of phosphorous on grain quality of upland and paddy rice under different cultivations. Chin. J. Rice Sci. 2011, 25, 399–406. [Google Scholar]
- Fageria, N.; Slaton, N.; Baligar, V. Nutrient management for improving lowland rice productivity and sustainability. Adv. Agron. 2003, 80, 63–152. [Google Scholar]
- Paramasivam, K.; Saraswathi, R.; Subramanian, M.; Marimuthu, R.; Parthasarathy, P.; Ramanathan, S.; Manuel, S.; Ranganathan, T. A high yielding medium duration rice variety for Tamil Nadu. Mad. Agric. J. 2005, 92, 1–3. [Google Scholar]
- Sun, T.; Tong, L.; Zhao, S.; Wang, H.; Han, Y.; Zhang, Z.; Jin, Z. Effects of nitrogen fertilizer application on starch quality, activities and gene expression levels of related enzymes in rice endosperm. Chin. J. Rice Sci. 2018, 32, 475–484. [Google Scholar]
- Dou, Z.; Tang, S.; Li, G.; Liu, Z.; Ding, C.; Chen, L.; Wang, S.; Ding, Y. Application of nitrogen fertilizer at heading stage improves rice quality under elevated temperature during grain-filling stage. Crop Sci. 2017, 57, 2183–2192. [Google Scholar] [CrossRef] [Green Version]
- Bahmaniar, M.; Ranjbar, G. Response of rice (Oryza sativa L.) cooking quality properties to nitrogen and potassium application. Pak. J. Biol. Sci. 2007, 10, 1880–1884. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Chen, J.; Chen, L.; Wang, Z.; Zhang, H.; Yang, J. Grain quality changes and responses to nitrogen fertilizer of japonica rice cultivars released in the Yangtze River Basin from the 1950s to 2000s. Crop J. 2015, 3, 285–297. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Liu, S.; Gao, H.; Zhang, H.; Wei, H. High-Level Nitrogen Application Decreases Eating Quality of Rice by Affecting Pasting, Rheological, Water Migration, and Oral Processing Properties. Food Sci. Technol. 2021, 1, 754–761. [Google Scholar]
- Zhu, D.-W.; Zhang, H.-C.; Guo, B.-W.; Ke, X.; Dai, Q.-G.; Wei, H.-Y.; Hui, G.; Hu, Y.-J.; Cui, P.-Y.; Huo, Z.-Y. Effects of nitrogen level on yield and quality of japonica soft super rice. J. Integr. Agric. 2017, 16, 1018–1027. [Google Scholar] [CrossRef]
- Sangwongchai, W.; Krusong, K.; Thitisaksakul, M. Salt Tolerance at Vegetative Stage is Partially Associated with Changes in Grain Quality and Starch Physicochemical Properties of Rice Exposed to Salinity Stress at Reproductive Stage. J. Sci. Food Agric. 2021. [Google Scholar] [CrossRef] [PubMed]
- Thitisaksakul, M.; Tananuwong, K.; Shoemaker, C.F.; Chun, A.; Tanadul, O.-U.-M.; Labavitch, J.M.; Beckles, D.M. Effects of timing and severity of salinity stress on rice (Oryza sativa L.) yield, grain composition, and starch functionality. J. Agric. Food Chem. 2015, 63, 2296–2304. [Google Scholar] [CrossRef] [Green Version]
- Counce, P.A.; Keisling, T.C.; Mitchell, A.J. A uniform, objective, and adaptive system for expressing rice development. Crop Sci. 2000, 40, 436–443. [Google Scholar] [CrossRef] [Green Version]
- Luengwilai, K.; Beckles, D.M. Climacteric ethylene is not essential for initiating chilling injury in tomato (Solanum lycopersicum) cv. Ailsa Craig. J. Stored Prod. Res. 2010, 1, 1–8. [Google Scholar]
- Odenigbo, A.M.; Ngadi, M.; Ejebe, C.; Nwankpa, C.; Danbaba, N.; Ndindeng, S.; Manful, J. Study on the gelatinization properties and amylose content of rice varieties from Nigeria and Cameroun. Int. J. Food Sci. Nutr. 2013, 2, 181–186. [Google Scholar] [CrossRef]
- Lumdubwong, N.; Seib, P. Rice starch isolation by alkaline protease digestion of wet-milled rice flour. J. Cereal Sci. 2000, 31, 63–74. [Google Scholar] [CrossRef]
- Kittisuban, P.; Lee, B.-H.; Suphantharika, M.; Hamaker, B.R. Slow glucose release property of enzyme-synthesized highly branched maltodextrins differs among starch sources. Carbohydr. Polym. 2014, 107, 182–191. [Google Scholar] [CrossRef]
- Lee, C.-K.; Le, Q.-T.; Kim, Y.-H.; Shim, J.-H.; Lee, S.-J.; Park, J.-H.; Lee, K.-P.; Song, S.-H.; Auh, J.H.; Lee, S.-J. Enzymatic synthesis and properties of highly branched rice starch amylose and amylopectin cluster. J. Agric. Food Chem. 2008, 56, 126–131. [Google Scholar] [CrossRef]
- Ramadoss, B.R.; Gangola, M.P.; Agasimani, S.; Jaiswal, S.; Venkatesan, T.; Sundaram, G.R.; Chibbar, R.N. Starch granule size and amylopectin chain length influence starch in vitro enzymatic digestibility in selected rice mutants with similar amylose concentration. J. Food Sci. Technol. 2019, 56, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, N.; Isono, N.; Noda, T. Relationship of granule size distribution and amylopectin structure with pasting, thermal, and retrogradation properties in wheat starch. J. Agric. Food Chem. 2010, 58, 1180–1188. [Google Scholar] [CrossRef]
- Shujun, W.; Jinglin, Y.; Wenyuan, G. Use of X-ray diffractometry (XRD) for identification of Fritillaria according to geographical origin. Am. J. Biochem. Biotechnol. 2005, 1, 207–211. [Google Scholar] [CrossRef]
- Vandeputte, G.E.; Derycke, V.; Geeroms, J.; Delcour, J.A. Rice starches. II. Structural aspects provide insight into swelling and pasting properties. J. Cereal Sci. 2003, 38, 53–59. [Google Scholar] [CrossRef]
- Wang, L.; Xie, B.; Shi, J.; Xue, S.; Deng, Q.; Wei, Y.; Tian, B. Physicochemical properties and structure of starches from Chinese rice cultivars. Food Hydrocoll. 2010, 24, 208–216. [Google Scholar] [CrossRef]
- Tananuwong, K.; Malila, Y. Changes in physicochemical properties of organic hulled rice during storage under different conditions. Food Chem. 2011, 125, 179–185. [Google Scholar] [CrossRef]
- Kong, X.; Zhu, P.; Sui, Z.; Bao, J. Physicochemical properties of starches from diverse rice cultivars varying in apparent amylose content and gelatinisation temperature combinations. Food Chem. 2015, 172, 433–440. [Google Scholar] [CrossRef] [PubMed]
- AACC. AACC International Approved Method 76-21.01 General Pasting Method for Wheat or Rye Flour or Starch Using the Rapid Visco Analyser, 11th ed.; Cereals & Grains Association: St. Paul, MN, USA, 2000. [Google Scholar]
- Singh, N.; Pal, N.; Mahajan, G.; Singh, S.; Shevkani, K. Rice grain and starch properties: Effects of nitrogen fertilizer application. Carbohydr. Polym. 2011, 86, 219–225. [Google Scholar] [CrossRef]
- Patindol, J.A.; Siebenmorgen, T.J.; Wang, Y.J.; Lanning, S.B.; Counce, P.A. Impact of elevated nighttime air temperatures during kernel development on starch properties of field-grown rice. Cereal Chem. 2014, 91, 350–357. [Google Scholar] [CrossRef] [Green Version]
- Baker, N.T.; Capel, P.D. Environmental Factors That Influence the Location of Crop Agriculture in the Conterminous United States; U.S. Geological Survey: Michigan, MI, USA, 2011; p. 72.
- Guo, L.; Liu, M.; Tao, Y.; Zhang, Y.; Li, G.; Lin, S.; Dittert, K. Innovative water-saving ground cover rice production system increases yield with slight reduction in grain quality. Agric. Syst. 2020, 180, 102795. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, C.; Zhao, Y.; Sun, S.; Liu, Z. Effects of growing seasons and genotypes on the accumulation of cadmium and mineral nutrients in rice grown in cadmium contaminated soil. Sci. Total Environ. 2017, 579, 1282–1288. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, R.P.; Pede, V.O.; McKinley, J.; Velarde, O.; Demont, M. Rice grain quality and consumer preferences: A case study of two rural towns in the Philippines. PLoS ONE 2016, 11, e0150345. [Google Scholar] [CrossRef] [Green Version]
- Cameron, D.K.; Wang, Y.J.; Moldenhauer, K.A. Comparison of starch physicochemical properties from medium-grain rice cultivars grown in California and Arkansas. Starch/Staerke 2007, 59, 600–608. [Google Scholar] [CrossRef]
- Li, X.; Wu, L.; Geng, X.; Xia, X.; Wang, X.; Xu, Z.; Xu, Q. Deciphering the Environmental Impacts on Rice Quality for Different Rice Cultivated Areas. Rice 2018, 11, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarwar, G.; Schmeisky, H.; Hussain, N.; Muhammad, S.; Ibrahim, M.; Safdar, E. Improvement of soil physical and chemical properties with compost application in rice-wheat cropping system. Pak. J. Bot. 2008, 40, 275–282. [Google Scholar]
- Mannan, M.; Bhuiya, M.; Hossain, H.; Akhand, M. Optimization of nitrogen rate for aromatic Basmati rice (Oriza sativa L.). Bangladesh J. Agric. Res. 2010, 35, 157–165. [Google Scholar] [CrossRef] [Green Version]
- Deng, F.; Wang, L.; Pu, S.-L.; Mei, X.-F.; Li, S.-X.; Li, Q.-P.; Ren, W.-J. Shading stress increases chalkiness by postponing caryopsis development and disturbing starch characteristics of rice grains. Agric. For. Meteorol. 2018, 263, 49–58. [Google Scholar] [CrossRef]
- Wang, L.; Deng, F.; Ren, W.-J.; Yang, W.-Y. Effects of shading on starch pasting characteristics of indica hybrid rice (Oryza sativa L.). PLoS ONE 2013, 8, e68220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, G.; Liu, X.; Wang, Q.; Xiong, R.; Hang, Y. Effects of screenhouse cultivation and organic materials incorporation on global warming potential in rice fields. Environ. Sci. Pollut. Res. 2017, 24, 6581–6591. [Google Scholar] [CrossRef]
- Deng, F.; Li, Q.; Chen, H.; Zeng, Y.; Li, B.; Zhong, X.; Wang, L.; Ren, W. Relationship between chalkiness and the structural and thermal properties of rice starch after shading during grain-filling stage. Carbohydr. Polym. 2021, 252, 117–212. [Google Scholar] [CrossRef] [PubMed]
- Abbasi, M.; Sepaskhah, A. Response of different rice cultivars (Oryza sativa L.) to water-saving irrigation in greenhouse conditions. Int. J. Plant Prod. 2011, 5, 37–48. [Google Scholar] [CrossRef]
- Aref, F.; Ebrahimi, H. Physiological characterization of rice under salinity stress during vegetative and reproductive stages. Indian J. Sci. Technol. 2012, 5, 2578–2586. [Google Scholar] [CrossRef]
- Lizana, X.C.; Riegel, R.; Gomez, L.D.; Herrera, J.; Isla, A.; McQueen-Mason, S.J.; Calderini, D.F. Expansins expression is associated with grain size dynamics in wheat (Triticum aestivum L.). J. Exp. Bot. 2010, 61, 1147–1157. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Li, M.; Ashraf, U.; Liu, S.; Zhang, J. Exploring the relationships between yield and yield-related traits for rice varieties released in China from 1978 to 2017. Front. Plant Sci. 2019, 10, 543. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Chen, W.; Xu, Z. Relationship between grain yield and quality in rice germplasms grown across different growing areas. Breed. Sci. 2015, 65, 226–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhekale, J.; Thorat, B.; Kunkerkar, R.; Raut, S.; Desai, S.; Keluskar, M.; Gavai, M. Correlation Studies in Hybrid Rice (Oryza sativa L.). Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 1158–1164. [Google Scholar] [CrossRef]
- Joshi, R.; Mo, C.; Lee, W.-H.; Lee, S.H.; Cho, B.-K. Review of rice quality under various growth and storage conditions and its evaluation using spectroscopic technology. Biosyst. Eng. 2015, 40, 124–136. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Cozzolino, D.; Cynkar, W.; Dambergs, R.; Janik, L.; O’neill, B.; Colby, C.; Gishen, M. Preliminary study on the application of visible–near infrared spectroscopy and chemometrics to classify Riesling wines from different countries. Food Chem. 2008, 106, 781–786. [Google Scholar] [CrossRef]
- Kumari, M.; Asthir, B. Transformation of sucrose to starch and protein in rice leaves and grains under two establishment methods. Rice Sci. 2016, 23, 255–265. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.-J.; Zhan, H.-H. Effect of soil texture on starch accumulation and activities of key enzymes of starch synthesis in the Kernel of ZM 9023. Agric. Sci. China 2008, 7, 686–691. [Google Scholar] [CrossRef]
- Leesawatwong, M.; Jamjod, S.; Kuo, J.; Dell, B.; Rerkasem, B. Nitrogen fertilizer increases seed protein and milling quality of rice. Cereal Chem. 2005, 82, 588–593. [Google Scholar] [CrossRef] [Green Version]
- Seila, S.; Tizzotti, M.J.; Hasjim, J.; GILBERT, R.G. Effects of rice variety and growth location in Cambodia on grain composition and starch structure. Rice Sci. 2014, 21, 47–58. [Google Scholar]
- Aboubacar, A.; Moldenhauer, K.A.; McClung, A.M.; Beighley, D.H.; Hamaker, B.R. Effect of growth location in the United States on amylose content, amylopectin fine structure, and thermal properties of starches of long grain rice cultivars. Cereal Chem. 2006, 83, 93–98. [Google Scholar] [CrossRef]
- Inukai, T.; Hirayama, Y. Comparison of starch levels reduced by high temperature during ripening in japonica rice lines near-isogenic for the Wx locus. J. Agron. Crop Sci. 2010, 196, 296–301. [Google Scholar] [CrossRef]
- Jeon, J.-S.; Ryoo, N.; Hahn, T.-R.; Walia, H.; Nakamura, Y. Starch biosynthesis in cereal endosperm. Plant Physiol. Biochem. 2010, 48, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Hirano, H.-Y.; Sano, Y. Enhancement of Wx gene expression and the accumulation of amylose in response to cool temperatures during seed development in rice. Plant Cell Physiol. 1998, 39, 807–812. [Google Scholar] [CrossRef]
- Patindol, J.; Siebenmorgen, T.; Wang, Y.-J. Impact of environmental factors on rice starch structure: A review. Starch/Staerke 2014, 67, 42–54. [Google Scholar] [CrossRef]
- Teng, B.; Xi, M.; Du, S.; Zhou, Y.; Zhang, Y.; Zhang, Y.; Luo, Z.; Wu, W. Structural and functional properties of indica rice starch as influenced by late-stage nitrogen fertilization. Int. J. Food Prop. 2021, 24, 249–263. [Google Scholar] [CrossRef]
- Dhital, S.; Shrestha, A.K.; Hasjim, J.; Gidley, M.J. Physicochemical and structural properties of maize and potato starches as a function of granule size. J. Agric. Food Chem. 2011, 59, 10151–10161. [Google Scholar] [CrossRef]
- Pan, D.D.; Jane, J.-L. Internal structure of normal maize starch granules revealed by chemical surface gelatinization. Biomacromolecules 2000, 1, 126–132. [Google Scholar] [CrossRef]
- Ali, I.; He, L.; Ullah, S.; Quan, Z.; Wei, S.; Iqbal, A.; Munsif, F.; Shah, T.; Xuan, Y.; Luo, Y. Biochar addition coupled with nitrogen fertilization impacts on soil quality, crop productivity, and nitrogen uptake under double-cropping system. Food Energy Secur. 2020, 9, e208. [Google Scholar] [CrossRef]
- He, W.; Lin, L.; Wang, J.; Zhang, L.; Liu, Q.; Wei, C. Inhibition of starch branching enzymes in waxy rice increases the proportion of long branch-chains of amylopectin resulting in the comb-like profiles of starch granules. Plant Sci. 2018, 277, 177–187. [Google Scholar] [CrossRef]
- Wenting, W.; Wenpei, C.; Ke, X.; Hui, G.; Haiyan, W.; Hongcheng, Z. Effects of Early-and Late-Sowing on Starch Accumulation and Associated Enzyme Activities During Grain Filling Stage in Rice. Rice Sci. 2021, 28, 191–199. [Google Scholar] [CrossRef]
- Yamakawa, H.; Hirose, T.; Kuroda, M.; Yamaguchi, T. Comprehensive expression profiling of rice grain filling-related genes under high temperature using DNA microarray. Plant Physiol. 2007, 144, 258–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, N.; Nakaura, Y.; Inouchi, N.; Nishinari, K. Fine structure, thermal and viscoelastic properties of starches separated from Indica rice cultivars. Starch/Staerke 2007, 59, 10–20. [Google Scholar] [CrossRef]
- Cai, J.; Man, J.; Huang, J.; Liu, Q.; Wei, W.; Wei, C. Relationship between structure and functional properties of normal rice starches with different amylose contents. Carbohydr. Polym. 2015, 125, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Gu, J.; Wang, Z.; Wei, C.; Yang, J.; Zhang, J. Comparison of structural and functional properties of wheat starch under different soil drought conditions. Sci. Rep. 2017, 7, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.; Kumar, P.; Gautam, V.; Rengasamy, B.; Adhikari, B.; Udayakumar, M.; Sarkar, A.K. Root transcriptome of two contrasting indica rice cultivars uncovers regulators of root development and physiological responses. Sci. Rep. 2016, 6, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Himmelsbach, D.; Manful, J.; Coker, R. Changes in rice with variable temperature parboiling: Thermal and spectroscopic assessment. Cereal Chem. 2008, 85, 384–390. [Google Scholar] [CrossRef] [Green Version]
- Zhu, F. Relationships between amylopectin internal molecular structure and physicochemical properties of starch. Trends Food Sci. Technol. 2018, 78, 234–242. [Google Scholar] [CrossRef]
- Koroteeva, D.A.; Kiseleva, V.I.; Sriroth, K.; Piyachomkwan, K.; Bertoft, E.; Yuryev, P.V.; Yuryev, V.P. Structural and thermodynamic properties of rice starches with different genetic background: Part 1. Differentiation of amylopectin and amylose defects. Int. J. Biol. Macromol. 2007, 41, 391–403. [Google Scholar] [CrossRef] [PubMed]
- Man, J.; Lin, L.; Wang, Z.; Wang, Y.; Liu, Q.; Wei, C. Different structures of heterogeneous starch granules from high-amylose rice. J. Agric. Food Chem. 2014, 62, 11254–11263. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Qin, F.; Zhou, W.; Xu, B.; Chen, C.; Chen, Y.; Wang, Y.; Gu, M.; Liu, Q. Comparison of the crystalline properties and structural changes of starches from high-amylose transgenic rice and its wild type during heating. Food Chem. 2011, 128, 645–652. [Google Scholar] [CrossRef]
- Sodhi, N.S.; Singh, N. Morphological, thermal and rheological properties of starches separated from rice cultivars grown in India. Food Chem. 2003, 80, 99–108. [Google Scholar] [CrossRef]
- Srichuwong, S.; Jane, J.-I. Physicochemical properties of starch affected by molecular composition and structures: A review. Food Sci. Biotechnol. 2007, 16, 663–674. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Sources | Physical Characteristics | Chemical Characteristics | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Texture (%) | Soil Textural Classes | OM (%) | LR (t CaCO3 ha−1) | Total N (%) | Available P (mg kg−1) | Available K (mg kg−1) | CEC (cmol(+) kg−1) | Water pH (1:1) | EC (1:5) (dS/m) | |||
| Sand | Silt | Clay | ||||||||||
| Potting mix | - | - | - | - | 9.56 | - | 0.249 | 1500.00 | 2941.30 | 25.60 | 6.28 | 1.834 |
| RD6 field | 86.59 | 11.24 | 2.17 | Sand | 0.33 | 1.512 | 0.025 | 8.17 | 9.50 | 3.40 | 5.46 | 0.018 |
| KDML105 field | 58.00 | 23.89 | 18.11 | Sandy loam | 0.25 | 7.600 | 0.032 | 2.67 | 29.23 | 8.20 | 5.41 | 0.006 |
| Cultivars | Production Systems | Panicle Number | Total Grain Weight (g) | 100-Grain Weight (g) | %Fertility |
|---|---|---|---|---|---|
| RD6 | Net-house pot | 5.11 ± 0.35 | 15.32 ± 1.68 | 2.23 ± 0.04 | 91.78 ± 1.57 |
| Open-field | 14.67 ± 0.67** | 43.76 ± 1.25 ** | 2.53 ± 0.05 ** | 91.56 ± 1.58 | |
| KDML105 | Net-house pot | 4.89 ± 0.26 | 15.90 ± 0.77 | 2.34 ± 0.03 | 93.44 ± 1.17 |
| Open-field | 9.33 ± 0.69 ** | 32.17 ± 1.16 ** | 2.58 ± 0.03 ** | 91.78 ± 1.39 |
| Cultivars | Production Systems | Length (mm) | Width (mm) | Length-to- Width Ratio | Perimeter (mm) | Volume (mm3) |
|---|---|---|---|---|---|---|
| RD6 | Net-house pot | 6.65 ± 0.03 | 2.39 ± 0.01 | 2.78 ± 0.01 | 14.19 ± 0.05 | 19.86 ± 0.20 |
| Open-field | 7.02 ± 0.03 ** | 2.39 ± 0.01 | 2.95 ± 0.02 ** | 14.77 ± 0.05 ** | 20.84 ± 0.23 ** | |
| KDML105 | Net-house pot | 7.19 ± 0.09 | 2.22 ± 0.01 | 3.24 ± 0.04 | 14.77 ± 0.16 | 18.56 ± 0.34 |
| Open-field | 7.63 ± 0.01 ** | 2.24 ± 0.01 | 3.41 ± 0.02 ** | 15.51 ± 0.03 ** | 20.13 ± 0.23 ** |
| Cultivars | Production Systems | Reducing Sugar Content (mg gFW−1) | Starch Content (mg gFW−1) | N Content (%) | Protein Content (N × 5.95) (%) | Amylose Content (%) | Moisture Content (%) |
|---|---|---|---|---|---|---|---|
| RD6 | Net-house pot | 0.54 ± 0.02 | 719.73 ± 19.40 | 1.41 ± 0.06 | 8.37 ± 0.33 | 2.15 ± 0.22 | 9.35 ± 0.12 |
| Open-field | 0.63 ± 0.02 * | 743.89 ± 20.60 | 1.30 ± 0.00 | 7.76 ± 0.02 | 1.69 ± 0.09 | 10.73 ± 0.95 | |
| KDML105 | Net-house pot | 0.81 ± 0.08 | 723.47 ± 7.36 | 1.34 ± 0.01 | 7.97 ± 0.07 | 13.15 ± 0.63 | 8.75 ± 0.66 |
| Open-field | 0.95 ± 0.05 | 748.92 ± 10.90 | 1.40 ± 0.05 | 8.31 ± 0.28 | 12.05 ± 0.82 | 8.42 ± 0.34 |
| Cultivars | Production Systems | Amylopectin Branch Chain Length Distribution (%) | ||||
|---|---|---|---|---|---|---|
| DP6–12 (A Chains) | DP13–24 (B1 Chains) | DP25–36 (B2 Chains) | DP ≥ 37 (B3+ Chains) | CL (AGU) | ||
| RD6 | Net-house pot | 33.66 ± 0.24 | 52.68 ± 0.11 | 9.72 ± 0.05 | 3.95 ± 0.18 | 17.00 ± 0.06 |
| Open-field | 34.01 ± 0.14 | 52.23 ± 0.15 | 10.01 ± 0.07 * | 3.75 ± 0.26 | 16.97 ± 0.08 | |
| KDML105 | Net-house pot | 32.46 ± 0.33 * | 51.20 ± 0.18 ** | 9.53 ± 0.12 | 6.80 ± 0.48 | 17.94 ± 0.14 |
| Open-field | 30.61 ± 0.25 | 47.84 ± 0.18 | 12.17 ± 0.26 ** | 9.38 ± 0.04 ** | 19.00 ± 0.04 ** | |
| Cultivars | Production Systems | Starch Granule Size Distribution (%) | d(0.5) (μm) | Relative Crystallinity (%) | ||
|---|---|---|---|---|---|---|
| C-Type <5 (μm) | B-Type 5–15 (μm) | A-Type >15 (μm) | ||||
| RD6 | Net-house pot | 34.44 ± 1.87 | 61.49 ± 2.33 | 4.07 ± 0.46 | 5.75 ± 0.42 | 38.01 ± 0.57 * |
| Open-field | 38.02 ± 0.90 | 58.38 ± 0.73 | 3.60 ± 0.25 | 5.61 ± 0.13 | 35.65 ± 0.58 | |
| KDML105 | Net-house pot | 40.43 ± 0.20 | 55.49 ± 0.34 * | 4.07 ± 0.54 | 5.65 ± 0.04 ** | 33.52 ± 1.06 |
| Open-field | 57.39 ± 3.03 * | 41.79 ± 2.95 | 0.82 ± 0.15 ** | 4.73 ± 0.13 | 34.68 ± 0.44 | |
| Cultivars | Production Systems | Gelatinization Properties | ||||
|---|---|---|---|---|---|---|
| To (°C) | Tp (°C) | Tc (°C) | ∆T (°C) | ∆H * (J/g) | ||
| RD6 | Net-house pot | 58.63 ± 0.11 ** | 66.82 ± 0.29 | 76.46 ± 0.19 | 17.83 ± 0.22 | 11.31 ± 0.13 |
| Open-field | 55.31 ± 0.09 | 66.60 ± 0.20 | 78.29 ± 0.20 ** | 22.98 ± 0.18 ** | 15.02 ± 0.06 ** | |
| KDML105 | Net-house pot | 57.82 ± 0.15 | 67.90 ± 0.16 | 75.88 ± 0.23 | 18.06 ± 0.25 ** | 11.18 ± 0.09 |
| Open-field | 59.67 ± 0.16 ** | 67.90 ± 0.32 | 75.63 ± 0.21 | 15.96 ± 0.33 | 10.74 ± 0.25 | |
| Cultivars | Production Systems | Retrogradation Properties | |||||
|---|---|---|---|---|---|---|---|
| To(r) (°C) | Tp(r) (°C) | Tc(r) (°C) | ∆T(r) (°C) | ∆H * (r) (J/g) | R (%) | ||
| RD6 | Net-house pot | 46.04 ± 0.05 ** | 62.26 ± 0.05 ** | 70.28 ± 0.30 ** | 24.24 ± 0.25 ** | 5.12 ± 0.13 ** | 45.34 ± 1.02 ** |
| Open-field | 43.46 ± 0.21 | 54.52 ± 0.30 | 64.12 ± 0.12 | 20.67 ± 0.20 | 2.89 ± 0.07 | 19.22 ± 0.41 | |
| KDML105 | Net-house pot | 45.06 ± 0.24 | 54.56 ± 0.11 | 60.37 ± 0.25 | 15.31 ± 0.46 | 2.65 ± 0.03 | 23.87 ± 0.31 |
| Open-field | 46.74 ± 0.21 ** | 55.23 ± 0.20 * | 63.51 ± 0.36 ** | 16.78 ± 0.49 | 2.44 ± 0.11 | 22.71 ± 1.73 | |
| Cultivars | Production Systems | SP | S | Pasting Properties | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| (g/g) | (%) | PTemp (°C) | PTime (min) | PV (cP) | TV (cP) | BD (cP) | FV (cP) | SB (cP) | ||
| RD6 | Net-house pot | 39.33 ± 1.19 | 4.23 ± 0.86 | 68.35 ± 0.20 | 3.72 ± 0.03 | 2833.00 ± 141.42 | 1772.00 ± 182.78 | 1136.00 ± 45.34 | 2004.25 ± 162.52 | 282.25 ± 7.16 |
| Open-field | 38.91 ± 1.15 | 3.97 ± 0.39 | 69.16 ± 0.40 | 3.67 ± 0.03 | 2583.50 ± 233.31 | 1348.00 ± 204.52 | 1310.75 ± 55.55 | 1564.25 ± 188.58 | 266.25 ± 14.65 | |
| KDML105 | Net-house pot | 25.21 ± 0.49 | 11.52 ± 0.71 | 72.55 ± 0.18 | 5.05 ± 0.09 | 3279.00 ± 67.22 * | 1387.75 ± 75.04 | 1891.25 ± 65.56 | 2557.50 ± 129.32 | 1219.75 ± 49.05 |
| Open-field | 23.68 ± 0.52 | 13.17 ± 0.76 | 72.18 ± 0.41 | 5.00 ± 0.08 | 2960.50 ± 97.41 | 1221.00 ± 32.71 | 1714.50 ± 77.45 | 2410.75 ± 53.43 | 1189.75 ± 40.81 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sangwongchai, W.; Tananuwong, K.; Krusong, K.; Thitisaksakul, M. Yield, Grain Quality, and Starch Physicochemical Properties of 2 Elite Thai Rice Cultivars Grown under Varying Production Systems and Soil Characteristics. Foods 2021, 10, 2601. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10112601
Sangwongchai W, Tananuwong K, Krusong K, Thitisaksakul M. Yield, Grain Quality, and Starch Physicochemical Properties of 2 Elite Thai Rice Cultivars Grown under Varying Production Systems and Soil Characteristics. Foods. 2021; 10(11):2601. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10112601
Chicago/Turabian StyleSangwongchai, Wichian, Kanitha Tananuwong, Kuakarun Krusong, and Maysaya Thitisaksakul. 2021. "Yield, Grain Quality, and Starch Physicochemical Properties of 2 Elite Thai Rice Cultivars Grown under Varying Production Systems and Soil Characteristics" Foods 10, no. 11: 2601. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10112601